Progesterone receptor
The progesterone receptor (PR), also known as NR3C3 or nuclear receptor subfamily 3, group C, member 3, is a protein found inside cells. It is activated by the steroid hormone progesterone.
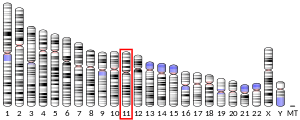

In humans, PR is encoded by a single PGR gene residing on chromosome 11q22,[5][6][7] it has two isoforms, PR-A and PR-B, that differ in their molecular weight.[8][9][10] The PR-B is the positive regulator of the effects of progesterone, while PR-A serve to antagonize the effects of PR-B.[11]
Mechanism
Progesterone is necessary to induce the progesterone receptors. When no binding hormone is present the carboxyl terminal inhibits transcription. Binding to a hormone induces a structural change that removes the inhibitory action. Progesterone antagonists prevent the structural reconfiguration.
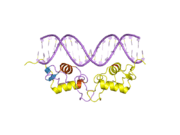
After progesterone binds to the receptor, restructuring with dimerization follows and the complex enters the nucleus and binds to DNA. There transcription takes place, resulting in formation of messenger RNA that is translated by ribosomes to produce specific proteins.
Structure
| Progesterone receptor, N-terminal | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | Progest_rcpt_N | ||||||||
| Pfam | PF02161 | ||||||||
| InterPro | IPR000128 | ||||||||
| |||||||||
In common with other steroid receptors, the progesterone receptor has a N-terminal regulatory domain, a DNA binding domain, a hinge section, and a C-terminal ligand binding domain. A special transcription activation function (TAF), called TAF-3, is present in the progesterone receptor-B, in a B-upstream segment (BUS) at the amino acid terminal. This segment is not present in the receptor-A.
Isoforms
As demonstrated in progesterone receptor-deficient mice, the physiological effects of progesterone depend completely on the presence of the human progesterone receptor (hPR), a member of the steroid-receptor superfamily of nuclear receptors. The single-copy human (hPR) gene uses separate promoters and translational start sites to produce two isoforms, hPR-A and -B, which are identical except for an additional 165 amino acids present only in the N terminus of hPR-B.[12] Although hPR-B shares many important structural domains with hPR-A, they are in fact two functionally distinct transcription factors, mediating their own response genes and physiological effects with little overlap. Selective ablation of PR-A in a mouse model, resulting in exclusive production of PR-B, unexpectedly revealed that PR-B contributes to, rather than inhibits, epithelial cell proliferation both in response to estrogen alone and in the presence of progesterone and estrogen. These results suggest that in the uterus, the PR-A isoform is necessary to oppose estrogen-induced proliferation as well as PR-B-dependent proliferation.
Functional polymorphisms
Six variable sites, including four polymorphisms and five common haplotypes have been identified in the human PR gene .[13] One promoter region polymorphism, +331G/A, creates a unique transcription start site. Biochemical assays showed that the +331G/A polymorphism increases transcription of the PR gene, favoring production of hPR-B in an Ishikawa endometrial cancer cell line.[14]
Several studies have now shown no association between progesterone receptor gene +331G/A polymorphisms and breast or endometrial cancers.[15][16] However, these follow-up studies lacked the sample size and statistical power to make any definitive conclusions, due to the rarity of the +331A SNP. It is currently unknown which if any polymorphisms in this receptor are of significance to cancer. A study of in 21 non-European populations identified two markers within the PROGINS haplotype of the PR gene as positively correlated with ovarian and breast cancer. [17]
Animal studies
Development
Knockout mice of the PR have been found to have severely impaired lobuloalveolar development of the mammary glands[18] as well as delayed but otherwise normal mammary ductal development at puberty.[19][20]
Behavior
During rodent perinatal life, progesterone receptor (PR) is known to be transiently expressed in both the ventral tegmental area (VTA) and the medial prefrontal cortex (mPFC) of the mesocortical dopaminergic pathway. PR activity during this time period impacts the development of dopaminergic innervation of the mPFC from the VTA. If PR activity is altered, a change in dopaminergic innervation of the mPFC is seen and tyrosine hydroxylase (TH), the rate-limiting enzyme for dopamine synthesis, in the VTA will also be impacted. TH expression in this area is an indicator of dopaminergic activity, which is believed to be involved in normal and critical development of complex cognitive behaviors that are mediated by the mesocortical dopaminergic pathway, such as working memory, attention, behavioral inhibition, and cognitive flexibility.[21]
Research has shown that when a PR antagonist, such as RU 486, is administered to rats during the neonatal period, decreased tyrosine hydroxylase immunoreactive (TH-ir) cells density, a strong co-expresser with PR-immunoreactivity (PR-ir), is seen in the mPFC of juvenile rodents. Later on, in adulthood, decreased levels of TH-ir in the VTA are also shown. This alteration in TH-ir fiber expression, an indicator of altered dopaminergic activity resulting from neonatal PR antagonist administration, has been shown to impair later performance on tasks that measure behavioral inhibition and impulsivity, as well as cognitive flexibility in adulthood. Similar cognitive flexibility impairments were also seen in PR knockout mice as a result of reduced dopaminergic activity in the VTA.[21]
Conversely, when a PR agonist, such as 17α-hydroxyprogesterone caproate, is administered to rodents during perinatal life, as the mesocortical dopaminergic pathway is developing, dopaminergic innervation of the mPFC increases. As a result, TH-ir fiber density also increases. Interestingly, this increase in TH-ir fibers and dopaminergic activity is also linked to impaired cognitive flexibility with increased perseveration later on in life.[22]
In combination, these findings suggest that PR expression during early development impact later cognitive functioning in rodents. Furthermore, it appears as though abnormal levels of PR activity during this critical period of mesocortical dopaminergic pathway development may have profound effects on specific behavioral neural circuits involved in the formation of later complex cognitive behavior.[21][22]
Ligands
Agonists
- Endogenous progestogens (e.g., progesterone)
- Synthetic progestogens (e.g., norethisterone, levonorgestrel, medroxyprogesterone acetate, megestrol acetate, dydrogesterone, drospirenone)
Mixed
- Selective progesterone receptor modulators (e.g., ulipristal acetate, telapristone acetate, vilaprisan, asoprisnil, asoprisnil ecamate)[23]
Antagonists
Interactions
Progesterone receptor has been shown to interact with:
References
- GRCh38: Ensembl release 89: ENSG00000082175 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000031870 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Misrahi M, Atger M, d'Auriol L, Loosfelt H, Meriel C, Fridlansky F, Guiochon-Mantel A, Galibert F, Milgrom E (March 1987). "Complete amino acid sequence of the human progesterone receptor deduced from cloned cDNA". Biochemical and Biophysical Research Communications. 143 (2): 740–8. doi:10.1016/0006-291X(87)91416-1. PMID 3551956.
- Law ML, Kao FT, Wei Q, Hartz JA, Greene GL, Zarucki-Schulz T, Conneely OM, Jones C, Puck TT, O'Malley BW (May 1987). "The progesterone receptor gene maps to human chromosome band 11q13, the site of the mammary oncogene int-2". Proceedings of the National Academy of Sciences of the United States of America. 84 (9): 2877–81. doi:10.1073/pnas.84.9.2877. PMC 304763. PMID 3472240.
- ensembl.org, Gene: ESR1 (ENSG00000091831)
- Gadkar-Sable S, Shah C, Rosario G, Sachdeva G, Puri C (2005). "Progesterone receptors: various forms and functions in reproductive tissues". Frontiers in Bioscience. 10 (1–3): 2118–30. doi:10.2741/1685. PMID 15970482.
- Kase NG, Speroff L, Glass RL (1999). Clinical gynecologic endocrinology and infertility. Hagerstown, MD: Lippincott Williams & Wilkins. ISBN 978-0-683-30379-7.
- Fritz MA, Speroff L (2005). Clinical gynecologic endocrinology and infertility. Hagerstown, MD: Lippincott Williams & Wilkins. ISBN 978-0-7817-4795-0.
- Falcone T, Hurd WW (22 May 2013). Clinical Reproductive Medicine and Surgery: A Practical Guide. Springer Science & Business Media. pp. 39–. ISBN 978-1-4614-6837-0.
- Kastner P, Krust A, Turcotte B, Stropp U, Tora L, Gronemeyer H, Chambon P (May 1990). "Two distinct estrogen-regulated promoters generate transcripts encoding the two functionally different human progesterone receptor forms A and B". The EMBO Journal. 9 (5): 1603–14. doi:10.1002/j.1460-2075.1990.tb08280.x. PMC 551856. PMID 2328727.
- Terry KL, De Vivo I, Titus-Ernstoff L, Sluss PM, Cramer DW (March 2005). "Genetic variation in the progesterone receptor gene and ovarian cancer risk". American Journal of Epidemiology. 161 (5): 442–51. doi:10.1093/aje/kwi064. PMC 1380205. PMID 15718480.
- De Vivo I, Huggins GS, Hankinson SE, Lescault PJ, Boezen M, Colditz GA, Hunter DJ (September 2002). "A functional polymorphism in the promoter of the progesterone receptor gene associated with endometrial cancer risk". Proceedings of the National Academy of Sciences of the United States of America. 99 (19): 12263–8. doi:10.1073/pnas.192172299. PMC 129433. PMID 12218173.
- Feigelson HS, Rodriguez C, Jacobs EJ, Diver WR, Thun MJ, Calle EE (June 2004). "No association between the progesterone receptor gene +331G/A polymorphism and breast cancer". Cancer Epidemiology, Biomarkers & Prevention. 13 (6): 1084–5. PMID 15184270.
- Dossus L, Canzian F, Kaaks R, Boumertit A, Weiderpass E (July 2006). "No association between progesterone receptor gene +331G/A polymorphism and endometrial cancer". Cancer Epidemiology, Biomarkers & Prevention. 15 (7): 1415–6. doi:10.1158/1055-9965.EPI-06-0215. PMID 16835347.
- https://pubmed.ncbi.nlm.nih.gov/22121098/
- Macias H, Hinck L (2012). "Mammary gland development". Wiley Interdisciplinary Reviews. Developmental Biology. 1 (4): 533–57. doi:10.1002/wdev.35. PMC 3404495. PMID 22844349.
- Hilton HN, Graham JD, Clarke CL (September 2015). "Minireview: Progesterone Regulation of Proliferation in the Normal Human Breast and in Breast Cancer: A Tale of Two Scenarios?". Molecular Endocrinology. 29 (9): 1230–42. doi:10.1210/me.2015-1152. PMC 5414684. PMID 26266959.
- Aupperlee MD, Leipprandt JR, Bennett JM, Schwartz RC, Haslam SZ (May 2013). "Amphiregulin mediates progesterone-induced mammary ductal development during puberty". Breast Cancer Research. 15 (3): R44. doi:10.1186/bcr3431. PMC 3738150. PMID 23705924.
- Willing J, Wagner CK (2016). "Progesterone Receptor Expression in the Developing Mesocortical Dopamine Pathway: Importance for Complex Cognitive Behavior in Adulthood". Neuroendocrinology. 103 (3–4): 207–22. doi:10.1159/000434725. PMC 4675705. PMID 26065828.
- Willing J, Wagner CK (January 2016). "Exposure to the Synthetic Progestin, 17α-Hydroxyprogesterone Caproate During Development Impairs Cognitive Flexibility in Adulthood". Endocrinology. 157 (1): 77–82. doi:10.1210/en.2015-1775. PMC 4701880. PMID 26556535.
- Knutson TP, Lange CA (April 2014). "Tracking progesterone receptor-mediated actions in breast cancer". Pharmacology & Therapeutics. 142 (1): 114–25. doi:10.1016/j.pharmthera.2013.11.010. PMC 3943696. PMID 24291072.
- Zhang XL, Zhang D, Michel FJ, Blum JL, Simmen FA, Simmen RC (June 2003). "Selective interactions of Kruppel-like factor 9/basic transcription element-binding protein with progesterone receptor isoforms A and B determine transcriptional activity of progesterone-responsive genes in endometrial epithelial cells". The Journal of Biological Chemistry. 278 (24): 21474–82. doi:10.1074/jbc.M212098200. PMID 12672823.
- Giangrande PH, Kimbrel EA, Edwards DP, McDonnell DP (May 2000). "The opposing transcriptional activities of the two isoforms of the human progesterone receptor are due to differential cofactor binding". Molecular and Cellular Biology. 20 (9): 3102–15. doi:10.1128/MCB.20.9.3102-3115.2000. PMC 85605. PMID 10757795.
- Nawaz Z, Lonard DM, Smith CL, Lev-Lehman E, Tsai SY, Tsai MJ, O'Malley BW (February 1999). "The Angelman syndrome-associated protein, E6-AP, is a coactivator for the nuclear hormone receptor superfamily". Molecular and Cellular Biology. 19 (2): 1182–9. doi:10.1128/mcb.19.2.1182. PMC 116047. PMID 9891052.
Further reading
- Butnor KJ, Burchette JL, Robboy SJ (July 1999). "Progesterone receptor activity in leiomyomatosis peritonealis disseminata". International Journal of Gynecological Pathology. 18 (3): 259–64. doi:10.1097/00004347-199907000-00012. PMID 12090595.
- Leonhardt SA, Boonyaratanakornkit V, Edwards DP (November 2003). "Progesterone receptor transcription and non-transcription signaling mechanisms". Steroids. 68 (10–13): 761–70. doi:10.1016/S0039-128X(03)00129-6. PMID 14667966.
- Conneely OM, Mulac-Jericevic B, Lydon JP (November 2003). "Progesterone-dependent regulation of female reproductive activity by two distinct progesterone receptor isoforms". Steroids. 68 (10–13): 771–8. doi:10.1016/S0039-128X(03)00126-0. PMID 14667967.
- Bagchi MK, Tsai SY, Tsai MJ, O'Malley BW (April 1992). "Ligand and DNA-dependent phosphorylation of human progesterone receptor in vitro". Proceedings of the National Academy of Sciences of the United States of America. 89 (7): 2664–8. doi:10.1073/pnas.89.7.2664. PMC 48722. PMID 1557371.
- Kastner P, Krust A, Turcotte B, Stropp U, Tora L, Gronemeyer H, Chambon P (May 1990). "Two distinct estrogen-regulated promoters generate transcripts encoding the two functionally different human progesterone receptor forms A and B". The EMBO Journal. 9 (5): 1603–14. doi:10.1002/j.1460-2075.1990.tb08280.x. PMC 551856. PMID 2328727.
- Guiochon-Mantel A, Loosfelt H, Lescop P, Sar S, Atger M, Perrot-Applanat M, Milgrom E (June 1989). "Mechanisms of nuclear localization of the progesterone receptor: evidence for interaction between monomers". Cell. 57 (7): 1147–54. doi:10.1016/0092-8674(89)90052-4. PMID 2736623.
- Fernandez MD, Carter GD, Palmer TN (January 1983). "The interaction of canrenone with oestrogen and progesterone receptors in human uterine cytosol". British Journal of Clinical Pharmacology. 15 (1): 95–101. doi:10.1111/j.1365-2125.1983.tb01470.x. PMC 1427833. PMID 6849751.
- Oñate SA, Tsai SY, Tsai MJ, O'Malley BW (November 1995). "Sequence and characterization of a coactivator for the steroid hormone receptor superfamily". Science. 270 (5240): 1354–7. doi:10.1126/science.270.5240.1354. PMID 7481822.
- Zhang Y, Beck CA, Poletti A, Edwards DP, Weigel NL (December 1994). "Identification of phosphorylation sites unique to the B form of human progesterone receptor. In vitro phosphorylation by casein kinase II". The Journal of Biological Chemistry. 269 (49): 31034–40. PMID 7983041.
- Mansour I, Reznikoff-Etievant MF, Netter A (August 1994). "No evidence for the expression of the progesterone receptor on peripheral blood lymphocytes during pregnancy". Human Reproduction. 9 (8): 1546–9. doi:10.1093/oxfordjournals.humrep.a138746. PMID 7989520.
- Kalkhoven E, Wissink S, van der Saag PT, van der Burg B (March 1996). "Negative interaction between the RelA(p65) subunit of NF-kappaB and the progesterone receptor". The Journal of Biological Chemistry. 271 (11): 6217–24. doi:10.1074/jbc.271.11.6217. PMID 8626413.
- Wang JD, Zhu JB, Fu Y, Shi WL, Qiao GM, Wang YQ, Chen J, Zhu PD (February 1996). "Progesterone receptor immunoreactivity at the maternofetal interface of first trimester pregnancy: a study of the trophoblast population". Human Reproduction. 11 (2): 413–9. doi:10.1093/humrep/11.2.413. PMID 8671234.
- Thénot S, Henriquet C, Rochefort H, Cavaillès V (May 1997). "Differential interaction of nuclear receptors with the putative human transcriptional coactivator hTIF1". The Journal of Biological Chemistry. 272 (18): 12062–8. doi:10.1074/jbc.272.18.12062. PMID 9115274.
- Jenster G, Spencer TE, Burcin MM, Tsai SY, Tsai MJ, O'Malley BW (July 1997). "Steroid receptor induction of gene transcription: a two-step model". Proceedings of the National Academy of Sciences of the United States of America. 94 (15): 7879–84. doi:10.1073/pnas.94.15.7879. PMC 21523. PMID 9223281.
- Shanker YG, Sharma SC, Rao AJ (September 1997). "Expression of progesterone receptor mRNA in the first trimester human placenta". Biochemistry and Molecular Biology International. 42 (6): 1235–40. doi:10.1080/15216549700203701. PMID 9305541.
- Richer JK, Lange CA, Wierman AM, Brooks KM, Tung L, Takimoto GS, Horwitz KB (April 1998). "Progesterone receptor variants found in breast cells repress transcription by wild-type receptors". Breast Cancer Research and Treatment. 48 (3): 231–41. doi:10.1023/A:1005941117247. PMID 9598870.
- Williams SP, Sigler PB (May 1998). "Atomic structure of progesterone complexed with its receptor". Nature. 393 (6683): 392–6. doi:10.1038/30775. PMID 9620806.
- Boonyaratanakornkit V, Melvin V, Prendergast P, Altmann M, Ronfani L, Bianchi ME, Taraseviciene L, Nordeen SK, Allegretto EA, Edwards DP (August 1998). "High-mobility group chromatin proteins 1 and 2 functionally interact with steroid hormone receptors to enhance their DNA binding in vitro and transcriptional activity in mammalian cells". Molecular and Cellular Biology. 18 (8): 4471–87. doi:10.1128/mcb.18.8.4471. PMC 109033. PMID 9671457.
- Nawaz Z, Lonard DM, Smith CL, Lev-Lehman E, Tsai SY, Tsai MJ, O'Malley BW (February 1999). "The Angelman syndrome-associated protein, E6-AP, is a coactivator for the nuclear hormone receptor superfamily". Molecular and Cellular Biology. 19 (2): 1182–9. doi:10.1128/mcb.19.2.1182. PMC 116047. PMID 9891052.
External links
- Progesterone+Receptors at the US National Library of Medicine Medical Subject Headings (MeSH)
PDB gallery | |
|---|---|
|