GJA1
Gap junction alpha-1 protein (GJA1), also known as connexin 43 (Cx43), is a protein that in humans is encoded by the GJA1 gene on chromosome 6.[5][6][7] As a connexin, GJA1 is a component of gap junctions, which allow for gap junction intercellular communication (GJIC) between cells to regulate cell death, proliferation, and differentiation.[8] As a result of its function, GJA1 is implicated in many biological processes, including muscle contraction, embryonic development, inflammation, and spermatogenesis, as well as diseases, including oculodentodigital dysplasia (ODDD), heart malformations, and cancers.[7][9][10]
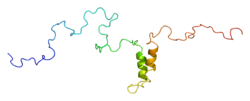


| Connexin43 | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 connexin 43 carboxyl terminal domain | |||||||||
| Identifiers | |||||||||
| Symbol | Connexin43 | ||||||||
| Pfam | PF03508 | ||||||||
| InterPro | IPR013124 | ||||||||
| TCDB | 1.A.24 | ||||||||
| |||||||||
Structure
GJA1 is a 43.0 kDa protein composed of 382 amino acids.[11] GJA1 contains a long C-terminal tail, an N-terminal domain, and multiple transmembrane domains. The protein passes through the phospholipid bilayer four times, leaving its C- and N-terminals exposed to the cytoplasm.[12] The C-terminal tail is composed of 50 amino acids and includes post-translational modification sites, as well as binding sites for transcription factors, cytoskeleton elements, and other proteins.[12][13] As a result, the C-terminal tail is central to functions such as regulating pH gating and channel assembly. Notably, the DNA region of the GJA1 gene encoding this tail is highly conserved, indicating that it is either resistant to mutations or becomes lethal when mutated. Meanwhile, the N-terminal domain is involved in channel gating and oligomerization and, thus, may control the switch between the channel's open and closed states. The transmembrane domains form the gap junction channel while the extracellular loops facilitate proper channel docking. Moreover, two extracellular loops form disulfide bonds that interact with two hexamers to form a complete gap junction channel.[12]
The connexin-43 internal ribosome entry site is an RNA element present in the 5' UTR of the mRNA of GJA1. This internal ribosome entry site (IRES) allows cap independent translation during conditions such as heat shock and stress.[14]
| Connexin-43 internal ribosome entry site (IRES) | |
|---|---|
 Predicted secondary structure and sequence conservation of IRES_Cx43 | |
| Identifiers | |
| Symbol | IRES_Cx43 |
| Rfam | RF00487 |
| Other data | |
| RNA type | Cis-reg; IRES |
| Domain(s) | Eukaryota |
| GO | 0043022 |
| SO | 0000243 |
| PDB structures | PDBe |
Function

As a member of the connexin family, GJA1 is a component of gap junctions, which are intercellular channels that connect adjacent cells to permit the exchange of low molecular weight molecules, such as small ions and secondary messengers, to maintain homeostasis.[7][12][15]
GJA1 is the most ubiquitously expressed connexin and is detected in most cell types.[7][9][12] It is the major protein in heart gap junctions and is purported to play a crucial role in the synchronized contraction of the heart.[7] Despite its key role in the heart and other vital organs, GJA1 has a short half-life (only two to four hours), indicating that the protein undergoes daily turnover in the heart and may be highly abundant or compensated with other connexins.[12] GJA1 is also largely involved in embryonic development.[7][8] For instance, transforming growth factor-beta 1 (TGF-β1) was observed to induce GJA1 expression via the Smad and ERK1/2 signaling pathways, resulting in trophoblast cell differentiation into the placenta.[8]
Furthermore, GJA1 is expressed in many immune cells, such as eosinophils and T cells, where its gap junction function promotes the maturation and activation of these cells and, by extension, the cross-communication necessary to mount an inflammatory response.[10]
In addition, GJA1 can be found in the Leydig cells and seminiferous tubules between Sertoli cells and spermatogonia or primary spermatocytes, where it plays a key role in spermatogenesis and testis development through controlling the tight junction proteins in the blood-testis barrier.
While it is a channel protein, GJA1 can also perform channel-independent functions. In the cytoplasm, the protein regulates the microtubule network and, by extension, cell migration and polarity.[9][13] This function has been observed in brain and heart development, as well as wound-healing in endothelial cells.[13] GJA1 has also been observed to localize to the mitochondria, where it promotes cell survival by downregulating the intrinsic apoptotic pathway during conditions of oxidative stress.[15]
Clinical significance
Mutations in this gene have been associated with ODDD; craniometaphyseal dysplasia; sudden infant death syndrome, which is linked to cardiac arrhythmia; Hallermann–Streiff syndrome; and heart malformations, such as viscero-atrial heterotaxia.[7][9][12][16] There have also been a few cases of reported hearing loss and skin disorders unrelated to ODDD.[12] Ultimately, GJA1 has low tolerance for deviations from its original sequence, with mutations resulting in loss- or gain-of-channel function that lead to disease phenotypes.[12] It is paradoxical, however, that patients with an array of somatic mutations in GJA1 most often do not present with cardiac arrhythmias, even though connexin-43 is the most abundant protein forming gap junctional pores in cardiomyocytes and are essential for normal action potential propagation.[17]
Notably, GJA1 expression has been associated with a wide variety of cancers, including nasopharyngeal carcinoma, meningioma, hemangiopericytoma, liver tumor, colon cancer, esophageal cancer, breast cancer, mesothelioma, glioblastoma, lung cancer, adrenocortical tumors, renal cell cancer, cervical carcinoma, ovarian carcinoma, endometrial carcinoma, prostate cancer, thyroid carcinoma, and testis cancer.[9] Its role in controlling cell motility and polarity was thought to contribute to cancer development and metastasis, though its role as a gap junction protein may also be involved.[9][15] Moreover, the cytoprotective effects of this protein can promote tumor cell survival in radiotherapy treatments, while silencing its gene increases radiosensitivity. As a result, GJA1 may serve as a target for improving the success of radiotherapeutic treatment of cancer.[15] As a biomarker, GJA1 could also be used to screen young males for risk of testis cancer.[9]
Currently, only rotigaptide, an antiarrhythmic peptide-based drug, and its derivatives, such as danegaptide, have reached clinical trials for treating cardiac pathologies by enhancing GJA1 expression. Alternatively, drugs could target complementary connexins, such as Cx40, which function similarly to GJA1. However, both approaches still require a system to target the diseased tissue to avoid inducing developmental abnormalities elsewhere.[12] Thus, a more effective approach entails designing a miRNA through antisense oligonucleotides, transfection, or infection to knock down only mutant GJA1 mRNA, thus allowing the expression of wildtype GJA1 and retaining normal phenotype.[9][12]
Interactions
Gap junction protein, alpha 1 has been shown to interact with:
- Cx37,[12]
- Cx40,[12]
- Cx45,[12]
- MAPK7,[18]
- Caveolin 1,[19]
- Tight junction protein 1[20]
- CSNK1D,[21] and
- PTPmu (PTPRM).[22]
See also
References
- GRCh38: Ensembl release 89: ENSG00000152661 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000050953 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Boyadjiev SA, Jabs EW, LaBuda M, Jamal JE, Torbergsen T, Ptácek LJ, Rogers RC, Nyberg-Hansen R, Opjordsmoen S, Zeller CB, Stine OC, Stalker HJ, Zori RT, Shapiro RE (May 1999). "Linkage analysis narrows the critical region for oculodentodigital dysplasia to chromosome 6q22-q23". Genomics. 58 (1): 34–40. doi:10.1006/geno.1999.5814. PMID 10331943.
- Fishman GI, Eddy RL, Shows TB, Rosenthal L, Leinwand LA (May 1991). "The human connexin gene family of gap junction proteins: distinct chromosomal locations but similar structures". Genomics. 10 (1): 250–256. doi:10.1016/0888-7543(91)90507-B. PMID 1646158.
- "Entrez Gene: GJA1 gap junction protein, alpha 1, 43kDa".
- Cheng JC, Chang HM, Fang L, Sun YP, Leung PC (Jul 2015). "TGF-β1 up-regulates connexin43 expression: a potential mechanism for human trophoblast cell differentiation". Journal of Cellular Physiology. 230 (7): 1558–1566. doi:10.1002/jcp.24902. PMID 25560303.
- Chevallier D, Carette D, Segretain D, Gilleron J, Pointis G (Apr 2013). "Connexin 43 a check-point component of cell proliferation implicated in a wide range of human testis diseases". Cellular and Molecular Life Sciences. 70 (7): 1207–1220. doi:10.1007/s00018-012-1121-3. PMID 22918484.
- Vliagoftis H, Ebeling C, Ilarraza R, Mahmudi-Azer S, Abel M, Adamko D, Befus AD, Moqbel R (2014). "Connexin 43 expression on peripheral blood eosinophils: role of gap junctions in transendothelial migration". BioMed Research International. 2014: 803257. doi:10.1155/2014/803257. PMC 4109672. PMID 25110696.
- "Protein sequence of human GJA1 (Uniprot ID: P17302)". Cardiac Organellar Protein Atlas Knowledgebase (COPaKB). Archived from the original on 5 October 2015. Retrieved 18 September 2015.
- Laird DW (Apr 2014). "Syndromic and non-syndromic disease-linked Cx43 mutations". FEBS Letters. 588 (8): 1339–1348. doi:10.1016/j.febslet.2013.12.022. PMID 24434540.
- Kameritsch P, Pogoda K, Pohl U (Aug 2012). "Channel-independent influence of connexin 43 on cell migration". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1818 (8): 1993–2001. doi:10.1016/j.bbamem.2011.11.016. PMID 22155212.
- Schiavi A, Hudder A, Werner R (Dec 1999). "Connexin43 mRNA contains a functional internal ribosome entry site". FEBS Letters. 464 (3): 118–122. doi:10.1016/S0014-5793(99)01699-3. PMID 10618489.
- Ghosh S, Kumar A, Chandna S (Jul 2015). "Connexin-43 downregulation in G2/M phase enriched tumour cells causes extensive low-dose hyper-radiosensitivity (HRS) associated with mitochondrial apoptotic events". Cancer Letters. 363 (1): 46–59. doi:10.1016/j.canlet.2015.03.046. PMID 25843295.
- Pizzuti A, Flex E, Mingarelli R, Salpietro C, Zelante L, Dallapiccola B (Mar 2004). "A homozygous GJA1 gene mutation causes a Hallermann-Streiff/ODDD spectrum phenotype". Human Mutation. 23 (3): 286. doi:10.1002/humu.9220. PMID 14974090.
- Delmar M, Makita N (May 2012). "Cardiac connexins, mutations and arrhythmias". Current Opinion in Cardiology. 27 (3): 236–241. doi:10.1097/HCO.0b013e328352220e. PMID 22382502.
- Cameron SJ, Malik S, Akaike M, Lerner-Marmarosh N, Yan C, Lee JD, Abe J, Yang J (May 2003). "Regulation of epidermal growth factor-induced connexin 43 gap junction communication by big mitogen-activated protein kinase1/ERK5 but not ERK1/2 kinase activation". The Journal of Biological Chemistry. 278 (20): 18682–18688. doi:10.1074/jbc.M213283200. PMID 12637502.
- Schubert AL, Schubert W, Spray DC, Lisanti MP (May 2002). "Connexin family members target to lipid raft domains and interact with caveolin-1". Biochemistry. 41 (18): 5754–5764. doi:10.1021/bi0121656. PMID 11980479.
- Giepmans BN, Moolenaar WH (1998). "The gap junction protein connexin43 interacts with the second PDZ domain of the zona occludens-1 protein". Current Biology. 8 (16): 931–934. doi:10.1016/S0960-9822(07)00375-2. PMID 9707407.
- Cooper CD, Lampe PD (Nov 2002). "Casein kinase 1 regulates connexin-43 gap junction assembly". The Journal of Biological Chemistry. 277 (47): 44962–44968. doi:10.1074/jbc.M209427200. PMID 12270943.
- Giepmans BN, Feiken E, Gebbink MF, Moolenaar WH (2003). "Association of connexin43 with a receptor protein tyrosine phosphatase". Cell Communication & Adhesion. 10 (4–6): 201–205. doi:10.1080/cac.10.4-6.201.205. PMID 14681016.
Further reading
- Andrew L Harris and Darren Locke (2009). Connexins, A Guide. New York: Springer. p. 574. ISBN 978-1-934115-46-6.
- Saffitz JE, Laing JG, Yamada KA (Apr 2000). "Connexin expression and turnover : implications for cardiac excitability". Circulation Research. 86 (7): 723–728. doi:10.1161/01.res.86.7.723. PMID 10764404.
External links
PDB gallery | |
|---|---|
|