PER1
The PER1 gene encodes the period circadian protein homolog 1 protein in humans.[5]




Function
The PER1 protein is important to the maintenance of circadian rhythms in cells, and may also play a role in the development of cancer. This gene is a member of the period family of genes. It is expressed with a daily oscillating circadian rhythm, or an oscillation that cycles with a period of approximately 24 hours. PER1 is most notably expressed in the region of the brain called the suprachiasmatic nucleus (SCN), which is the primary circadian pacemaker in the mammalian brain. PER1 is also expressed throughout mammalian peripheral tissues.[6] Genes in this family encode components of the circadian rhythms of locomotor activity, metabolism, and behavior. Circadian expression of PER1 in the suprachiasmatic nucleus will free-run in constant darkness, meaning that the 24-hour period of the cycle will persist without the aid of external light cues. Subsequently, a shift in the light/dark cycle evokes a proportional shift of gene expression in the suprachiasmatic nucleus. The time of gene expression is sensitive to light, as light during a mammal's subjective night results in a sudden increase in per expression and thus a shift in phase in the suprachiasmatic nucleus.[7] Alternative splicing has been observed in this gene; however, these variants have not been fully described.[8] There is some disagreement between experts over the occurrence of polymorphisms with functional significance. Many scientists state that there are no known polymorphisms of the human PER1 gene with significance at a population level that results in measurable behavioral or physiological changes.[9] Still, some believe that even silent mutations can cause significant behavioral phenotypes,and result in major phase changes.[10]
Functional conservation of the PER gene is shown in a study by Shigeyoshi et al. 2002. In this study, mouse mPer1 and mPer2 genes were driven by Drosophila timeless promoter in Drosophila melanogaster. They found that both mPer constructs could restore rhythm to arrythmic flies (per01 flies). Thus mPer1 and mPer2 can function as clock components in flies and may have implications concerning the homology of per genes.[11]
Role in chronobiology
The PER1 gene, also called rigui, is a characteristic circadian oscillator. PER1 is rhythmically transcribed in the SCN, keeping a period of approximately 24 hours. This rhythm is sustained in constant darkness, and can also be entrained to changing light cycles.[5] PER1 is involved in generating circadian rhythms in the SCN, and also has an effect on other oscillations throughout the body. For example, PER1 knockouts affect food entrainable oscillators and methamphetamine-sensitive circadian oscillators, whose periods are altered in the absence of PER1.[12] In addition, mice with knockouts in both the PER1 and PER2 genes show no circadian rhythmicity.[13] Phase shifts in PER1 neurons can be induced by a strong, brief light stimulus to the SCN of rats. This light exposure causes increases in PER1 mRNA, suggesting that the PER1 gene plays an important role in entrainment of the mammalian biological clock to the light-dark cycle.[14]
Feedback mechanism
The PER1 mRNA is expressed in all cells, acting as a part of a transcription-translation negative feedback mechanism, which creates a cell autonomous molecular clock. PER1 transcription is regulated by protein interactions with its five E-box and one D-box elements in its promoter region. Heterodimer CLOCK-BMAL1 activates E-box elements present in the PER1 promoter, as well activating the E box promoters of other components of the molecular clock such as PER2, CRY1, and CRY2. The phase of PER1 mRNA expression varies between tissues,[15] The transcript leaves the nucleus and is translated into a protein with PAS domains, which enable protein-protein interactions. PER1 and PER2 are phosphorylated by CK1ε, which leads to increased ubiquitylation and degradation.[16] This phosphorylation is counteracted by PP1 phosphatase, resulting in a more gradual increase in phosphorylated PER, and an additional control over the period of the molecular clock.[17] Phosphorylation of PER1 can also lead to masking of its leucine-rich nuclear localization sequence and thus impeded heterodimer import.[18]
PER interacts with other PER proteins as well as the E-box regulated, clock controlled proteins CRY1 and CRY2 to create a heterodimer which translocates into the nucleus. There it inhibits CLOCK-BMAL activation.[19] PER1 is not necessary for the creation circadian rhythms, but homozygous PER1 mutants display a shortened period of mRNA expression.[13] While PER1 must be mutated in conjunction with PER2 to result in arhythmiticity, the two translated PER proteins have been shown to have slightly different roles, as PER1 acts preferentially through interaction with other clock proteins.[20]
Clinical significance
PER1 expression may have significant effects on the cell cycle. Cancer is often a result of unregulated cell growth and division, which can be controlled by circadian mechanisms. Therefore, a cell's circadian clock may play a large role in its likelihood of developing into a cancer cell. PER1 is a gene that plays an important role in such a circadian mechanism. Its overexpression, in particular, causes DNA-damage induced apoptosis. In addition, down-regulation of PER1 can enhance tumor growth in mammals.[21] PER1 also interacts with proteins ATM and Chk2. These proteins are key checkpoint proteins in the cell cycle.[22] Cancer patients have a lowered expression of per1. Gery, et al. suggests that regulation of PER1 expression may be useful for cancer treatment in the future.[23]
Gene
Orthologs
The following is a list of some orthologs of the PER1 gene in other species:[24]
- PER1 (Rattus norvegicus)
- PER1 (Mus musculus)
- per1a (Danio rerio)
- PER1 (Homo sapiens)
- lin-42 (Caenorhabditis elegans)
- PER1 (Bos taurus)
- per1b (Danio rerio)
- PER (Drosophila melanogaster)
- PER1 (Xenopus tropicalis)
- PER1 (Equus caballus)
- PER1 (Macaca mulatta)
- PER1 (Sus scrofa)
Location
The human PER1 gene is located on chromosome 17 at the following location:[25]
- Start: 8,140,470
- Finish: 8,156,405
- Length: 15,936
- Exons: 24
PER1 has 19 transcripts (splice variants).
Discovery
The PER1 ortholog was first discovered by Ronald Konopka and Seymour Benzer in 1971. During 1997, Period 1 (mPer1) and Period 2 (mPer2) genes were discovered (Sun et al., 1997 and Albretch et al., 1997). Through homology screens with the Drosophila per, these genes were discovered. It was independently discovered by Sun et al. 1997, naming it RIGUI and by Tei et al. 1997, who named it hper because of the protein sequence similarity with Drosophila per. They found that the mouse homolog had the properties of a circadian regulator. It had circadian expression in the suprachiasmatic nucleus (SCN), self-sustained oscillation, and entrainment of circadian expression by external light cues.[26]
References
- GRCh38: Ensembl release 89: ENSG00000179094 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000020893 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Sun ZS, Albrecht U, Zhuchenko O, Bailey J, Eichele G, Lee CC (September 1997). "RIGUI, a putative mammalian ortholog of the Drosophila period gene". Cell. 90 (6): 1003–11. doi:10.1016/S0092-8674(00)80366-9. PMID 9323128.
- Lamont EW, James FO, Boivin DB, Cermakian N (September 2007). "From circadian clock gene expression to pathologies". Sleep Med. 8 (6): 547–56. doi:10.1016/j.sleep.2006.11.002. PMID 17395534.
- Ebling FJ (October 1996). "The role of glutamate in the photic regulation of the suprachiasmatic nucleus". Prog. Neurobiol. 50 (2–3): 109–32. doi:10.1016/S0301-0082(96)00032-9. PMID 8971980.
- EntrezGene 5187
- Lim AS, Chang AM, Shulman JM, Raj T, Chibnik LB, Cain SW, Rothamel K, Benoist C, Myers AJ, Czeisler CA, Buchman AS, Bennett DA, Duffy JF, Saper CB, De Jager PL (September 2012). "A common polymorphism near PER1 and the timing of human behavioral rhythms". Ann. Neurol. 72 (3): 324–34. doi:10.1002/ana.23636. PMC 3464954. PMID 23034908.
- Carpen JD, von Schantz M, Smits M, Skene DJ, Archer SN (2006). "A silent polymorphism in the PER1 gene associates with extreme diurnal preference in humans". J. Hum. Genet. 51 (12): 1122–25. doi:10.1007/s10038-006-0060-y. PMID 17051316.
- Shigeyoshi Y, Meyer-Bernstein E, Yagita K, Fu W, Chen Y, Takumi T, Schotland P, Sehgal A, Okamura H (February 2002). "Restoration of circadian behavioural rhythms in a period null Drosophila mutant (per01) by mammalian period homologues mPer1 and mPer2". Genes Cells. 7 (2): 163–71. doi:10.1046/j.1356-9597.2001.00503.x. PMID 11895480.
- Pendergast JS, Oda GA, Niswender KD, Yamazaki S (August 2012). "Period determination in the food-entrainable and methamphetamine-sensitive circadian oscillator(s)". Proc. Natl. Acad. Sci. U.S.A. 109 (35): 14218–23. doi:10.1073/pnas.1206213109. PMC 3435193. PMID 22891330.
- Zheng B, Albrecht U, Kaasik K, Sage M, Lu W, Vaishnav S, Li Q, Sun ZS, Eichele G, Bradley A, Lee CC (June 2001). "Nonredundant roles of the mPer1 and mPer2 genes in the mammalian circadian clock". Cell. 105 (5): 683–94. doi:10.1016/S0092-8674(01)00380-4. PMID 11389837.
- Miyake S, Sumi Y, Yan L, Takekida S, Fukuyama T, Ishida Y, Yamaguchi S, Yagita K, Okamura H (November 2000). "Phase-dependent responses of Per1 and Per2 genes to a light-stimulus in the suprachiasmatic nucleus of the rat". Neurosci. Lett. 294 (1): 41–44. doi:10.1016/S0304-3940(00)01545-7. PMID 11044582.
- Yamajuku D, Shibata Y, Kitazawa M, Katakura T, Urata H, Kojima T, Nakata O, Hashimoto S (December 2010). "Identification of functional clock-controlled elements involved in differential timing of Per1 and Per2 transcription". Nucleic Acids Res. 38 (22): 7964–73. doi:10.1093/nar/gkq678. PMC 3001056. PMID 20693532.
- Keesler GA, Camacho F, Guo Y, Virshup D, Mondadori C, Yao Z (April 2000). "Phosphorylation and destabilization of human period I clock protein by human casein kinase I epsilon". NeuroReport. 11 (5): 951–55. doi:10.1097/00001756-200004070-00011. PMID 10790862.
- Lee HM, Chen R, Kim H, Etchegaray JP, Weaver DR, Lee C (September 2011). "The period of the circadian oscillator is primarily determined by the balance between casein kinase 1 and protein phosphatase 1". Proc. Natl. Acad. Sci. U.S.A. 108 (39): 16451–56. doi:10.1073/pnas.1107178108. PMC 3182690. PMID 21930935.
- Vielhaber EL, Duricka D, Ullman KS, Virshup DM (December 2001). "Nuclear export of mammalian PERIOD proteins". J. Biol. Chem. 276 (49): 45921–27. doi:10.1074/jbc.M107726200. PMC 1513547. PMID 11591712.
- Ko CH, Takahashi JS (October 2006). "Molecular components of the mammalian circadian clock". Hum. Mol. Genet. 15 Spec No 2: R271–77. doi:10.1093/hmg/ddl207. PMID 16987893.
- Bae K, Jin X, Maywood ES, Hastings MH, Reppert SM, Weaver DR (May 2001). "Differential functions of mPer1, mPer2, and mPer3 in the SCN circadian clock". Neuron. 30 (2): 525–36. doi:10.1016/S0896-6273(01)00302-6. PMID 11395012.
- Yang X, Wood PA, Ansell CM, Quiton DF, Oh EY, Du-Quiton J, Hrushesky WJ (October 2009). "The circadian clock gene Per1 suppresses cancer cell proliferation and tumor growth at specific times of day". Chronobiol. Int. 26 (7): 1323–39. doi:10.3109/07420520903431301. PMID 19916834.
- Buscemi G, Perego P, Carenini N, Nakanishi M, Chessa L, Chen J, Khanna K, Delia D (October 2004). "Activation of ATM and Chk2 kinases in relation to the amount of DNA strand breaks". Oncogene. 23 (46): 7691–700. doi:10.1038/sj.onc.1207986. PMID 15361830.
- Gery S, Komatsu N, Baldjyan L, Yu A, Koo D, Koeffler HP (May 2006). "The circadian gene per1 plays an important role in cell growth and DNA damage control in human cancer cells". Mol. Cell. 22 (3): 375–82. doi:10.1016/j.molcel.2006.03.038. PMID 16678109.
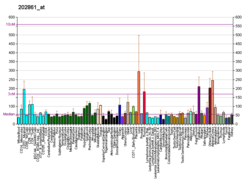
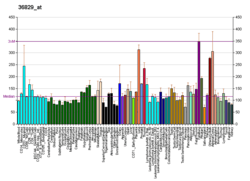
- "PER1". Expression Atlas. European Molecular Biology Laboratory. 2013. Retrieved 13 Apr 2013.
- "PER1". NCBI. 2015. Retrieved 18 Jan 2015.
- Albrecht U, Sun ZS, Eichele G, Lee CC (December 1997). "A differential response of two putative mammalian circadian regulators, mper1 and mper2, to light". Cell. 91 (7): 1055–64. doi:10.1016/S0092-8674(00)80495-X. PMID 9428527.
External links
- PER1+protein,+human at the US National Library of Medicine Medical Subject Headings (MeSH)
- More information on PER1 introns and exons
- Splice variants of PER1
This article incorporates text from the United States National Library of Medicine, which is in the public domain.