Opsin
Opsins are a group of proteins, made light-sensitive, via the chromophore retinal (or a variant) found in photoreceptor cells of the retina. Five classical groups of opsins are involved in vision, mediating the conversion of a photon of light into an electrochemical signal, the first step in the visual transduction cascade. Another opsin found in the mammalian retina, melanopsin, is involved in circadian rhythms and pupillary reflex but not in vision.
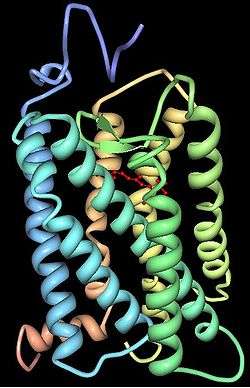
Opsin classification
Opsins can be classified several ways, including function (vision, phototaxis, photoperiodism, etc.), type of chromophore (retinal, flavine, bilin), molecular structure (tertiary, quaternary), signal output (phosphorylation, reduction, oxidation), etc.[1]
There are two groups of protein termed opsins.[2][3] Type I opsins are employed by prokaryotes and by some algae (as a component of channelrhodopsins) and fungi,[4] whereas animals use type II opsins. No opsins have been found outside these groups (for instance in plants, or placozoans).[2]
At one time it was thought that type I and type II were related because of structural and functional similarities. With the advent of genetic sequencing it became apparent that sequence identity was no greater than could be accounted for by random chance. However, in recent years new methods have been developed specific to deep phylogeny. As a result, several studies have found evidence of a possible phylogenetic relationship between the two.[5][6][7] However, this does not necessarily mean that the last common ancestor of type I and II opsins was itself an opsin, a light sensitive receptor: all animal opsins arose (by gene duplication and divergence) late in the history of the large G-protein coupled receptor (GPCR) gene family, which itself arose after the divergence of plants, fungi, choanflagellates and sponges from the earliest animals. The retinal chromophore is found solely in the opsin branch of this large gene family, meaning its occurrence elsewhere represents convergent evolution, not homology. Microbial rhodopsins are, by sequence, very different from any of the GPCR families.[8] According to one hypothesis, both type-I and type-II opsins belong to the transporter-opsin-G protein-coupled receptor (TOG) superfamily, a proposed clade that includes G protein-coupled receptor (GPCR), Ion-translocating microbial rhodopsin (MR), and seven others.[9]
Type I opsins
Type I opsins (also known as microbial opsins) are seven-transmembrane-domain proteins. Most of them are ion channels or pumps instead of proper receptors and do not bind to a G protein. Type I opsins are found in all three domains of life: Archaea, Bacteria, and Eukaryota. In Eukaryota, type I opsins are found mainly in unicellular organisms such as green algae, and in fungi. In most complex multicellular eukaryotes, type I opsins have been replaced with other light-sensitive molecules such as cryptochrome and phytochrome in plants, and type II opsins in Metazoa (animals).[10]
Microbial opsins are often known by the rhodopsin form of the molecule, i.e., rhodopsin (in the broad sense) = opsin + chromophore. Among the many kinds of microbial opsins are the proton pumps bacteriorhodopsin (BR) and xanthorhodopsin (xR), the chloride pump halorhodopsin (HR) the photosensors sensory rhodopsin I (SRI) and sensory rhodopsin II (SRII), as well as proteorhodopsin (PR), Neurospora opsin I (NOPI), Chlamydomonas sensory rhodopsins A (CSRA), Chlamydomonas sensory rhodopsins B (CSRB), channelrhodopsin (ChR), and archaerhodopsin (Arch).[11]
Several type I opsins, such as proteo- and bacteriorhodopsin, are used by various bacterial groups to harvest energy from light to carry out metabolic processes using a non-chlorophyll-based pathway. Beside that, halorhodopsins of Halobacteria and channelrhodopsins of some algae, e.g. Volvox, serve them as light-gated ion channels, amongst others also for phototactic purposes. Sensory rhodopsins exist in Halobacteria that induce a phototactic response by interacting with transducer membrane-embedded proteins that have no relation to G proteins.[12]
Type I opsins (like channelrhodopsin, halorhodopsin, and archaerhodopsin) are used in optogenetics to switch on or off neuronal activity. Type I opsins are preferred if the neuronal activity should be modulated at higher frequency, because they respond faster than type II opsins. This is because type I opsins are ion channels or proton/ion pumps and thus are activated by light directly, while type II opsins activate G-proteins, which then activate effector enzymes that produce metabolites to open ion channels.[13]
Type II opsins
Type II opsins (or animal opsins) are members of the seven-transmembrane-domain proteins (35–55 kDa) of the G protein-coupled receptor (GPCR) superfamily.[14]
Type II opsins fall phylogenetically into four groups: C-opsins (Ciliary), Cnidops (cnidarian opsins), R-opsins (rhabdomeric), and Go/RGR opsins (also known as RGR/Go or Group 4 opsins). The Go/RGR opsins are divided into four sub-clades: Go-opsins, RGR, Peropsins, and Neuropsins. C-opsins, R-opsins, and the Go/RGR opsins are found only in Bilateria.[15] [16]
Type II visual opsins are traditionally classified as either ciliary or rhabdomeric. Ciliary opsins, found in vertebrates and cnidarians, attach to ciliary structures such as rods and cones. Rhabdomeric opsins are attached to light-gathering organelles called rhabdomeres. This classification cuts across phylogenetic categories (clades) so that both the terms "ciliary" and "rhabdomeric" can be ambiguous. Here, "C-opsins (ciliary)" refers to a clade found exclusively in Bilateria and excludes cnidarian ciliary opsins such as those found in the box jellyfish. Similarly, "R-opsin (rhabdomeric)" includes melanopsin even though it does not occur on rhabdomeres in vertebrates.[15]
C-opsins (ciliary)
Ciliary opsins (or c-opsins) are expressed in ciliary photoreceptor cells, and include the vertebrate visual opsins and encephalopsins.[17] They convert light signals to nerve impulses via cyclic nucleotide gated ion channels, which work by increasing the charge differential across the cell membrane (i.e. hyperpolarization.[2])
Vertebrate visual opsins
Vertebrate visual opsins are a subset of C-opsins (ciliary). They are expressed in the vertebrate retina and mediate vision. They can be further subdivided into rod opsins and four types of cone opsin.[17] Rod opsins (rhodopsins, usually denoted Rh),[18] are used in dim-light vision, are thermally stable, and are found in the rod photoreceptor cells. Cone opsins, employed in color vision, are less-stable opsins located in the cone photoreceptor cells. Cone opsins are further subdivided according to their absorption maxima (λmax), the wavelength at which the highest light absorption is observed. Evolutionary relationships, deduced using the amino acid sequence of the opsins, are also frequently used to categorize cone opsins into their respective group. Both methods predict four general cone opsin groups in addition to rhodopsin.[19]
| ||||||||||||||||||||||||||||
Vertebrates typically have four cone opsins (LWS, SWS1, SWS2, and Rh2) inherited from the first vertebrate (and thus predating the first vertebrate), as well as the rod opsin, rhodopsin (Rh1), which emerged after the first vertebrate but before the first Gnathostome (jawed vertebrate). These five opsins emerged through a series of gene duplications beginning with LWS and ending with Rh1. Each one has since evolved into numerous variants and thus constitutes an opsin family or subtype.[20][21]
| Name | Abbr. | Photo receptor |
λmax | Color | Human variant |
|---|---|---|---|---|---|
| Long-wave sensitive | LWS | Cone | 500–570 nm | Green, yellow, red | OPN1LW "red" / OPN1MW "green" |
| Short-wave sensitive 1 | SWS1 | Cone | 355–445 nm | Ultraviolet, violet | OPN1SW "blue" (extinct in monotremes) |
| Short-wave sensitive 2 | SWS2 | Cone | 400–470 nm | Violet, blue | (Extinct in therian mammals) |
| Rhodopsin-like 2 | Rh2 | Cone | 480–530 nm | Green | (Extinct in mammals) |
| Rhodopsin-like 1 (vertebrate rhodopsin) |
Rh1 | Rod | ~500 nm | Blue–green | OPN2/Rho, human rhodopsin |
Humans have the following set of photoreceptor proteins responsible for vision:
- Rhodopsin (Rh1, OPN2, RHO) – expressed in rod cells, used in night vision
- Three cone opsins (also known as photopsins) – expressed in cone cells, used in color vision
- Long-wavelength sensitive (OPN1LW) Opsin – λmax of 560 nm, in the yellow-green region of the electromagnetic spectrum.[22] May be called the "red opsin," "erythrolabe," "L opsin" or "LWS opsin." Note that despite its common name as the "red" opsin, this opsin's peak sensitivity is not in the red region of the spectrum. However, it is more sensitive to red than the other two human opsins.[22] This receptor also has a secondary response in the violet high frequencies.[23][24]
- Middle-wavelength sensitive (OPN1MW) Opsin – λmax of 530 nm, in the green region of the electromagnetic spectrum.[22] May be called the "green opsin," "chlorolabe," "M opsin" or "MWS opsin."
- Short-wavelength sensitive (OPN1SW) Opsin – λmax of 430 nm, in the blue region of the electromagnetic spectrum.[22] May be called the "blue opsin," "cyanolabe," "S opsin" or "SWS opsin."
Pinopsins
The first Pineal Opsin (Pinopsin) was found in the chicken pineal gland. It is a blue sensitive opsin (λmax = 470 nm).[25]
wide range of expression in the brain, most notably in the pineal region
Vertebrate Ancient (VA) opsin
Vertebrate Ancient (VA) opsin has three isoforms VA short (VAS), VA medium (VAM), and VA long (VAL). It is expressed in the inner retina, within the horizontal and amacrine cells, as well as the pineal organ and habenular region of the brain.[26] It is sensitive to approximately 500 nm [14], found in most vertebrate classes, but not in mammals.[27]
Parapinopsins
The first parapinopsin (PP) opsin was found in the parapineal organ of the catfish.[28] The parapinopsin of lamprey is a UV-sensitive opsin (λmax = 370 nm).[29] The teleosts have two groups of parapinopsins, one is sensitive to UV (λmax = 360-370 nm), the other is sensitive to blue (λmax = 460-480 nm) light.[30]
Parietopsins
The first parietopsin was found in the photoreceptor cells of the lizard parietal eye. The lizard parietopsin is green-sensitive (λmax = 522 nm), and despite it is a c-opsin, like the vertebrate visual opsins, it does not induce hyperpolarization via a Gt-protein, but induces depolarization via a Go-protein.[31][32]
OPN3 (Encephalopsin or Panopsin)
Panopsins are found in many tissues (skin,[33] brain,[34][35] testes,[34] heart, liver,[35] kidney, skeletal muscle, lung, pancreas and retina[35]). They were originally found in the human and mouse brain and thus called encephalopsin.[34]
The first invertebrate panopsin was found in the ciliary photoreceptor cells of the annelid Platynereis dumerilii and is called c(iliary)-opsin.[36] This c-opsin is UV-sensitive (λmax = 383 nm) and can be tuned by 125 nm at a single amino-acid (range λmax = 377 - 502 nm).[37] Thus, not unsurprisingly, a second but cyan sensitive c-opsin (λmax = 490 nm) exists in Platynereis dumerilii.[38] The first c-opsin mediates in the larva UV induced gravitaxis. The gravitaxis forms with phototaxis a ratio-chromatic depth-gauge.[39] In different depths, the light in water is composed of different wavelengths: First the red (> 600 nm) and the UV and violet (< 420 nm) wavelengths disappear. The higher the depth the narrower the spectrum so that only cyan light (480 nm) is left.[40] Thus, the larvae can determine their depth by color. The color unlike brightness stays almost constant independent of time of day or the weather, for instance if it is cloudy.[41][42]
Panopsins are also expressed in the brains of some insects.[17] The panopsins of mosquito and pufferfish absorb maximally at 500 nm and 460 nm, respectively. Both activate in vitro Gi and Go proteins.[43]
The panopsins of teleost fish are called: Teleost multiple tissue (TMT) opsins.
Teleost Multiple Tissue (TMT) Opsin
Teleost fish opsins are expressed in many tissues and therefore called Teleost Multiple Tissue (TMT) opsins.[44] TMT opsins form three groups which are most closely related to a fourth groups the panopsins.[45][46] In fact, TMT opsins in teleost fish are orthologous to the panopsins in the other vertebrates. They also have the same introns and the same place, which confirms that they belong together.[44]
Cnidarian opsins
Cnidaria, which include jellyfish, corals, and sea anemones, are the most basal animals to possess complex eyes. Jellyfish opsins in the rhopalia couple to Gs-proteins raising the intracellular cAMP level.[47][48] Coral opsins can couple to Gq-proteins and Gc-proteins. Gc-proteins are a subtype of G-proteins specific to cnidarians.[49] The cnidarian opsins have been identified as one group and so called cnidops,[15] however at least some of them belong to the c-opsins, r-opsins, and Go/RGR-opsins found in bilaterians.[14][50][51]
r-opsins (rhabdomeric) / Gq-coupled
Rhabdomeric opsins (or r-opsins) are also known as Gq-opsins, because they couple to a Gq-protein. R-opsins are used by molluscs and arthropods. Arthropods appear to attain colour vision in a similar fashion to the vertebrates, by using three (or more) distinct groups of opsins, distinct both in terms of phylogeny and spectral sensitivity.[17] The r-opsin melanopsin is also expressed in vertebrates, where it regulates circadian rhythms and mediates the pupillary reflex.[17]
Unlike c-opsins, r-opsins are associated with canonical transient receptor potential ion channels; these lead to the electric potential difference across a cell membrane being eradicated (i.e. depolarization).[2]
The identification of the crystal structure of squid rhodopsin[52] is likely to further our understanding of its function in this group.
Arthropods use different opsins in their different eye types, but at least in Limulus the opsins expressed in the lateral and the compound eyes are 99% identical and presumably diverged recently.[53]
Melanopsin OPN4
Involved in circadian rhythms, pupillary reflex, and color correction in high-brightness situations. Phylogenetically a member of the R-opsin (rhabdomeric) group, functionally and structurally an r-opsin, but does not occur in rhabdomeres.
Go/RGR (Group 4) opsins
Go/RGR opsins include Go-opsins, RGR-opsins, neuropsins, and peropsins.
Go-opsins
Go-opsins are absent from higher vertebrates[15] and ecdysozoans.[54] They are found in the ciliary photoreceptor cells of the scallop eye[55] and the basal chordate amphioxus.[56] In Platynereis dumerilii however, a Go-opsin is expressed in the rhabdomeric photoreceptor cells of the eyes.[40]
RGR opsins
RGR opsins, also known as Retinal G protein coupled receptors are expressed in the retinal pigment epithelium (RPE) and Müller cells.[57] They preferentially bind all-trans-retinal in the dark instead of 11-cis-retinal.[58] RGR opsins were thought to be photomerases.[19] But instead, they regulate retinoid traffic and production.[17][59] In particular, they speed up light-independently the production of 11-cis-retinol (a precursor of 11-cis-retinal) from all-trans-retinyl-esters.[60] However, the all-trans-retinyl-esters are made available light-dependently by RGR-opsins. Whether RGR-opsins regulate this via a G-protein or another signaling mechanism is unknown.[61] The cattle RGR opsin absorbs maximally at different wavelengths depending on the pH-value. At high pH it absorbs maximally blue (469 nm) light and at low pH it absorbs maximally UV (370 nm) light.[62]
Peropsin
Peropsin, a visual pigment-like receptor, is a protein that in humans is encoded by the RRH gene.[63]
Neuropsins
Neuropsins are sensitive to UVA, typically at 380 nm. They are found in the brain, testes, skin, and retina of humans and rodents, as well as in the brain and retina of birds. In birds and rodents they mediate ultraviolet vision.[33][64][65] They couple to Gi-proteins.[64][65] In humans, Neuropsin is encoded by the OPN5 gene. In the human retina, its function is unknown. In the mouse, it photo-entrains the retina and cornea at least ex vivo.[66]
Unclassified
Extraretinal (or extra-ocular) Rhodopsin-Like Opsins (Exo-Rh)
These pineal opsins, found in the Actinopterygii (ray-finned fish) apparently arose as a result of gene duplication from Rh1 (rhodopsin). These opsins appear to serve functions similar to those of pinopsin found in birds and reptiles.[67] [68]
Structure and function
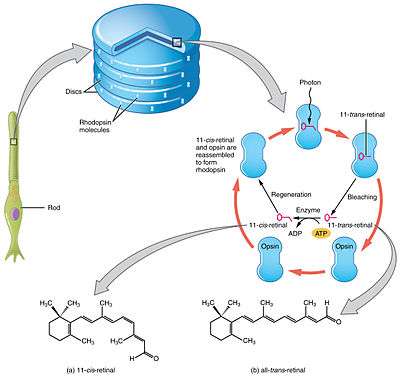
Opsin proteins covalently bind to a vitamin A-based retinaldehyde chromophore through a Schiff base linkage to a lysine residue in the seventh transmembrane alpha helix. In vertebrates, the chromophore is either 11-cis-retinal (A1) or 11-cis-3,4-didehydroretinal (A2) and is found in the retinal binding pocket of the opsin. The absorption of a photon of light results in the photoisomerization of the chromophore from the 11-cis to an all-trans conformation. The photoisomerization induces a conformational change in the opsin protein, causing the activation of the phototransduction cascade. The opsin remains insensitive to light in the trans form. It is regenerated by the replacement of the all-trans retinal by a newly synthesized 11-cis-retinal provided from the retinal epithelial cells. Opsins are functional while bound to either chromophore, with A2-bound opsin λmax being at a longer wavelength than A1-bound opsin.
Opsins contain seven transmembrane α-helical domains connected by three extra-cellular and three cytoplasmic loops. Many amino acid residues, termed functionally conserved residues, are highly conserved between all opsin groups, indicative of important functional roles. All residue positions discussed henceforth are relative to the 348 amino acid bovine rhodopsin crystallized by Palczewski et al.[69] Lys296 is conserved in all known opsins and serves as the site for the Schiff base linkage with the chromophore. Cys138 and Cys110 form a highly conserved disulfide bridge. Glu113 serves as the counterion, stabilizing the protonation of the Schiff linkage between Lys296 and the chromophore. The Glu134-Arg135-Tyr136 is another highly conserved motif, involved in the propagation of the transduction signal once a photon has been absorbed.
Certain amino acid residues, termed spectral tuning sites, have a strong effect on λmax values. Using site-directed mutagenesis, it is possible to selectively mutate these residues and investigate the resulting changes in light absorption properties of the opsin. It is important to differentiate spectral tuning sites, residues that affect the wavelength at which the opsin absorbs light, from functionally conserved sites, residues important for the proper functioning of the opsin. They are not mutually exclusive, but, for practical reasons, it is easier to investigate spectral tuning sites that do not affect opsin functionality. For a comprehensive review of spectral tuning sites see Yokoyama[70] and Deeb.[71] The impact of spectral tuning sites on λmax differs between different opsin groups and between opsin groups of different species.
Opsins in the human eye, brain, and skin
| Abbr. | Name | λmax | Color | Eye | Brain | Skin | Chromosomal location a |
|---|---|---|---|---|---|---|---|
| OPN1LW | L-cone (red-cone) opsin | 557 nm | Yellow | Cone | N/A | N/A | Xq28[19] |
| OPN1MW | M-cone (green-cone) opsin | 527 nm | Green | Cone | N/A | N/A | Xq28[19] |
| OPN1SW | S-cone (blue-cone) opsin | 420 nm | Violet | Cone | N/A | Melanocytes, keratinocytes[33] | 7q32.1[19] |
| OPN2 (RHO) | Rhodopsin | 505 nm | Blue–green | Rod | N/A | Melanocytes, keratinocytes[33] | 3q22.1[19] |
| OPN3 | Encephalopsin, panopsin | S-M | Blue–green | Rod, cone, OPL, IPL, GCL[72] | Cerebral cortex, cerebellum, striatum, thalamus, hypothalamus[34][73] | Melanocytes, keratinocytes[33] | 1q43[19] |
| OPN4 | Melanopsin | 480 nm[74] | Sky blue | ipRGC[74] | N/A | N/A | 10q23.2[19] |
| OPN5 | Neuropsin | 380 nm[64] | Ultraviolet[64] | Neural retina, RPE[75] | Anterior hypothalamus[76] | Melanocytes, keratinocytes[33] | 6p12.3[19] |
| RRH | Peropsin | RPE cells - microvilli | N/A | N/A | 4q25[19] | ||
| RGR | Retinal G protein coupled receptor | RPE cells | N/A | N/A | 10q23.1[19] |
RPE, retinal pigment epithelium; ipRGC, intrinsically photosensitive retinal ganglion cells; OPL, outer plexiform layer; IPL, inner plexiform layer; GCL, ganglion cell layer
See also
External links
- Review of opsins and current research: Shichida Y, Matsuyama T (October 2009). "Evolution of opsins and phototransduction". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 364 (1531): 2881–95. doi:10.1098/rstb.2009.0051. PMC 2781858. PMID 19720651.
- Illustration at Baldwin-Wallace College
- Opsin at the US National Library of Medicine Medical Subject Headings (MeSH)
References
- Björn LO (2 January 2015). Photobiology: The Science of Light and Life. Springer. p. 169. ISBN 978-1-4939-1468-5. Retrieved 3 September 2015.
- Plachetzki DC, Fong CR, Oakley TH (July 2010). "The evolution of phototransduction from an ancestral cyclic nucleotide gated pathway". Proceedings. Biological Sciences. 277 (1690): 1963–9. doi:10.1098/rspb.2009.1797. PMC 2880087. PMID 20219739.
- Fernald RD (September 2006). "Casting a genetic light on the evolution of eyes" (PDF). Science. 313 (5795): 1914–8. Bibcode:2006Sci...313.1914F. doi:10.1126/science.1127889. PMID 17008522.
- Waschuk SA, Bezerra AG, Shi L, Brown LS (May 2005). "Leptosphaeria rhodopsin: bacteriorhodopsin-like proton pump from a eukaryote". Proceedings of the National Academy of Sciences of the United States of America. 102 (19): 6879–83. Bibcode:2005PNAS..102.6879W. doi:10.1073/pnas.0409659102. PMC 1100770. PMID 15860584.
- Shen L, Chen C, Zheng H, Jin L (2013). "The evolutionary relationship between microbial rhodopsins and metazoan rhodopsins". TheScientificWorldJournal. 2013: 435651. doi:10.1155/2013/435651. PMC 3583139. PMID 23476135.
- Devine EL, Oprian DD, Theobald DL (August 2013). "Relocating the active-site lysine in rhodopsin and implications for evolution of retinylidene proteins". Proceedings of the National Academy of Sciences of the United States of America. 110 (33): 13351–5. Bibcode:2013PNAS..11013351D. doi:10.1073/pnas.1306826110. PMC 3746867. PMID 23904486.
- Zhang Z, Jin Z, Zhao Y, Zhang Z, Li R, Xiao J, Wu J (August 2014). "Systematic study on G-protein couple receptor prototypes: did they really evolve from prokaryotic genes?". IET Systems Biology. 8 (4): 154–61. doi:10.1049/iet-syb.2013.0037. PMID 25075528.
- Nordström KJ, Sällman Almén M, Edstam MM, Fredriksson R, Schiöth HB (September 2011). "Independent HHsearch, Needleman--Wunsch-based, and motif analyses reveal the overall hierarchy for most of the G protein-coupled receptor families". Molecular Biology and Evolution. 28 (9): 2471–80. doi:10.1093/molbev/msr061. PMID 21402729.
- Yee DC, Shlykov MA, Västermark A, Reddy VS, Arora S, Sun EI, Saier MH (November 2013). "The transporter-opsin-G protein-coupled receptor (TOG) superfamily". The FEBS Journal. 280 (22): 5780–800. doi:10.1111/febs.12499. PMC 3832197. PMID 23981446.
- Yoshizawa S, Kumagai Y, Kim H, Ogura Y, Hayashi T, Iwasaki W, et al. (May 2014). "Functional characterization of flavobacteria rhodopsins reveals a unique class of light-driven chloride pump in bacteria". Proceedings of the National Academy of Sciences of the United States of America. 111 (18): 6732–7. Bibcode:2014PNAS..111.6732Y. doi:10.1073/pnas.1403051111. PMC 4020065. PMID 24706784.
- Grote M, Engelhard M, Hegemann P (May 2014). "Of ion pumps, sensors and channels - perspectives on microbial rhodopsins between science and history". Biochimica et Biophysica Acta (BBA) - Bioenergetics. 1837 (5): 533–45. doi:10.1016/j.bbabio.2013.08.006. PMID 23994288.
- Römpler H, Stäubert C, Thor D, Schulz A, Hofreiter M, Schöneberg T (February 2007). "G protein-coupled time travel: evolutionary aspects of GPCR research". Molecular Interventions. 7 (1): 17–25. doi:10.1124/mi.7.1.5. PMID 17339603.
- Zhang F, Vierock J, Yizhar O, Fenno LE, Tsunoda S, Kianianmomeni A, et al. (December 2011). "The microbial opsin family of optogenetic tools". Cell. 147 (7): 1446–57. doi:10.1016/j.cell.2011.12.004. PMC 4166436. PMID 22196724.
- Feuda R, Hamilton SC, McInerney JO, Pisani D (November 2012). "Metazoan opsin evolution reveals a simple route to animal vision". Proceedings of the National Academy of Sciences of the United States of America. 109 (46): 18868–72. Bibcode:2012PNAS..10918868F. doi:10.1073/pnas.1204609109. PMC 3503164. PMID 23112152.
- Porter ML, Blasic JR, Bok MJ, Cameron EG, Pringle T, Cronin TW, Robinson PR (January 2012). "Shedding new light on opsin evolution". Proceedings. Biological Sciences. 279 (1726): 3–14. doi:10.1098/rspb.2011.1819. PMC 3223661. PMID 22012981.
- Delroisse J, Ullrich-Lüter E, Ortega-Martinez O, Dupont S, Arnone MI, Mallefet J, Flammang P (November 2014). "High opsin diversity in a non-visual infaunal brittle star". BMC Genomics. 15 (1): 1035. doi:10.1186/1471-2164-15-1035. PMC 4289182. PMID 25429842.
- Shichida Y, Matsuyama T (October 2009). "Evolution of opsins and phototransduction". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 364 (1531): 2881–95. doi:10.1098/rstb.2009.0051. PMC 2781858. PMID 19720651.
- Gulati S, Jastrzebska B, Banerjee S, Placeres ÁL, Miszta P, Gao S, et al. (March 2017). "Photocyclic behavior of rhodopsin induced by an atypical isomerization mechanism". Proceedings of the National Academy of Sciences of the United States of America. 114 (13): E2608–E2615. doi:10.1073/pnas.1617446114. PMC 5380078. PMID 28289214.
- Terakita A (2005). "The opsins". Genome Biology. 6 (3): 213. doi:10.1186/gb-2005-6-3-213. PMC 1088937. PMID 15774036.
- Hunt DM, Carvalho LS, Cowing JA, Davies WL (October 2009). "Evolution and spectral tuning of visual pigments in birds and mammals". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 364 (1531): 2941–55. doi:10.1098/rstb.2009.0044. PMC 2781856. PMID 19720655.
- Trezise AE, Collin SP (October 2005). "Opsins: evolution in waiting". Current Biology. 15 (19): R794-6. doi:10.1016/j.cub.2005.09.025. PMID 16213808.
- Solomon SG, Lennie P (April 2007). "The machinery of colour vision" (PDF). Nature Reviews. Neuroscience. 8 (4): 276–86. doi:10.1038/nrn2094. PMID 17375040.
- "Color Space, Physical Space, and Fourier Transforms". Mathpages.
- "Seeing colour". Excerpts from "Theory of Color". Archived from the original on 2012-08-01.
- Okano T, Yoshizawa T, Fukada Y (November 1994). "Pinopsin is a chicken pineal photoreceptive molecule". Nature. 372 (6501): 94–7. Bibcode:1994Natur.372...94O. doi:10.1038/372094a0. PMID 7969427.
- Philp AR, Garcia-Fernandez JM, Soni BG, Lucas RJ, Bellingham J, Foster RG (June 2000). "Vertebrate ancient (VA) opsin and extraretinal photoreception in the Atlantic salmon (Salmo salar)". The Journal of Experimental Biology. 203 (Pt 12): 1925–36. PMID 10821749.
- Poletini MO, Ramos BC, Moraes MN, Castrucci AM (2015). "Nonvisual Opsins and the Regulation of Peripheral Clocks by Light and Hormones". Photochemistry and Photobiology. 91 (5): 1046–55. doi:10.1111/php.12494. PMID 26174318.
- Blackshaw S, Snyder SH (November 1997). "Parapinopsin, a novel catfish opsin localized to the parapineal organ, defines a new gene family". The Journal of Neuroscience. 17 (21): 8083–92. doi:10.1523/JNEUROSCI.17-21-08083.1997. PMC 6573767. PMID 9334384.
- Koyanagi M, Kawano E, Kinugawa Y, Oishi T, Shichida Y, Tamotsu S, Terakita A (April 2004). "Bistable UV pigment in the lamprey pineal". Proceedings of the National Academy of Sciences of the United States of America. 101 (17): 6687–91. Bibcode:2004PNAS..101.6687K. doi:10.1073/pnas.0400819101. PMC 404106. PMID 15096614.
- Koyanagi M, Wada S, Kawano-Yamashita E, Hara Y, Kuraku S, Kosaka S, et al. (September 2015). "Diversification of non-visual photopigment parapinopsin in spectral sensitivity for diverse pineal functions". BMC Biology. 13 (1): 73. doi:10.1186/s12915-015-0174-9. PMC 4570685. PMID 26370232.
- Su CY, Luo DG, Terakita A, Shichida Y, Liao HW, Kazmi MA, et al. (March 2006). "Parietal-eye phototransduction components and their potential evolutionary implications". Science. 311 (5767): 1617–21. Bibcode:2006Sci...311.1617S. doi:10.1126/science.1123802. PMID 16543463.
- Koyanagi M, Terakita A (May 2014). "Diversity of animal opsin-based pigments and their optogenetic potential". Biochimica et Biophysica Acta (BBA) - Bioenergetics. 1837 (5): 710–6. doi:10.1016/j.bbabio.2013.09.003. PMID 24041647.
- Haltaufderhyde K, Ozdeslik RN, Wicks NL, Najera JA, Oancea E (2015). "Opsin expression in human epidermal skin". Photochemistry and Photobiology. 91 (1): 117–23. doi:10.1111/php.12354. PMC 4303996. PMID 25267311.
- Blackshaw S, Snyder SH (May 1999). "Encephalopsin: a novel mammalian extraretinal opsin discretely localized in the brain". The Journal of Neuroscience. 19 (10): 3681–90. doi:10.1523/JNEUROSCI.19-10-03681.1999. PMC 6782724. PMID 10234000.
- Halford S, Freedman MS, Bellingham J, Inglis SL, Poopalasundaram S, Soni BG, et al. (March 2001). "Characterization of a novel human opsin gene with wide tissue expression and identification of embedded and flanking genes on chromosome 1q43". Genomics. 72 (2): 203–8. doi:10.1006/geno.2001.6469. PMID 11401433.
- Arendt D, Tessmar-Raible K, Snyman H, Dorresteijn AW, Wittbrodt J (October 2004). "Ciliary photoreceptors with a vertebrate-type opsin in an invertebrate brain". Science. 306 (5697): 869–71. Bibcode:2004Sci...306..869A. doi:10.1126/science.1099955. PMID 15514158.
- Tsukamoto H, Chen IS, Kubo Y, Furutani Y (August 2017). "A ciliary opsin in the brain of a marine annelid zooplankton is ultraviolet-sensitive, and the sensitivity is tuned by a single amino acid residue". The Journal of Biological Chemistry. 292 (31): 12971–12980. doi:10.1074/jbc.M117.793539. PMC 5546036. PMID 28623234.
- Ayers T, Tsukamoto H, Gühmann M, Veedin Rajan VB, Tessmar-Raible K (April 2018). "o-type opsin mediates the shadow reflex in the annelid Platynereis dumerilii". BMC Biology. 16 (1): 41. doi:10.1186/s12915-018-0505-8. PMC 5904973. PMID 29669554.
- Verasztó C, Gühmann M, Jia H, Rajan VB, Bezares-Calderón LA, Piñeiro-Lopez C, et al. (May 2018). "Ciliary and rhabdomeric photoreceptor-cell circuits form a spectral depth gauge in marine zooplankton". eLife. 7. doi:10.7554/eLife.36440. PMC 6019069. PMID 29809157.
- Gühmann M, Jia H, Randel N, Verasztó C, Bezares-Calderón LA, Michiels NK, et al. (August 2015). "Spectral Tuning of Phototaxis by a Go-Opsin in the Rhabdomeric Eyes of Platynereis". Current Biology. 25 (17): 2265–71. doi:10.1016/j.cub.2015.07.017. PMID 26255845.
- Nilsson DE (October 2009). "The evolution of eyes and visually guided behaviour". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 364 (1531): 2833–47. doi:10.1098/rstb.2009.0083. PMC 2781862. PMID 19720648.
- Nilsson DE (March 2013). "Eye evolution and its functional basis". Visual Neuroscience. 30 (1–2): 5–20. doi:10.1017/S0952523813000035. PMC 3632888. PMID 23578808.
- Koyanagi M, Takada E, Nagata T, Tsukamoto H, Terakita A (March 2013). "Homologs of vertebrate Opn3 potentially serve as a light sensor in nonphotoreceptive tissue". Proceedings of the National Academy of Sciences of the United States of America. 110 (13): 4998–5003. Bibcode:2013PNAS..110.4998K. doi:10.1073/pnas.1219416110. PMC 3612648. PMID 23479626.
- Moutsaki P, Whitmore D, Bellingham J, Sakamoto K, David-Gray ZK, Foster RG (April 2003). "Teleost multiple tissue (tmt) opsin: a candidate photopigment regulating the peripheral clocks of zebrafish?". Brain Research. Molecular Brain Research. 112 (1–2): 135–45. doi:10.1016/S0169-328X(03)00059-7. PMID 12670711.
- Fischer RM, Fontinha BM, Kirchmaier S, Steger J, Bloch S, Inoue D, et al. (11 June 2013). "Co-expression of VAL- and TMT-opsins uncovers ancient photosensory interneurons and motorneurons in the vertebrate brain". PLOS Biology. 11 (6): e1001585. doi:10.1371/journal.pbio.1001585. PMC 3679003. PMID 23776409.
- Sakai K, Yamashita T, Imamoto Y, Shichida Y (22 October 2015). "Diversity of Active States in TMT Opsins". PLOS ONE. 10 (10): e0141238. Bibcode:2015PLoSO..1041238S. doi:10.1371/journal.pone.0141238. PMC 4619619. PMID 26491964.
- Koyanagi M, Takano K, Tsukamoto H, Ohtsu K, Tokunaga F, Terakita A (October 2008). "Jellyfish vision starts with cAMP signaling mediated by opsin-G(s) cascade". Proceedings of the National Academy of Sciences of the United States of America. 105 (40): 15576–80. Bibcode:2008PNAS..10515576K. doi:10.1073/pnas.0806215105. PMC 2563118. PMID 18832159.
- Liegertová M, Pergner J, Kozmiková I, Fabian P, Pombinho AR, Strnad H, et al. (July 2015). "Cubozoan genome illuminates functional diversification of opsins and photoreceptor evolution". Scientific Reports. 5: 11885. Bibcode:2015NatSR...511885L. doi:10.1038/srep11885. PMC 5155618. PMID 26154478.
- Mason B, Schmale M, Gibbs P, Miller MW, Wang Q, Levay K, et al. (5 December 2012). "Evidence for multiple phototransduction pathways in a reef-building coral". PLOS ONE. 7 (12): e50371. Bibcode:2012PLoSO...750371M. doi:10.1371/journal.pone.0050371. PMC 3515558. PMID 23227169.
- Suga H, Schmid V, Gehring WJ (January 2008). "Evolution and functional diversity of jellyfish opsins". Current Biology. 18 (1): 51–5. doi:10.1016/j.cub.2007.11.059. PMID 18160295.
- Feuda R, Rota-Stabelli O, Oakley TH, Pisani D (July 2014). "The comb jelly opsins and the origins of animal phototransduction". Genome Biology and Evolution. 6 (8): 1964–71. doi:10.1093/gbe/evu154. PMC 4159004. PMID 25062921.
- Murakami M, Kouyama T (May 2008). "Crystal structure of squid rhodopsin". Nature. 453 (7193): 363–7. Bibcode:2008Natur.453..363M. doi:10.1038/nature06925. PMID 18480818.
- Smith WC, Price DA, Greenberg RM, Battelle BA (July 1993). "Opsins from the lateral eyes and ocelli of the horseshoe crab, Limulus polyphemus". Proceedings of the National Academy of Sciences of the United States of America. 90 (13): 6150–4. Bibcode:1993PNAS...90.6150S. doi:10.1073/pnas.90.13.6150. PMC 46885. PMID 8327495.
- Hering L, Mayer G (September 2014). "Analysis of the opsin repertoire in the tardigrade Hypsibius dujardini provides insights into the evolution of opsin genes in panarthropoda". Genome Biology and Evolution. 6 (9): 2380–91. doi:10.1093/gbe/evu193. PMC 4202329. PMID 25193307.
- Kojima D, Terakita A, Ishikawa T, Tsukahara Y, Maeda A, Shichida Y (September 1997). "A novel Go-mediated phototransduction cascade in scallop visual cells". The Journal of Biological Chemistry. 272 (37): 22979–82. doi:10.1074/jbc.272.37.22979. PMID 9287291.
- Koyanagi M, Terakita A, Kubokawa K, Shichida Y (November 2002). "Amphioxus homologs of Go-coupled rhodopsin and peropsin having 11-cis- and all-trans-retinals as their chromophores". FEBS Letters. 531 (3): 525–8. doi:10.1016/s0014-5793(02)03616-5. PMID 12435605.
- Jiang M, Pandey S, Fong HK (December 1993). "An opsin homologue in the retina and pigment epithelium". Investigative Ophthalmology & Visual Science. 34 (13): 3669–78. PMID 8258527.
- Hao W, Fong HK (March 1999). "The endogenous chromophore of retinal G protein-coupled receptor opsin from the pigment epithelium". The Journal of Biological Chemistry. 274 (10): 6085–90. doi:10.1074/jbc.274.10.6085. PMID 10037690.
- Nagata T, Koyanagi M, Terakita A (20 October 2010). "Molecular Evolution and Functional Diversity of Opsin-Based Photopigments". Retrieved 7 May 2018. Cite journal requires
|journal=(help) - Wenzel A, Oberhauser V, Pugh EN, Lamb TD, Grimm C, Samardzija M, et al. (August 2005). "The retinal G protein-coupled receptor (RGR) enhances isomerohydrolase activity independent of light". The Journal of Biological Chemistry. 280 (33): 29874–84. doi:10.1074/jbc.M503603200. PMID 15961402.
- Radu RA, Hu J, Peng J, Bok D, Mata NL, Travis GH (July 2008). "Retinal pigment epithelium-retinal G protein receptor-opsin mediates light-dependent translocation of all-trans-retinyl esters for synthesis of visual chromophore in retinal pigment epithelial cells". The Journal of Biological Chemistry. 283 (28): 19730–8. doi:10.1074/jbc.M801288200. PMC 2443657. PMID 18474598.
- Hao W, Fong HK (May 1996). "Blue and ultraviolet light-absorbing opsin from the retinal pigment epithelium". Biochemistry. 35 (20): 6251–6. doi:10.1021/bi952420k. PMID 8639565.
- Sun H, Gilbert DJ, Copeland NG, Jenkins NA, Nathans J (September 1997). "Peropsin, a novel visual pigment-like protein located in the apical microvilli of the retinal pigment epithelium". Proceedings of the National Academy of Sciences of the United States of America. 94 (18): 9893–8. Bibcode:1997PNAS...94.9893S. doi:10.1073/pnas.94.18.9893. PMC 23288. PMID 9275222.
- Kojima D, Mori S, Torii M, Wada A, Morishita R, Fukada Y (17 October 2011). "UV-sensitive photoreceptor protein OPN5 in humans and mice". PLOS ONE. 6 (10): e26388. Bibcode:2011PLoSO...626388K. doi:10.1371/journal.pone.0026388. PMC 3197025. PMID 22043319.
- Yamashita T, Ohuchi H, Tomonari S, Ikeda K, Sakai K, Shichida Y (December 2010). "Opn5 is a UV-sensitive bistable pigment that couples with Gi subtype of G protein". Proceedings of the National Academy of Sciences of the United States of America. 107 (51): 22084–9. Bibcode:2010PNAS..10722084Y. doi:10.1073/pnas.1012498107. PMC 3009823. PMID 21135214.
- Buhr ED, Yue WW, Ren X, Jiang Z, Liao HW, Mei X, et al. (October 2015). "Neuropsin (OPN5)-mediated photoentrainment of local circadian oscillators in mammalian retina and cornea". Proceedings of the National Academy of Sciences of the United States of America. 112 (42): 13093–8. Bibcode:2015PNAS..11213093B. doi:10.1073/pnas.1516259112. PMC 4620855. PMID 26392540.
- Mano H, Kojima D, Fukada Y (November 1999). "Exo-rhodopsin: a novel rhodopsin expressed in the zebrafish pineal gland". Brain Research. Molecular Brain Research. 73 (1–2): 110–8. doi:10.1016/S0169-328X(99)00242-9. PMID 10581404.
- Tarttelin EE, Fransen MP, Edwards PC, Hankins MW, Schertler GF, Vogel R, et al. (November 2011). "Adaptation of pineal expressed teleost exo-rod opsin to non-image forming photoreception through enhanced Meta II decay". Cellular and Molecular Life Sciences. 68 (22): 3713–23. doi:10.1007/s00018-011-0665-y. PMC 3203999. PMID 21416149.
- Palczewski K, Kumasaka T, Hori T, Behnke CA, Motoshima H, Fox BA, et al. (August 2000). "Crystal structure of rhodopsin: A G protein-coupled receptor". Science. 289 (5480): 739–45. Bibcode:2000Sci...289..739P. CiteSeerX 10.1.1.1012.2275. doi:10.1126/science.289.5480.739. PMID 10926528.
- Yokoyama S (July 2000). "Molecular evolution of vertebrate visual pigments". Progress in Retinal and Eye Research. 19 (4): 385–419. doi:10.1016/S1350-9462(00)00002-1. PMID 10785616.
- Deeb SS (May 2005). "The molecular basis of variation in human color vision". Clinical Genetics. 67 (5): 369–77. doi:10.1111/j.1399-0004.2004.00343.x. PMID 15811001.
- White JH, Chiano M, Wigglesworth M, Geske R, Riley J, White N, et al. (July 2008). "Identification of a novel asthma susceptibility gene on chromosome 1qter and its functional evaluation". Human Molecular Genetics. 17 (13): 1890–903. doi:10.1093/hmg/ddn087. PMID 18344558.
- Nissilä J, Mänttäri S, Särkioja T, Tuominen H, Takala T, Timonen M, Saarela S (November 2012). "Encephalopsin (OPN3) protein abundance in the adult mouse brain". Journal of Comparative Physiology A. 198 (11): 833–9. doi:10.1007/s00359-012-0754-x. PMC 3478508. PMID 22991144.
- Bailes HJ, Lucas RJ (May 2013). "Human melanopsin forms a pigment maximally sensitive to blue light (λmax ≈ 479 nm) supporting activation of G(q/11) and G(i/o) signalling cascades". Proceedings. Biological Sciences. 280 (1759): 20122987. doi:10.1098/rspb.2012.2987. PMC 3619500. PMID 23554393.
- Tarttelin EE, Bellingham J, Hankins MW, Foster RG, Lucas RJ (November 2003). "Neuropsin (Opn5): a novel opsin identified in mammalian neural tissue". FEBS Letters. 554 (3): 410–6. doi:10.1016/S0014-5793(03)01212-2. PMID 14623103.
- Yamashita T, Ono K, Ohuchi H, Yumoto A, Gotoh H, Tomonari S, et al. (February 2014). "Evolution of mammalian Opn5 as a specialized UV-absorbing pigment by a single amino acid mutation". The Journal of Biological Chemistry. 289 (7): 3991–4000. doi:10.1074/jbc.M113.514075. PMC 3924266. PMID 24403072.