Rhodopsin
Rhodopsin (also known as visual purple) is a light-sensitive receptor protein involved in visual phototransduction. It is named after ancient Greek ῥόδον (rhódon) for rose, due to its pinkish color, and ὄψις (ópsis) for sight.[5] Rhodopsin is a biological pigment found in the rods of the retina and is a G-protein-coupled receptor (GPCR). It belongs to opsins. Rhodopsin is extremely sensitive to light, and thus enables vision in low-light conditions.[6] When rhodopsin is exposed to light, it immediately photobleaches. In humans, it is regenerated fully in about 30 minutes, after which rods are more sensitive.[7]

Rhodopsin was discovered by Franz Christian Boll in 1876.[8][9]
Structure
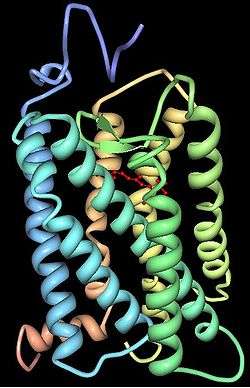
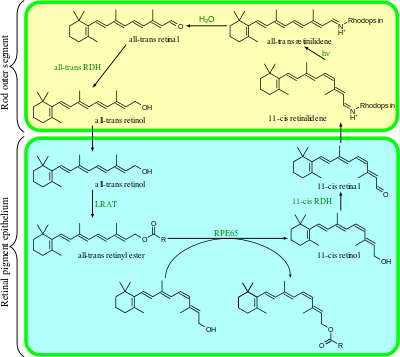
Rhodopsin consists of two components, a protein molecule also called scotopsin and a covalently-bound cofactor called retinal. Scotopsin is an opsin, a light-sensitive G protein coupled receptor that embeds in the lipid bilayer of cell membranes using seven protein transmembrane domains. These domains form a pocket where the photoreactive chromophore, retinal, lies horizontally to the cell membrane, linked to a lysine residue in the seventh transmembrane domain of the protein. Thousands of rhodopsin molecules are found in each outer segment disc of the host rod cell. Retinal is produced in the retina from vitamin A, from dietary beta-carotene. Isomerization of 11-cis-retinal into all-trans-retinal by light sets off a series of conformational changes ('bleaching') in the opsin, eventually leading it to a form called metarhodopsin II (Meta II), which activates an associated G protein, transducin, to trigger a cyclic guanosine monophosphate (cGMP) second messenger cascade.[7][10][11]
Rhodopsin of the rods most strongly absorbs green-blue light and, therefore, appears reddish-purple, which is why it is also called "visual purple".[12] It is responsible for monochromatic vision in the dark.[7]
Several closely related opsins differ only in a few amino acids and in the wavelengths of light that they absorb most strongly. Humans have eight other opsins besides rhodopsin, as well as cryptochrome (light-sensitive, but not an opsin).[13][14]
The photopsins are found in the cone cells of the retina and are the basis of color vision. They have absorption maxima for yellowish-green (photopsin I), green (photopsin II), and bluish-violet (photopsin III) light. The remaining opsin, melanopsin, is found in photosensitive ganglion cells and absorbs blue light most strongly.
In rhodopsin, the aldehyde group of retinal is covalently linked to the amino group of a lysine residue on the protein in a protonated Schiff base (-NH+=CH-).[15] When rhodopsin absorbs light, its retinal cofactor isomerizes from the 11-cis to the all-trans configuration, and the protein subsequently undergoes a series of relaxations to accommodate the altered shape of the isomerized cofactor. The intermediates formed during this process were first investigated in the laboratory of George Wald, who received the Nobel prize for this research in 1967.[16] The photoisomerization dynamics has been subsequently investigated with time-resolved IR spectroscopy and UV/Vis spectroscopy. A first photoproduct called photorhodopsin forms within 200 femtoseconds after irradiation, followed within picoseconds by a second one called bathorhodopsin with distorted all-trans bonds. This intermediate can be trapped and studied at cryogenic temperatures, and was initially referred to as prelumirhodopsin.[17] In subsequent intermediates lumirhodopsin and metarhodopsin I, the Schiff's base linkage to all-trans retinal remains protonated, and the protein retains its reddish color. The critical change that initiates the neuronal excitation involves the conversion of metarhodopsin I to metarhodopsin II, which is associated with deprotonation of the Schiff's base and change in color from red to yellow.[18]
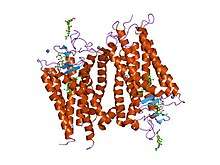
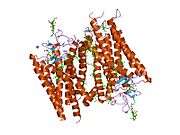
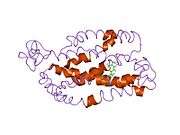
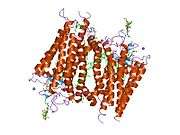
The structure of rhodopsin has been studied in detail via x-ray crystallography on rhodopsin crystals.[19] Several models (e.g., the bicycle-pedal mechanism, hula-twist mechanism) attempt to explain how the retinal group can change its conformation without clashing with the enveloping rhodopsin protein pocket.[20][21][22] Recent data support that rhodopsin is a functional monomer, instead of a dimer, which was the paradigm of G-protein-coupled receptors for many years.[23]
Phototransduction
Rhodopsin is an essential G-protein coupled receptor in phototransduction.
Function
The product of light activation, Metarhodopsin II, initiates the visual phototransduction pathway by stimulating the G protein transducin (Gt), resulting in the liberation of its α subunit. This GTP-bound subunit in turn activates cGMP phosphodiesterase. cGMP phosphodiesterase hydrolyzes (breaks down) cGMP, lowering its local concentration so it can no longer activate cGMP-dependent cation channels. This leads to the hyperpolarization of photoreceptor cells, changing the rate at which they release transmitters.
Deactivation
Meta II (metarhodopsin II) is deactivated rapidly after activating transducin by rhodopsin kinase and arrestin.[24] Rhodopsin pigment must be regenerated for further phototransduction to occur. This means replacing all-trans-retinal with 11-cis-retinal and the decay of Meta II is crucial in this process. During the decay of Meta II, the Schiff base link that normally holds all-trans-retinal and the apoprotein opsin (aporhodopsin) is hydrolyzed and becomes Meta III. In the rod outer segment, Meta III decays into separate all-trans-retinal and opsin.[24] A second product of Meta II decay is an all-trans-retinal opsin complex in which the all-trans-retinal has been translocated to second binding sites. Whether the Meta II decay runs into Meta III or the all-trans-retinal opsin complex seems to depend on the pH of the reaction. Higher pH tends to drive the decay reaction towards Meta III.[24]
Retinal disease
Mutation of the rhodopsin gene is a major contributor to various retinopathies such as retinitis pigmentosa. In general, the disease-causing protein aggregates with ubiquitin in inclusion bodies, disrupts the intermediate filament network, and impairs the ability of the cell to degrade non-functioning proteins, which leads to photoreceptor apoptosis.[25] Other mutations on rhodopsin lead to X-linked congenital stationary night blindness, mainly due to constitutive activation, when the mutations occur around the chromophore binding pocket of rhodopsin.[26] Several other pathological states relating to rhodopsin have been discovered including poor post-Golgi trafficking, dysregulative activation, rod outer segment instability and arrestin binding.[26]
Microbial rhodopsins
Some prokaryotes express proton pumps called bacteriorhodopsins, archaerhodopsins, proteorhodopsins, and xanthorhodopsins to carry out phototrophy.[27] Like animal visual pigments, these contain a retinal chromophore (although it is an all-trans, rather than 11-cis form) and have seven transmembrane alpha helices; however, they are not coupled to a G protein. Prokaryotic halorhodopsins are light-activated chloride pumps.[27] Unicellular flagellate algae contain channelrhodopsins that act as light-gated cation channels when expressed in heterologous systems. Many other pro- and eukaryotic organisms (in particular, fungi such as Neurospora) express rhodopsin ion pumps or sensory rhodopsins of yet-unknown function. Very recently, microbial rhodopsins with guanylyl cyclase activity have been discovered.[28][29][30] While all microbial rhodopsins have significant sequence homology to one another, they have no detectable sequence homology to the G-protein-coupled receptor (GPCR) family to which animal visual rhodopsins belong. Nevertheless, microbial rhodopsins and GPCRs are possibly evolutionarily related, based on the similarity of their three-dimensional structures. Therefore, they have been assigned to the same superfamily in Structural Classification of Proteins (SCOP).[31]
References
- GRCh38: Ensembl release 89: ENSG00000163914 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000030324 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Perception (2008), Guest Editorial Essay, Perception, p. 1
- Litmann BJ, Mitchell DC (1996). "Rhodopsin structure and function". In Lee AG (ed.). Rhodopsin and G-Protein Linked Receptors, Part A (Vol 2, 1996) (2 Vol Set). Greenwich, Conn: JAI Press. pp. 1–32. ISBN 978-1-55938-659-3.
- Stuart JA, Brige RR (1996). "Characterization of the primary photochemical events in bacteriorhodopsin and rhodopsin". In Lee AG (ed.). Rhodopsin and G-Protein Linked Receptors, Part A (Vol 2, 1996) (2 Vol Set). Greenwich, Conn: JAI Press. pp. 33–140. ISBN 978-1-55938-659-3.
- Encyclopedia of the Neurological Sciences. Academic Press. 29 April 2014. pp. 441–. ISBN 978-0-12-385158-1.
- Giese AC (24 September 2013). Photophysiology: General Principles; Action of Light on Plants. Elsevier. p. 9. ISBN 978-1-4832-6227-7. Retrieved 23 September 2015.
- Hofmann KP, Heck M (1996). "Light-induced protein-protein interactions on the rod photoreceptor disc membrane". In Lee AG (ed.). Rhodopsin and G-Protein Linked Receptors, Part A (Vol 2, 1996) (2 Vol Set). Greenwich, Conn: JAI Press. pp. 141–198. ISBN 978-1-55938-659-3.
- Kolb H, Fernandez E, Nelson R, Jones BW (1 March 2010). "Webvision: Photoreceptors". University of Utah. Archived from the original on 16 August 2000.
- Rogers K. "Rhodopsin". Encyclopædia Britannica. Britannica.com. Retrieved 30 January 2016.
- Terakita A (2005). "The opsins". Genome Biology. 6 (3): 213. doi:10.1186/gb-2005-6-3-213. PMC 1088937. PMID 15774036.
- Foley LE, Gegear RJ, Reppert SM (June 2011). "Human cryptochrome exhibits light-dependent magnetosensitivity". Nature Communications. 2: 356. Bibcode:2011NatCo...2..356F. doi:10.1038/ncomms1364. PMC 3128388. PMID 21694704.
- Bownds D, Wald G (January 1965). "Reaction of the rhodopsin chromophore with sodium borohydride". Nature. 205 (4968): 254–7. Bibcode:1965Natur.205..254B. doi:10.1038/205254a0. PMID 14270706.
- The Nobel Foundation. "The Nobel Prize in Physiology or Medicine 1967". Nobelprize.org. Nobel Media AB 2014. Retrieved 12 December 2015.
- Yoshizawa T, Wald G (March 1963). "Pre-lumirhodopsin and the bleaching of visual pigments". Nature. 197 (Mar 30): 1279–86. Bibcode:1963Natur.197.1279Y. doi:10.1038/1971279a0. PMID 14002749.
- Matthews RG, Hubbard R, Brown PK, Wald G (November 1963). "Tautomeric forms of metarhodopsin". The Journal of General Physiology. 47 (2): 215–40. doi:10.1085/jgp.47.2.215. PMC 2195338. PMID 14080814.
- Gulati S, Jastrzebska B, Banerjee S, Placeres ÁL, Miszta P, Gao S, Gunderson K, Tochtrop GP, Filipek S, Katayama K, Kiser PD, Mogi M, Stewart PL, Palczewski K (March 2017). "Photocyclic behavior of rhodopsin induced by an atypical isomerization mechanism". Proceedings of the National Academy of Sciences. 114 (13): E2608-15. doi:10.1073/pnas.1617446114. PMC 5380078. PMID 28289214.
- Nakamichi H, Okada T (June 2006). "Crystallographic analysis of primary visual photochemistry". Angewandte Chemie. 45 (26): 4270–3. doi:10.1002/anie.200600595. PMID 16586416.
- Schreiber M, Sugihara M, Okada T, Buss V (June 2006). "Quantum mechanical studies on the crystallographic model of bathorhodopsin". Angewandte Chemie. 45 (26): 4274–7. doi:10.1002/anie.200600585. PMID 16729349.
- Weingart O (September 2007). "The twisted C11=C12 bond of the rhodopsin chromophore--a photochemical hot spot". Journal of the American Chemical Society. 129 (35): 10618–9. doi:10.1021/ja071793t. PMID 17691730.
- Chabre M, le Maire M (July 2005). "Monomeric G-protein-coupled receptor as a functional unit". Biochemistry. 44 (27): 9395–403. doi:10.1021/bi050720o. PMID 15996094.
- Heck M, Schädel SA, Maretzki D, Bartl FJ, Ritter E, Palczewski K, Hofmann KP (January 2003). "Signaling states of rhodopsin. Formation of the storage form, metarhodopsin III, from active metarhodopsin II". The Journal of Biological Chemistry. 278 (5): 3162–9. doi:10.1074/jbc.M209675200. PMC 1364529. PMID 12427735.
- Saliba RS, Munro PM, Luthert PJ, Cheetham ME (July 2002). "The cellular fate of mutant rhodopsin: quality control, degradation and aggresome formation". Journal of Cell Science. 115 (Pt 14): 2907–18. PMID 12082151.
- Mendes HF, van der Spuy J, Chapple JP, Cheetham ME (April 2005). "Mechanisms of cell death in rhodopsin retinitis pigmentosa: implications for therapy". Trends in Molecular Medicine. 11 (4): 177–85. doi:10.1016/j.molmed.2005.02.007. PMID 15823756.
- Bryant DA, Frigaard NU (November 2006). "Prokaryotic photosynthesis and phototrophy illuminated". Trends in Microbiology. 14 (11): 488–96. doi:10.1016/j.tim.2006.09.001. PMID 16997562.
- Gao SQ, Nagpal J, Schneider MW, Kozjak-Pavlovic V, Nagel G, Gottschalk A (July 2015). "Optogenetic manipulation of cGMP in cells and animals by the tightly light-regulated guanylyl-cyclase opsin CyclOp". Nature Communications. 6 (8046): 8046. Bibcode:2015NatCo...6.8046G. doi:10.1038/ncomms9046. PMC 4569695. PMID 26345128.
- Scheib U, Stehfest K, Gee CE, Körschen HG, Fudim R, Oertner TG, Hegemann P (August 2015). "The rhodopsin-guanylyl cyclase of the aquatic fungus Blastocladiella emersonii enables fast optical control of cGMP signaling". Science Signaling. 8 (389): rs8. doi:10.1126/scisignal.aab0611. PMID 26268609.
- Scheib U, Broser M, Constantin OM, Yang S, Gao S, Mukherjee S, et al. (May 2018). "Rhodopsin-cyclases for photocontrol of cGMP/cAMP and 2.3 Å structure of the adenylyl cyclase domain". Nature Communications. 9 (1): 2046. Bibcode:2018NatCo...9.2046S. doi:10.1038/s41467-018-04428-w. PMC 5967339. PMID 29799525.
- "Superfamily: Bacterial photosystem II reaction centre, L and M subunits". SCOP.
Further reading
- See also bacteriorhodopsin, used in some halobacteria as a light-driven proton pump.
- Humphries P, Kenna P, Farrar GJ (May 1992). "On the molecular genetics of retinitis pigmentosa". Science. 256 (5058): 804–8. Bibcode:1992Sci...256..804H. doi:10.1126/science.1589761. PMID 1589761.
- Edwards SC (July 1995). "Involvement of cGMP and calcium in the photoresponse in vertebrate photoreceptor cells". The Journal of the Florida Medical Association. 82 (7): 485–8. PMID 7673885.
- al-Maghtheh M, Gregory C, Inglehearn C, Hardcastle A, Bhattacharya S (1993). "Rhodopsin mutations in autosomal dominant retinitis pigmentosa". Human Mutation. 2 (4): 249–55. doi:10.1002/humu.1380020403. PMID 8401533.
- Garriga P, Manyosa J (September 2002). "The eye photoreceptor protein rhodopsin. Structural implications for retinal disease". FEBS Letters. 528 (1–3): 17–22. doi:10.1016/S0014-5793(02)03241-6. PMID 12297272.
- Inglehearn CF, Keen TJ, Bashir R, Jay M, Fitzke F, Bird AC, Crombie A, Bhattacharya S (April 1992). "A completed screen for mutations of the rhodopsin gene in a panel of patients with autosomal dominant retinitis pigmentosa". Human Molecular Genetics. 1 (1): 41–5. doi:10.1093/hmg/1.1.41. PMID 1301135.
- Farrar GJ, Findlay JB, Kumar-Singh R, Kenna P, Humphries MM, Sharpe E, Humphries P (December 1992). "Autosomal dominant retinitis pigmentosa: a novel mutation in the rhodopsin gene in the original 3q linked family". Human Molecular Genetics. 1 (9): 769–71. doi:10.1093/hmg/1.9.769. PMID 1302614.
- Robinson PR, Cohen GB, Zhukovsky EA, Oprian DD (October 1992). "Constitutively active mutants of rhodopsin". Neuron. 9 (4): 719–25. doi:10.1016/0896-6273(92)90034-B. PMID 1356370.
- Fujiki K, Hotta Y, Hayakawa M, Sakuma H, Shiono T, Noro M, Sakuma T, Tamai M, Hikiji K, Kawaguchi R (June 1992). "Point mutations of rhodopsin gene found in Japanese families with autosomal dominant retinitis pigmentosa (ADRP)". The Japanese Journal of Human Genetics. 37 (2): 125–32. doi:10.1007/BF01899733. PMID 1391967.
- Olsson JE, Gordon JW, Pawlyk BS, Roof D, Hayes A, Molday RS, Mukai S, Cowley GS, Berson EL, Dryja TP (November 1992). "Transgenic mice with a rhodopsin mutation (Pro23His): a mouse model of autosomal dominant retinitis pigmentosa". Neuron. 9 (5): 815–30. doi:10.1016/0896-6273(92)90236-7. PMID 1418997.
- Andréasson S, Ehinger B, Abrahamson M, Fex G (September 1992). "A six-generation family with autosomal dominant retinitis pigmentosa and a rhodopsin gene mutation (arginine-135-leucine)". Ophthalmic Paediatrics and Genetics. 13 (3): 145–53. doi:10.3109/13816819209046483. PMID 1484692.
- Inglehearn CF, Lester DH, Bashir R, Atif U, Keen TJ, Sertedaki A, Lindsey J, Jay M, Bird AC, Farrar GJ (March 1992). "Recombination between rhodopsin and locus D3S47 (C17) in rhodopsin retinitis pigmentosa families". American Journal of Human Genetics. 50 (3): 590–7. PMC 1684283. PMID 1539595.
- Fishman GA, Stone EM, Gilbert LD, Sheffield VC (May 1992). "Ocular findings associated with a rhodopsin gene codon 106 mutation. Glycine-to-arginine change in autosomal dominant retinitis pigmentosa". Archives of Ophthalmology. 110 (5): 646–53. doi:10.1001/archopht.1992.01080170068026. PMID 1580841.
- Keen TJ, Inglehearn CF, Lester DH, Bashir R, Jay M, Bird AC, Jay B, Bhattacharya SS (September 1991). "Autosomal dominant retinitis pigmentosa: four new mutations in rhodopsin, one of them in the retinal attachment site". Genomics. 11 (1): 199–205. doi:10.1016/0888-7543(91)90119-Y. PMID 1765377.
- Dryja TP, Hahn LB, Cowley GS, McGee TL, Berson EL (October 1991). "Mutation spectrum of the rhodopsin gene among patients with autosomal dominant retinitis pigmentosa". Proceedings of the National Academy of Sciences of the United States of America. 88 (20): 9370–4. Bibcode:1991PNAS...88.9370D. doi:10.1073/pnas.88.20.9370. PMC 52716. PMID 1833777.
- Gal A, Artlich A, Ludwig M, Niemeyer G, Olek K, Schwinger E, Schinzel A (October 1991). "Pro-347-Arg mutation of the rhodopsin gene in autosomal dominant retinitis pigmentosa". Genomics. 11 (2): 468–70. doi:10.1016/0888-7543(91)90159-C. PMID 1840561.
- Sung CH, Davenport CM, Hennessey JC, Maumenee IH, Jacobson SG, Heckenlively JR, Nowakowski R, Fishman G, Gouras P, Nathans J (August 1991). "Rhodopsin mutations in autosomal dominant retinitis pigmentosa". Proceedings of the National Academy of Sciences of the United States of America. 88 (15): 6481–5. Bibcode:1991PNAS...88.6481S. doi:10.1073/pnas.88.15.6481. PMC 52109. PMID 1862076.
- Jacobson SG, Kemp CM, Sung CH, Nathans J (September 1991). "Retinal function and rhodopsin levels in autosomal dominant retinitis pigmentosa with rhodopsin mutations". American Journal of Ophthalmology. 112 (3): 256–71. doi:10.1016/s0002-9394(14)76726-1. PMID 1882937.
- Sheffield VC, Fishman GA, Beck JS, Kimura AE, Stone EM (October 1991). "Identification of novel rhodopsin mutations associated with retinitis pigmentosa by GC-clamped denaturing gradient gel electrophoresis". American Journal of Human Genetics. 49 (4): 699–706. PMC 1683182. PMID 1897520.
External links
| Wikimedia Commons has media related to Rhodopsins. |
- Rhodopsin at the US National Library of Medicine Medical Subject Headings (MeSH)
- Kolb H, Fernandez E, Nelson R, Jones BW (1 March 2010). "Webvision Home Page: The organization of the retina and visual system". University of Utah.
- The Rhodopsin Protein
- Photoisomerization of rhodopsin, animation.
- Rhodopsin and the eye, summary with pictures.
PDB gallery | |
|---|---|
|