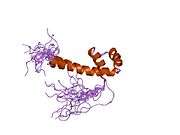
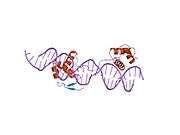
PAX6
Paired box protein Pax-6, also known as aniridia type II protein (AN2) or oculorhombin, is a protein that in humans is encoded by the PAX6 gene.[4]
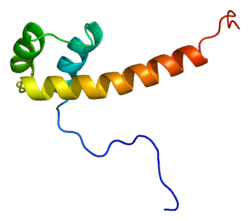
Function
-_Drosophila_Model.jpg)
PAX6 is a member of the Pax gene family which is responsible for carrying the genetic information that will encode the Pax-6 protein. It acts as a "master control" gene for the development of eyes and other sensory organs, certain neural and epidermal tissues as well as other homologous structures, usually derived from ectodermal tissues. However, it has been recognized that a suite of genes is necessary for eye development, and therefore the term of "master control" gene may be inaccurate.[5] Pax-6 is expressed as a transcription factor when neural ectoderm receives a combination of weak Sonic hedgehog (SHH) and strong TGF-Beta signaling gradients. Expression is first seen in the forebrain, hindbrain, head ectoderm and spinal cord followed by later expression in midbrain. This transcription factor is most noted for its use in the interspecifically induced expression of ectopic eyes and is of medical importance because heterozygous mutants produce a wide spectrum of ocular defects such as Aniridia in humans.[6]
Pax6 serves as a regulator in the coordination and pattern formation required for differentiation and proliferation to successfully take place, ensuring that the processes of neurogenesis and oculogenesis are carried out successfully. As a transcription factor, Pax6 acts at the molecular level in the signaling and formation of the central nervous system. The characteristic paired DNA binding domain of Pax6 utilizes two DNA-binding domains, the paired domain (PD), and the paired-type homeodomain (HD). These domains function separately via utilization by Pax6 to carry out molecular signaling that regulates specific functions of Pax6. An example of this lies in HD's regulatory involvement in the formation of the lens and retina throughout oculogenesis contrasted by the molecular mechanisms of control exhibited on the patterns of neurogenesis in brain development by PD. The HD and PD domains act in close coordination, giving Pax6 its multifunctional nature in directing molecular signaling in formation of the CNS. Although many functions of Pax6 are known, the molecular mechanisms of these functions remain largely unresolved.[7] High-throughput studies uncovered many new target genes of the Pax6 transcription factors during lens development.[8] They include the transcriptional activator BCL9, recently identified, together with Pygo2, to be downstream effectors of Pax6 functions.[9]
Species distribution

PAX6 protein function is highly conserved across bilaterian species. For instance, mouse PAX6 can trigger eye development in Drosophila melanogaster. Additionally, mouse and human PAX6 have identical amino acid sequences.[10]
Genomic organisation of the PAX6 locus varies among species, including the number and distribution of exons, cis-regulatory elements, and transcription start sites,[11][12] although most elements at the Vertebrata clade do line up with each other.[13][14] The first work on genomic organisation was performed in quail, but the picture of the mouse locus is the most complete to date. This consists of 3 confirmed promoters (P0, P1, Pα), 16 exons, and at least 6 enhancers. The 16 confirmed exons are numbered 0 through 13 with the additions of exon α located between exons 4 and 5, and the alternatively spliced exon 5a. Each promoter is associated with its own proximal exon (exon 0 for P0, exon 1 for P1) resulting in transcripts which are alternatively spliced in the 5' un-translated region.[15] By convention, exon for orthologs from other species are named relative to the human/mouse numbering, as long as the organization is reasonably well-conserved.[14]
Of the four Drosophila Pax6 orthologues, it is thought that the eyeless (ey) and twin of eyeless (toy) gene products share functional homology with the vertebrate canonical Pax6 isoform, while the eyegone (eyg) and twin of eyegone (toe) gene products share functional homology with the vertebrate Pax6(5a) isoform. Eyeless and eyegone were named for their respective mutant phenotypes. These paralogs also play a role in the development in the entire eye-antennal disc, and consequently in head formation.[16] toy positively regulates ey expression.[17]
Isoforms
The vertebrate PAX6 locus encodes at least three different protein isoforms, these being the canonical PAX6, PAX6(5a), and PAX6(ΔPD). The canonical PAX6 protein contains an N-terminal paired domain, connected by a linker region to a paired-type homeodomain, and a proline/serine/threonine (P/S/T)-rich C-terminal domain. The paired domain and paired-type homeodomain each have DNA binding activities, while the P/S/T-rich domain possesses a transactivation function. PAX6(5a) is a product of the alternatively spliced exon 5a resulting in a 14 residue insertion in the paired domain which alters the specificity of this DNA binding activity. The nucleotide sequence corresponding to the linker region encodes a set of three alternative translation start codons from which the third PAX6 isoform originates. Collectively known as the PAX6(ΔPD) or pairedless isoforms, these three gene products all lack a paired domain. The pairedless proteins possess molecular weights of 43, 33, or 32kDa, depending on the particular start codon used. PAX6 transactivation function is attributed to the variable length C-terminal P/S/T-rich domain which stretches to 153 residues in human and mouse proteins.
Clinical significance
Experiments in mice demonstrate that a deficiency in Pax-6 leads to decrease in brain size, brain structure abnormality leading to Autism, lack of iris formation or a thin cornea. Knockout experiments produced eyeless phenotypes reinforcing indications of the gene's role in eye development.[6]
Mutations
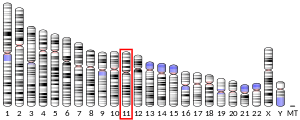
During embryological development the PAX6 gene, found on chromosome 2, can be seen expressed in multiple early structures such as the spinal cord, hindbrain, forebrain and eyes.[18] Mutations of the PAX6 gene in mammalian species can produce a drastic effect on the phenotype of the organism. This can be seen in mice that contain homozygous mutations of the 422 amino acid long transcription factor encoded by PAX6 in which they do not develop eyes or nasal cavities termed ‘small eye’ mice (PAX10sey/sey).[18][19] Deletion of PAX6 induces the same abnormal phenotypes indicating that mutations cause the protein to lose functionality. PAX6 is essential is the formation of the retina, lens and cornea due to its role in early cell determination when forming precursors of these structures such as the optic vesicle and overlying surface ectoderm.[19] PAX10 mutations also hinder nasal cavity development due to the similar precursor structures that in small eye mice do not express PAX10 mRNA.[20] Mice lacking any functional pax6 begin to be phenotypically differentiable from normal mouse embryos at about day 9 to 10 of gestation.[21] The full elucidation of the precise mechanisms and molecular components by which the PAX6 gene influences eye, nasal and central nervous system development are still researched however, the study of PAX6 has brought more understanding to the development and genetic complexities of these mammalian body systems.
References
- GRCh38: Ensembl release 89: ENSG00000007372 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Jordan T, Hanson I, Zaletayev D, Hodgson S, Prosser J, Seawright A, Hastie N, van Heyningen V (August 1992). "The human PAX6 gene is mutated in two patients with aniridia". Nature Genetics. 1 (5): 328–32. doi:10.1038/ng0892-328. PMID 1302030.
- Fernald RD (2004). "Eyes: variety, development and evolution". Brain, Behavior and Evolution. 64 (3): 141–7. doi:10.1159/000079743. PMID 15353906.
- Davis LK, Meyer KJ, Rudd DS, Librant AL, Epping EA, Sheffield VC, Wassink TH (May 2008). "Pax6 3' deletion results in aniridia, autism and mental retardation". Human Genetics. 123 (4): 371–8. doi:10.1007/s00439-008-0484-x. PMC 2719768. PMID 18322702.
- Walcher T, Xie Q, Sun J, Irmler M, Beckers J, Öztürk T, Niessing D, Stoykova A, Cvekl A, Ninkovic J, Götz M (March 2013). "Functional dissection of the paired domain of Pax6 reveals molecular mechanisms of coordinating neurogenesis and proliferation". Development. 140 (5): 1123–36. doi:10.1242/dev.082875. PMC 3583046. PMID 23404109.
- Sun J, Rockowitz S, Xie Q, Ashery-Padan R, Zheng D, Cvekl A (August 2015). "Identification of in vivo DNA-binding mechanisms of Pax6 and reconstruction of Pax6-dependent gene regulatory networks during forebrain and lens development". Nucleic Acids Research. 43 (14): 6827–46. doi:10.1093/nar/gkv589. PMC 4538810. PMID 26138486.
- Cantù C, Zimmerli D, Hausmann G, Valenta T, Moor A, Aguet M, Basler K (September 2014). "Pax6-dependent, but β-catenin-independent, function of Bcl9 proteins in mouse lens development". Genes & Development. 28 (17): 1879–84. doi:10.1101/gad.246140.114. PMC 4197948. PMID 25184676.
- Gehring WJ, Ikeo K (September 1999). "Pax 6: mastering eye morphogenesis and eye evolution". Trends in Genetics. 15 (9): 371–7. doi:10.1016/S0168-9525(99)01776-X. PMID 10461206.
- Irvine SQ, Fonseca VC, Zompa MA, Antony R (May 2008). "Cis-regulatory organization of the Pax6 gene in the ascidian Ciona intestinalis". Developmental Biology. 317 (2): 649–59. doi:10.1016/j.ydbio.2008.01.036. PMC 2684816. PMID 18342846.
- Fabian P, Kozmikova I, Kozmik Z, Pantzartzi CN (2015). "Pax2/5/8 and Pax6 alternative splicing events in basal chordates and vertebrates: a focus on paired box domain". Frontiers in Genetics. 6: 228. doi:10.3389/fgene.2015.00228. PMC 4488758. PMID 26191073.
- Bhatia S, Monahan J, Ravi V, Gautier P, Murdoch E, Brenner S, van Heyningen V, Venkatesh B, Kleinjan DA (March 2014). "A survey of ancient conserved non-coding elements in the PAX6 locus reveals a landscape of interdigitated cis-regulatory archipelagos". Developmental Biology. 387 (2): 214–28. doi:10.1016/j.ydbio.2014.01.007. PMID 24440152.
- Ravi V, Bhatia S, Gautier P, Loosli F, Tay BH, Tay A, Murdoch E, Coutinho P, van Heyningen V, Brenner S, Venkatesh B, Kleinjan DA (2013). "Sequencing of Pax6 loci from the elephant shark reveals a family of Pax6 genes in vertebrate genomes, forged by ancient duplications and divergences". PLoS Genetics. 9 (1): e1003177. doi:10.1371/journal.pgen.1003177. PMC 3554528. PMID 23359656.
- Anderson TR, Hedlund E, Carpenter EM (June 2002). "Differential Pax6 promoter activity and transcript expression during forebrain development". Mechanisms of Development. 114 (1–2): 171–5. doi:10.1016/s0925-4773(02)00051-5. PMID 12175506.
- Zhu J, Palliyil S, Ran C, Kumar JP (June 2017). "Drosophila Pax6 promotes development of the entire eye-antennal disc, thereby ensuring proper adult head formation". Proceedings of the National Academy of Sciences of the United States of America. 114 (23): 5846–5853. doi:10.1073/pnas.1610614114. PMC 5468661. PMID 28584125.
- Punzo C, Plaza S, Seimiya M, Schnupf P, Kurata S, Jaeger J, Gehring WJ (August 2004). "Functional divergence between eyeless and twin of eyeless in Drosophila melanogaster". Development. 131 (16): 3943–53. doi:10.1242/dev.01278. PMID 15253940.
- Freund C, Horsford DJ, McInnes RR (1996). "Transcription factor genes and the developing eye: a genetic perspective". Human Molecular Genetics. 5 Spec No: 1471–88. doi:10.1093/hmg/5.Supplement_1.1471. PMID 8875254.
- Walther C, Gruss P (December 1991). "Pax-6, a murine paired box gene, is expressed in the developing CNS". Development. 113 (4): 1435–49. PMID 1687460.
- Grindley JC, Davidson DR, Hill RE (May 1995). "The role of Pax-6 in eye and nasal development". Development. 121 (5): 1433–42. PMID 7789273.
- Kaufman MH, Chang HH, Shaw JP (June 1995). "Craniofacial abnormalities in homozygous Small eye (Sey/Sey) embryos and newborn mice". Journal of Anatomy. 186 (3): 607–17. PMC 1167018. PMID 7559133.
Further reading
- Callaerts P, Halder G, Gehring WJ (1997). "PAX-6 in development and evolution". Annual Review of Neuroscience. 20 (1): 483–532. doi:10.1146/annurev.neuro.20.1.483. PMID 9056723.
- Prosser J, van Heyningen V (1998). "PAX6 mutations reviewed". Human Mutation. 11 (2): 93–108. doi:10.1002/(SICI)1098-1004(1998)11:2<93::AID-HUMU1>3.0.CO;2-M. PMID 9482572.
- Hever AM, Williamson KA, van Heyningen V (June 2006). "Developmental malformations of the eye: the role of PAX6, SOX2 and OTX2". Clinical Genetics. 69 (6): 459–70. doi:10.1111/j.1399-0004.2006.00619.x. PMID 16712695.
- Glaser T, Walton DS, Maas RL (November 1992). "Genomic structure, evolutionary conservation and aniridia mutations in the human PAX6 gene". Nature Genetics. 2 (3): 232–9. doi:10.1038/ng1192-232. PMID 1345175.
- Ton CC, Hirvonen H, Miwa H, Weil MM, Monaghan P, Jordan T, van Heyningen V, Hastie ND, Meijers-Heijboer H, Drechsler M (December 1991). "Positional cloning and characterization of a paired box- and homeobox-containing gene from the aniridia region" (PDF). Cell. 67 (6): 1059–74. doi:10.1016/0092-8674(91)90284-6. hdl:2027.42/28976. PMID 1684738.
- O'Donnell FE, Pappas HR (February 1982). "Autosomal dominant foveal hypoplasia and presenile cataracts. A new syndrome". Archives of Ophthalmology. 100 (2): 279–81. doi:10.1001/archopht.1982.01030030281009. PMID 7065945.
- Martha A, Strong LC, Ferrell RE, Saunders GF (1995). "Three novel aniridia mutations in the human PAX6 gene". Human Mutation. 6 (1): 44–9. doi:10.1002/humu.1380060109. PMID 7550230.
- Hanson I, Brown A, van Heyningen V (June 1995). "A new PAX6 mutation in familial aniridia". Journal of Medical Genetics. 32 (6): 488–9. doi:10.1136/jmg.32.6.488. PMC 1050493. PMID 7666404.
- Mirzayans F, Pearce WG, MacDonald IM, Walter MA (September 1995). "Mutation of the PAX6 gene in patients with autosomal dominant keratitis". American Journal of Human Genetics. 57 (3): 539–48. PMC 1801269. PMID 7668281.
- van Heyningen V, Little PF (1995). "Report of the fourth international workshop on human chromosome 11 mapping 1994". Cytogenetics and Cell Genetics. 69 (3–4): 127–58. doi:10.1159/000133953. PMID 7698003.
- Auffray C, Behar G, Bois F, Bouchier C, Da Silva C, Devignes MD, Duprat S, Houlgatte R, Jumeau MN, Lamy B (February 1995). "[IMAGE: molecular integration of the analysis of the human genome and its expression]". Comptes Rendus de l'Académie des Sciences, Série III. 318 (2): 263–72. PMID 7757816.
- Martha A, Ferrell RE, Mintz-Hittner H, Lyons LA, Saunders GF (May 1994). "Paired box mutations in familial and sporadic aniridia predicts truncated aniridia proteins". American Journal of Human Genetics. 54 (5): 801–11. PMC 1918271. PMID 7909985.
- Glaser T, Jepeal L, Edwards JG, Young SR, Favor J, Maas RL (August 1994). "PAX6 gene dosage effect in a family with congenital cataracts, aniridia, anophthalmia and central nervous system defects". Nature Genetics. 7 (4): 463–71. doi:10.1038/ng0894-463. PMID 7951315.
- Epstein JA, Glaser T, Cai J, Jepeal L, Walton DS, Maas RL (September 1994). "Two independent and interactive DNA-binding subdomains of the Pax6 paired domain are regulated by alternative splicing". Genes & Development. 8 (17): 2022–34. doi:10.1101/gad.8.17.2022. PMID 7958875.
- Davis A, Cowell JK (December 1993). "Mutations in the PAX6 gene in patients with hereditary aniridia". Human Molecular Genetics. 2 (12): 2093–7. doi:10.1093/hmg/2.12.2093. PMID 8111379.
- Hanson IM, Fletcher JM, Jordan T, Brown A, Taylor D, Adams RJ, Punnett HH, van Heyningen V (February 1994). "Mutations at the PAX6 locus are found in heterogeneous anterior segment malformations including Peters' anomaly". Nature Genetics. 6 (2): 168–73. doi:10.1038/ng0294-168. PMID 8162071.
- Hanson IM, Seawright A, Hardman K, Hodgson S, Zaletayev D, Fekete G, van Heyningen V (July 1993). "PAX6 mutations in aniridia". Human Molecular Genetics. 2 (7): 915–20. doi:10.1093/hmg/2.7.915. PMID 8364574.
- Azuma N, Nishina S, Yanagisawa H, Okuyama T, Yamada M (June 1996). "PAX6 missense mutation in isolated foveal hypoplasia". Nature Genetics. 13 (2): 141–2. doi:10.1038/ng0696-141. PMID 8640214.
External links
- PAX6+protein at the US National Library of Medicine Medical Subject Headings (MeSH)
- GeneReviews/NCBI/NIH/UW entry on Anophthalmia / Microphthalmia Overview
- GeneReviews/NCBI/NIH/UW entry on Aniridia
- OMIM entries on Aniridia
- Gene Expression Patterns from the Allen Brain Atlases
- Overview of all the structural information available in the PDB for UniProt: P26367 (Paired box protein Pax-6) at the PDBe-KB.
PDB gallery | |
|---|---|
|