Genetic assimilation
Genetic assimilation is a process described by Conrad H. Waddington by which a phenotype originally produced in response to an environmental condition, such as exposure to a teratogen, later becomes genetically encoded via artificial selection or natural selection. Despite superficial appearances, this does not require the (Lamarckian) inheritance of acquired characters, although epigenetic inheritance could potentially influence the result.[1] Waddington stated that genetic assimilation overcomes the barrier to selection imposed by what he called canalization of developmental pathways; he supposed that the organism's genetics evolved to ensure that development proceeded in a certain way regardless of normal environmental variations.
The classic example of genetic assimilation was a pair of experiments in 1942 and 1953 by Waddington. He exposed Drosophila fruit fly embryos to ether, producing an extreme change in their phenotype: they developed a double thorax, resembling the effect of the bithorax gene. This is called a homeotic change. Flies which developed halteres (the modified hindwings of true flies, used for balance) with wing-like characteristics were chosen for breeding for 20 generations, by which point the phenotype could be seen without other treatment.[2]
Waddington's explanation has been controversial, and has been accused of being Lamarckian. More recent evidence appears to confirm the existence of genetic assimilation in evolution; in yeast, when a stop codon is lost by mutation, the reading frame is preserved much more often than would be expected.
History
Waddington's experiments
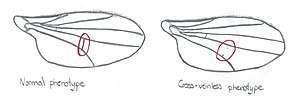
Conrad H. Waddington's classic experiment (1942) induced an extreme environmental reaction in the developing embryos of Drosophila. In response to ether vapor, a proportion of embryos developed a radical phenotypic change, a second thorax. At this point in the experiment bithorax is not innate; it is induced by an unusual environment. Waddington then repeatedly selected Drosophila for the bithorax phenotype over some 20 generations. After this time, some Drosophila developed bithorax without the ether treatment.[3]
Waddington carried out a similar experiment in 1953, this time inducing the cross-veinless phenocopy in Drosophila with a heat shock, with 40% of the flies showing the phenotype prior to selection. Again he selected for the phenotype over several generations, applying heat shock each time, and eventually the phenotype appeared even without heat shock.[4][5]
Waddington's explanation
Waddington called the effect he had seen "genetic assimilation". His explanation was that it was caused by a process he called "canalization". He compared embryonic development to a ball rolling down a slope in what he called an epigenetic landscape, where each point on the landscape is a possible state of the organism (involving many variables). As a particular pathway becomes entrenched or "canalized", it becomes more stable, likely to occur even in the face of environmental changes. Major perturbations such as ether or heat shock eject the developmental pathway from the metaphorical canal, exploring other parts of the epigenetic landscape. Selection in the presence of that perturbation leads to the evolution of a new canal; after the perturbation is discontinued, developmental trajectories continue to follow the canalized pathway.[5]
A Darwinian explanation
Other evolutionary biologists have agreed that assimilation occurs, but give a different, purely quantitative genetics explanation in terms of Darwin's natural or artificial selection. The phenotype, say cross-veinless, is presumed to be caused by a combination of multiple genes. The phenotype appears when the sum of gene effects exceeds a threshold; if that threshold is lowered by the perturbation, the phenotype is more likely to be seen. Continued selection under perturbing conditions increases the frequency of the alleles of genes that promote the phenotype until the threshold is breached, and the phenotype appears without requiring the heat shock.[5][6]
Perturbations can be genetic or epigenetic rather than environmental. For example, Drosophila fruit flies have a heat shock protein, Hsp90, which protects the development of many structures in the adult fly from heat shock. If the protein is damaged by a mutation, then just as if it were damaged by the environmental effects of drugs, many different phenotypic variants appear; if these are selected for, they quickly establish without further need for the mutant Hsp90.[7]
Neo-Darwinism or Lamarckism
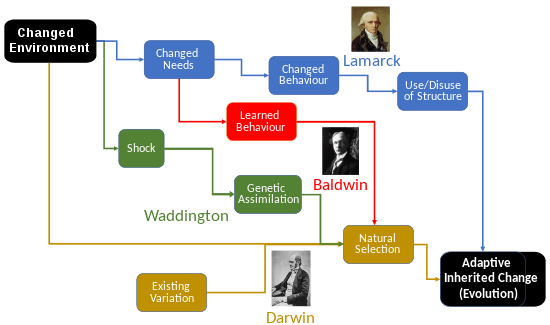
Waddington's theory of genetic assimilation was controversial.[8] The evolutionary biologists Theodosius Dobzhansky and Ernst Mayr both thought that Waddington was using genetic assimilation to support so-called Lamarckian inheritance. They denied that the inheritance of acquired characteristics had taken place, and asserted that Waddington had simply observed the natural selection of genetic variants that already existed in the study population.[9] Waddington himself interpreted his results in a Neo-Darwinian way, particularly emphasizing that they "could bring little comfort to those who wish to believe that environmental influences tend to produce heritable changes in the direction of adaptation."[1][10][11] The evolutionary developmental biologist Adam S. Wilkins wrote that "[Waddington] in his lifetime... was widely perceived primarily as a critic of Neo-Darwinian evolutionary theory. His criticisms ... were focused on what he saw as unrealistic, 'atomistic' models of both gene selection and trait evolution." In particular, according to Wilkins, Waddington felt that the Neo-Darwinians badly neglected the phenomenon of extensive gene interactions and that the 'randomness' of mutational effects, posited in the theory, was false.[12] Even though Waddington became critical of the neo-Darwinian synthetic theory of evolution, he still described himself as a Darwinian, and called for an extended evolutionary synthesis based on his research.[13] Waddington did not deny the threshold-based conventional genetic interpretation of his experiments, but regarded it "as a told to the children version of what I wished to say" and considered the debate to be about "mode of expression, rather than of substance".[14] Both genetic assimilation and the related Baldwin effect are theories of phenotypic plasticity, where aspects of an organism's physiology and behaviour are affected by the environment. The evolutionary ecologist Erika Crispo states that they differ in that genetic assimilation decreases the level of plasticity (returning to Waddington's original definition of canalization; whereas the Baldwin effect may increase it) but does not change the mean phenotypic value (where the Baldwin effect changes it).[15] Crispo defines genetic assimilation as a kind of genetic accommodation, "evolution in response to both genetically based and environmentally induced novel traits",[15] which in turn is in her view central to the Baldwin effect.[15]
Relationship to adaptation
Mathematical modeling suggests that under certain circumstances, natural selection favours the evolution of canalization that is designed to fail under extreme conditions.[16][17] If the result of such a failure is favoured by natural selection, genetic assimilation occurs. In the 1960s, Waddington and his colleague the animal geneticist J. M. Rendel argued for the importance of genetic assimilation in natural adaptation, as a means of providing new and potentially beneficial variation to populations under stress, enabling them to evolve rapidly.[18][19] Their contemporary George C. Williams argued that genetic assimilation proceeds at the cost of a loss of previously adaptive developmental plasticity, and therefore should be seen as resulting in a net loss rather than gain of complexity, making it in his view uninteresting from the perspective of the constructive process of adaptation.[20] However, the preceding phenotypic plasticity need not be adaptive, but simply represent a breakdown of canalization.[16]
In natural populations

Several instances of genetic assimilation have been documented contributing to natural selection in the wild. For example, populations of the island tiger snakes (Notechis scutatus) have become isolated on islands and have larger heads to cope with large prey animals. Young populations have larger heads by phenotypic plasticity, whereas large heads have become genetically assimilated in older populations.[21]
In another example, patterns of left-right asymmetry or "handedness", when present, can be determined either genetically or plastically. During evolution, genetically determined directional asymmetry, as in the left-oriented human heart, can arise either from a nonheritable (phenotypic) developmental process, or directly by mutation from a symmetric ancestor. An excess of transitions from plastically determined to genetically determined handedness points to the role of genetic assimilation in evolution.[22]
A third example has been seen in yeast. Evolutionary events in which stop codons are lost preserve the reading frame much more often than would be expected from mutation bias. This finding is consistent with the role of the yeast prion [PSI+] in epigenetically facilitating stop codon readthrough, followed by genetic assimilation via the permanent loss of the stop codon.[23]
See also
- Evolutionary developmental biology
- List of genetics-related topics
References
- Pocheville, Arnaud; Danchin, Etienne (January 1, 2017). "Chapter 3: Genetic assimilation and the paradox of blind variation". In Huneman, Philippe; Walsh, Denis (eds.). Challenging the Modern Synthesis. Oxford University Press.
- Gilbert, Scott F. (1991). "Induction and the Origins of Developmental Genetics". A Conceptual History of Modern Embryology. Plenum Press. pp. 181–206. ISBN 978-0306438424.
- Waddington, C. H. (1942). "Canalization of development and the inheritance of acquired characters". Nature. 150 (3811): 563–565. doi:10.1038/150563a0.
- Waddington, C. H. (1953). "Genetic Assimilation of an Acquired Character". Evolution. 7 (2): 118–126. doi:10.2307/2405747. JSTOR 2405747.
- Masel, Joanna (2004). "Genetic assimilation can occur in the absence of selection for the assimilating phenotype, suggesting a role for the canalization heuristic" (PDF). Journal of Evolutionary Biology. 17 (5): 1106–1110. doi:10.1111/j.1420-9101.2004.00739.x. PMID 15312082.
- Falconer, D. S.; Mackay, Trudy F. C. (1998). Introduction to quantitative genetics (4th ed.). Longman. pp. 309–310. ISBN 978-0-582-24302-6.
- Rutherford, S. L.; Lindquist, S. (1998). "Hsp90 as a capacitor for morphological evolution". Nature. 396 (6709): 336–342. doi:10.1038/24550. PMID 9845070.
- Pigliucci, M. (2006). "Phenotypic plasticity and evolution by genetic assimilation". Journal of Experimental Biology. 209 (12): 2362–2367. doi:10.1242/jeb.02070. ISSN 0022-0949.
- Gilbert, Scott F. (2013). A Conceptual History of Modern Embryology: Volume 7: A Conceptual History of Modern Embryology. Springer. p. 205. ISBN 978-1-4615-6823-0.
- Waddington, Conrad (1953). "Genetic assimilation of an acquired character". Evolution. 7 (2): 118–126. doi:10.2307/2405747. JSTOR 2405747.
- Noble, Denis (2015). "Conrad Waddington and the origin of epigenetics". Journal of Experimental Biology. 218 (6): 816–818. doi:10.1242/jeb.120071. PMID 25788723.
- Wilkins, Adam S. (2015). "Waddington's Unfinished Critique of Neo-Darwinian Genetics: Then and Now". Biological Theory. 3 (3): 224–232. doi:10.1162/biot.2008.3.3.224. ISSN 1555-5542.
- Huang, Sui (2012). "The molecular and mathematical basis of Waddington's epigenetic landscape: A framework for post-Darwinian biology?". BioEssays. 34 (2): 149–157. doi:10.1002/bies.201100031. ISSN 0265-9247.
- Waddington, C. H. (1 November 1958). "Comment on Professor Stern's Letter". The American Naturalist. 92 (867): 375–376. doi:10.1086/282049. ISSN 0003-0147.
- Crispo, Erika (2007). "The Baldwin effect and genetic assimilation: revisiting two mechanisms of evolutionary change mediated by phenotypic plasticity". Evolution. 61 (11): 2469–2479. doi:10.1111/j.1558-5646.2007.00203.x. PMID 17714500.
- Eshel, I.; Matessi, C. (August 1998). "Canalization, genetic assimilation and preadaptation. A quantitative genetic model". Genetics. 149 (4): 2119–2133. PMC 1460279. PMID 9691063.
- Pigliucci, Massimo; Murren, C. J. (July 2003). "Perspective: Genetic assimilation and a possible evolutionary paradox: can macroevolution sometimes be so fast as to pass us by?". Evolution. 57 (7): 1455–64. doi:10.1111/j.0014-3820.2003.tb00354.x. PMID 12940351.
- Hall, Brian K. (1992). "Waddington's Legacy in Development and Evolution" (PDF). American Zoologist. 32 (1): 113–122. doi:10.1093/icb/32.1.113. JSTOR 3883742.
- Rendel, J. M. (1968). R. C. Lewinton (ed.). Genetic control of developmental processes. Population biology and evolution. Syracuse University Press. pp. 47–68.
- Williams, George Christopher (2008). Adaptation and Natural Selection: A Critique of Some Current Evolutionary Thought. Princeton University Press. pp. 71–77. ISBN 978-1-4008-2010-8.
- Aubret, F.; Shine, R. (2009). "Genetic assimilation and the postcolonisation erosion of phenotypic plasticity in island Tiger snakes" (PDF). Current Biology. 19 (22): 1932–1936. doi:10.1016/j.cub.2009.09.061. PMID 19879141.
- Palmer, A. Richard (2004). "Symmetry breaking and the evolution of development". Science. 306 (5697): 828–833. CiteSeerX 10.1.1.631.4256. doi:10.1126/science.1103707. PMID 15514148.
- Giacomelli, M. G.; Hancock, A. S.; Masel, Joanna (13 November 2006). "The Conversion of 3' UTRs into Coding Regions". Molecular Biology and Evolution. 24 (2): 457–464. doi:10.1093/molbev/msl172. PMC 1808353. PMID 17099057.
