Octopamine
Octopamine (molecular formula C8H11NO2; also known as para-octopamines and others) is an organic chemical closely related to norepinephrine, and synthesized biologically by a homologous pathway. Its name derives from the fact that it was first identified in the salivary glands of the octopus.
 | |
-Octopamine_molecule_ball.png) | |
| Clinical data | |
|---|---|
| Other names | OCT, Norsympathol, Norsynephrine, para-Octopamine, beta-Hydroxytyramine, para-hydroxy-phenyl-ethanolamine, α-(Aminomethyl)-4 hydroxybenzenemethanol, 1-(p-Hydroxyphenyl)-2-aminoethanol |
| Routes of administration | Oral |
| ATC code | |
| Physiological data | |
| Source tissues | invertebrate nervous systems; trace amine in vertebrates |
| Target tissues | system-wide in invertebrates |
| Receptors | TAAR1 (mammals) OctαR, OctβR, TyrR (invertebrates), Oct-TyrR |
| Agonists | Formamidines (amitraz (AMZ) and chlordimeform (CDM)) |
| Antagonists | epinastine (3-amino-9, 13b-dihydro-1H-dibenz(c,f)imidazo(1,5a)azepine hydrochloride) |
| Precursor | tyramine |
| Biosynthesis | tyramine β-hydroxylase; dopamine β-hydroxylase |
| Metabolism | p-hydroxymandelic acid;[1][2] N-acetyltransferases; phenylethanolamine N-methyltransferase |
| Legal status | |
| Legal status |
|
| Pharmacokinetic data | |
| Bioavailability | 99.42 % |
| Metabolism | p-hydroxymandelic acid;[1][3] N-acetyltransferases; phenylethanolamine N-methyltransferase |
| Elimination half-life | 15 minutes in insects. Between 76 and 175 minutes in humans |
| Excretion | Up to 93% of ingested octopamine is eliminated via the urinary route within 24 hours[1] |
| Identifiers | |
| |
| CAS Number | |
| PubChem CID | |
| IUPHAR/BPS | |
| ChemSpider | |
| UNII | |
| ChEBI | |
| ChEMBL | |
| CompTox Dashboard (EPA) | |
| ECHA InfoCard | 100.002.890 |
| Chemical and physical data | |
| Formula | C8H11NO2 |
| Molar mass | 153.181 g·mol−1 |
| 3D model (JSmol) | |
| |
| |
| | |
In many types of invertebrates octopamine is an important neurotransmitter and hormone. In protostomes — arthropods, molluscs, and several types of worms — it substitutes for norephinephrine and performs functions apparently similar to those of norepinephrine in mammals, functions that have been described as mobilizing the body and nervous system for action. In mammals octopamine is found only in trace amounts, and no biological function has been solidly established for it. It is also found naturally in numerous plants, including bitter orange.[4][5] Octopamine has been sold under trade names such as Epirenor, Norden, and Norfen for use as a sympathomimetic drug, available by prescription.
Functions
Cellular effects
Octopamine exerts its effects by binding to and activating receptors located on the surface of cells. These receptors have mainly been studied in insects, where they can be divided into three types: alpha-adrenergic-like (OctαR), which are structurally and functionally similar to noradrenergic alpha-1 receptors in mammals; beta-adrenergic-like (OctβR), which are structurally and functionally similar to noradrenergic beta receptors in mammals; and mixed octopamine/tyramine receptors (TyrR), which are structurally and functionally similar to noradrenergic alpha-2 receptors in mammals.[6] Receptors in the TyrR class, however, are generally more strongly activated by tyramine than by octopamine.[6]
In vertebrates no octopamine-specific receptors have been identified. Octopamine binds weakly to receptors for norepinephrine and epinephrine, but it is not clear whether this has any functional significance. It binds more strongly to trace amine-associated receptors (TAARs), especially TAAR1.[6]
Invertebrates
Octopamine was first discovered by Italian scientist Vittorio Erspamer in 1948[7] in the salivary glands of the octopus and has since been found to act as a neurotransmitter, neurohormone and neuromodulator in invertebrates. Although Erspamer discovered its natural occurrence and named it, octopamine had actually existed for many years as a pharmaceutical product.[8] It is widely used in energy-demanding behaviors by all insects, crustaceans (crabs, lobsters, crayfish), and spiders. Such behaviors include flying,[9] egg-laying,[10] and jumping.[11][12]
Octopamine acts as the insect equivalent of norepinephrine and has been implicated in regulating aggression in invertebrates, with different effects on different species. Studies have shown that reducing the neurotransmitter octopamine and preventing coding of tyramine beta hydroxylase (an enzyme that converts tyramine to octopamine) decreases aggression in Drosophila without influencing other behaviors.[13]
In insects, octopamine is released by a select number of neurons, but acts broadly throughout the central brain, on all sense organs, and on several non-neuronal tissues.[14][15] In the thoracic ganglia, octopamine is primarily released by DUM (dorsal unpaired median) and VUM (ventral unpaired median) neurons, which release octopamine onto neural, muscular, and peripheral targets.[16][17] These neurons are important for mediating energy-demanding motor behaviors, such as escape-induced jumping and flight. For example, the locust DUMeti neuron releases octopamine onto the extensor tibia muscle to increase muscle tension and increase relaxation rate. These actions promote efficient leg muscle contraction for jumping.[14] During flight, DUM neurons are also active and release octopamine throughout the body to synchronize energy metabolism, respiration, muscle activity and flight interneuron activity.[9]
In the honey bee and fruit fly, octopamine has a major role in learning and memory. In the firefly, octopamine release leads to light production in the lantern.[18][19]
In lobsters, octopamine seems to direct and coordinate neurohormones to some extent in the central nervous system, and it was observed that injecting octopamine into a lobster and crayfish resulted in limb and abdomen extension.[20]
Heberlein et al.[21] have conducted studies of alcohol tolerance in fruit flies; they found that a mutation that caused octopamine deficiency also caused lower alcohol tolerance.[22][23][24][25]
The emerald cockroach wasp stings the host for its larvae (a cockroach) in the head ganglion (brain). The venom blocks octopamine receptors[26] and the cockroach fails to show normal escape responses, grooming itself excessively. It becomes docile and the wasp leads it to the wasp's den by pulling its antenna like a leash.[27]
Vertebrates
In vertebrates, octopamine replaces norepinephrine in sympathetic neurons with chronic use of monoamine oxidase inhibitors. It may be responsible for the common side effect of orthostatic hypotension with these agents, though there is also evidence that it is actually mediated by increased levels of N-acetylserotonin.
One study noted that octopamine might be an important amine that influences the therapeutic effects of inhibitors such as monoamine oxidase inhibitors, especially because a large increase in octopamine levels was observed when animals were treated with this inhibitor. Octopamine was positively identified in the urine samples of mammals such as humans, rats, and rabbits treated with monoamine oxidase inhibitors. Very small amounts of octopamine were also found in certain animal tissues. It was observed that within a rabbit's body, the heart and kidney held the highest concentrations of octopamine. Octopamine was found to be 93% eluted by urine within 24 hours of being produced in the body as a byproduct of Iproniazid in rabbits.[8]
Pharmacology
Octopamine has been sold under trade names such as Epirenor, Norden, and Norfen for use in medicine as a sympathomimetic drug, available by prescription. Very little information exists concerning its clinical usefulness or safety, though.[28]
In mammals, octopamine may mobilize the release of fat from adipocytes (fat cells), which has led to its promotion on the internet as a slimming aid. However, the released fat is likely to be promptly taken up into other cells, and there is no evidence that octopamine facilitates weight loss. Octopamine may also increase blood pressure significantly when combined with other stimulants, as in some weight loss supplements.[29]
The World Anti-Doping Agency lists octopamine as a banned substance for in competition use, as a "specified stimulant"[30] on the 2019 Prohibited List.
Biochemical mechanisms
Mammals
Octopamine is one of four primary endogenous agonists of human trace amine-associated receptor 1.[31]
Invertebrates
Octopamine binds to its respective G-protein coupled receptors (GPCRs) to initiate a cell signal transduction pathway. At least three groups of octopamine GPCR have been defined. OctαR (OCTOPAMINE1 receptors) are more closely related to α-adrenergic receptors, while OctβR (OCTOPAMINE2 receptors) are more closely related to β-adrenergic receptors. The Octopamine/Tyramine receptors (including Oct-TyrR) can bind both ligands, and display agonist-specific coupling. Oct-TyrR is listed in both OCTOPAMINE and TYRAMINE RECEPTORS gene groups.[32]
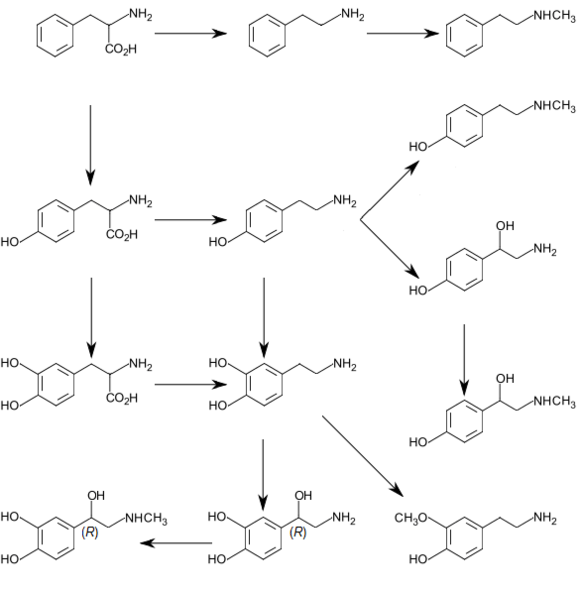
Biosynthesis
In humans
See also
References
- Hengstmann, J. H.; Konen, W; Konen, C; Eichelbaum, M; Dengler, H. J. (1974). "The physiological disposition of p-octopamine in man". Naunyn-Schmiedeberg's Archives of Pharmacology. 283 (1): 93–106. doi:10.1007/bf00500148. PMID 4277715.
- d’Andrea, Giovanni; Nordera, Gianpietro; Pizzolato, Gilberto; Bolner, Andrea; Colavito, Davide; Flaibani, Raffaella; Leon, Alberta (2010). "Trace amine metabolism in Parkinson's disease: Low circulating levels of octopamine in early disease stages". Neuroscience Letters. 469 (3): 348–51. doi:10.1016/j.neulet.2009.12.025. PMID 20026245.
- d’Andrea, Giovanni; Nordera, Gianpietro; Pizzolato, Gilberto; Bolner, Andrea; Colavito, Davide; Flaibani, Raffaella; Leon, Alberta (2010). "Trace amine metabolism in Parkinson's disease: Low circulating levels of octopamine in early disease stages". Neuroscience Letters. 469 (3): 348–51. doi:10.1016/j.neulet.2009.12.025. PMID 20026245.
- Tang F, Tao L, Luo X, Ding L, Guo M, Nie L, Yao S (September 2006). "Determination of octopamine, synephrine and tyramine in Citrus herbs by ionic liquid improved 'green' chromatography". Journal of Chromatography A. 1125 (2): 182–8. doi:10.1016/j.chroma.2006.05.049. PMID 16781718.
- Jagiełło-Wójtowicz E (1979). "Mechanism of central action of octopamine". Polish Journal of Pharmacology and Pharmacy. 31 (5): 509–16. PMID 121158.
- Pflüger HJ, Stevensonb PA (2005). "Evolutionary aspects of octopaminergic systems with emphasis on arthropods". Arthropod Structure & Development. 34 (3): 379–396. doi:10.1016/j.asd.2005.04.004.CS1 maint: uses authors parameter (link)
- Erspamer, V. (2009). "Active Substances in the Posterior Salivary Glands of Octopoda. II. Tyramine and Octopamine (Oxyoctopamine)". Acta Pharmacologica et Toxicologica. 4 (3–4): 224–47. doi:10.1111/j.1600-0773.1948.tb03345.x.
- Kakimoto Y, Armstrong MD (February 1962). "On the identification of octopamine in mammals". The Journal of Biological Chemistry. 237: 422–7. PMID 14453200.
- Orchard, I; Ramirez, J M; Lange, A B (January 1993). "A Multifunctional Role for Octopamine in Locust Flight". Annual Review of Entomology. 38 (1): 227–249. doi:10.1146/annurev.en.38.010193.001303. ISSN 0066-4170.
- Lee, Hyun-Gwan; Seong, Chang-Soo; Kim, Young-Cho; Davis, Ronald L; Han, Kyung-An (1 December 2003). "Octopamine receptor OAMB is required for ovulation in Drosophila melanogaster". Developmental Biology. 264 (1): 179–190. doi:10.1016/j.ydbio.2003.07.018. ISSN 0012-1606.
- Pollack, Alan J.; Ritzmann, Roy E.; Westin, Joanne (1988). "Activation of DUM cell interneurons by ventral giant interneurons in the cockroach, periplaneta americana". Journal of Neurobiology. 19 (6): 489–497. doi:10.1002/neu.480190602. ISSN 1097-4695.
- Orchard, Ian (1 April 1982). "Octopamine in insects: neurotransmitter, neurohormone, and neuromodulator". Canadian Journal of Zoology. 60 (4): 659–669. doi:10.1139/z82-095. ISSN 0008-4301.
- Zhou C, Rao Y, Rao Y (September 2008). "A subset of octopaminergic neurons are important for Drosophila aggression". Nature Neuroscience. 11 (9): 1059–67. doi:10.1038/nn.2164. PMID 19160504.
- Atwood, H. L.; Klose, M. K. (1 January 2009), Squire, Larry R. (ed.), "Neuromuscular Transmission Modulation at Invertebrate Neuromuscular Junctions", Encyclopedia of Neuroscience, Oxford: Academic Press, pp. 671–690, ISBN 978-0-08-045046-9, retrieved 10 July 2020
- Roeder, T. (December 1999). "Octopamine in invertebrates". Progress in Neurobiology. 59 (5): 533–561. doi:10.1016/s0301-0082(99)00016-7. ISSN 0301-0082. PMID 10515667.
- Eckert, Manfred; Rapus, Jürgen; Nürnberger, Asja; Penzlin, Heinz (1992). "A new specific antibody reveals octopamine-like immunoreactivity in cockroach ventral nerve cord". Journal of Comparative Neurology (in French). 322 (1): 1–15. doi:10.1002/cne.903220102. ISSN 1096-9861.
- Sinakevitch, Irina G.; Geffard, Michel; Pelhate, Marcel; Lapied, Bruno (April 1994). "Octopamine-like immunoreactivity in the dorsal unpaired median (DUM) neurons innervating the accessory gland of the male cockroach Periplaneta americana". Cell and Tissue Research. 276 (1): 15–21. doi:10.1007/bf00354779. ISSN 0302-766X.
- Greenfield MD (November 2001). "Missing link in firefly bioluminescence revealed: NO regulation of photocyte respiration". BioEssays. 23 (11): 992–5. doi:10.1002/bies.1144. PMID 11746215.
- Trimmer BA, Aprille JR, Dudzinski DM, Lagace CJ, Lewis SM, Michel T, et al. (June 2001). "Nitric oxide and the control of firefly flashing". Science. 292 (5526): 2486–8. doi:10.1126/science.1059833. PMID 11431567.
- Livingstone MS, Harris-Warrick RM, Kravitz EA (April 1980). "Serotonin and octopamine produce opposite postures in lobsters". Science. 208 (4439): 76–9. Bibcode:1980Sci...208...76L. doi:10.1126/science.208.4439.76. PMID 17731572.
- Heberlein U, Wolf FW, Rothenfluh A, Guarnieri DJ (August 2004). "Molecular Genetic Analysis of Ethanol Intoxication in Drosophila melanogaster". Integrative and Comparative Biology. 44 (4): 269–74. CiteSeerX 10.1.1.536.262. doi:10.1093/icb/44.4.269. PMID 21676709.
- Moore MS, DeZazzo J, Luk AY, Tully T, Singh CM, Heberlein U (June 1998). "Ethanol intoxication in Drosophila: Genetic and pharmacological evidence for regulation by the cAMP signaling pathway". Cell. 93 (6): 997–1007. doi:10.1016/S0092-8674(00)81205-2. PMID 9635429.
- Tecott LH, Heberlein U (December 1998). "Y do we drink?". Cell. 95 (6): 733–5. doi:10.1016/S0092-8674(00)81695-5. PMID 9865690.
- Williams, Ruth (22 June 2005). "Bar Flies: What our insect relatives can teach us about alcohol tolerance". Naked Scientist.
- Vince, Gaia (22 August 2005). "'Hangover gene' is key to alcohol tolerance". New Scientist.
- Hopkin, Michael (2007). "How to make a zombie cockroach". Nature. doi:10.1038/news.2007.312.
- Gal R, Rosenberg LA, Libersat F (December 2005). "Parasitoid wasp uses a venom cocktail injected into the brain to manipulate the behavior and metabolism of its cockroach prey". Archives of Insect Biochemistry and Physiology. 60 (4): 198–208. doi:10.1002/arch.20092. PMID 16304619.
- Stohs SJ (January 2015). "Physiological functions and pharmacological and toxicological effects of p-octopamine". Drug and Chemical Toxicology. 38 (1): 106–12. doi:10.3109/01480545.2014.900069. PMID 24654910.
- Haller CA, Benowitz NL, Jacob P (September 2005). "Hemodynamic effects of ephedra-free weight-loss supplements in humans". The American Journal of Medicine. 118 (9): 998–1003. doi:10.1016/j.amjmed.2005.02.034. PMID 16164886.
- "Prohibited In Competition – Stimulants". WADA. Retrieved 6 May 2019.
- Maguire JJ, Davenport AP (20 February 2018). "Trace amine receptor: TA1 receptor". IUPHAR/BPS Guide to PHARMACOLOGY. International Union of Basic and Clinical Pharmacology. Retrieved 16 July 2018.
- "Gene Group : OCTOPAMINE RECEPTORS", FlyBase, October 16, 2018.
- Broadley KJ (March 2010). "The vascular effects of trace amines and amphetamines". Pharmacology & Therapeutics. 125 (3): 363–375. doi:10.1016/j.pharmthera.2009.11.005. PMID 19948186.
- Lindemann L, Hoener MC (May 2005). "A renaissance in trace amines inspired by a novel GPCR family". Trends in Pharmacological Sciences. 26 (5): 274–281. doi:10.1016/j.tips.2005.03.007. PMID 15860375.
- Wang X, Li J, Dong G, Yue J (February 2014). "The endogenous substrates of brain CYP2D". European Journal of Pharmacology. 724: 211–218. doi:10.1016/j.ejphar.2013.12.025. PMID 24374199.