Formyl peptide receptor 1
Formyl peptide receptor 1 (FPR1, FPR1 receptor, fMet-Leu-Phe receptor 1, FMLP receptor 1, or N-formylmethionyl-leucyl-phenylalanine receptor 1) is a cell surface receptor protein that in humans is encoded by the formyl peptide receptor 1 (FPR1) gene. This gene encodes a G protein-coupled receptor cell surface protein that binds and is activated by N-Formylmethionine-containing oligopeptides, particularly N-Formylmethionine-leucyl-phenylalanine (FMLP). FPR1 is prominently expressed by mammalian phagocytic and blood leukocyte cells where it functions to mediate these cells' responses to the N-formylmethionine-containing oligopeptides which are released by invading microorganisms and injured tissues. FPR1 directs these cells to sites of invading pathogens or disrupted tissues and then stimulates these cells to kill the pathogens or to remove tissue debris; as such, it is an important component of the innate immune system that operates in host defense and damage control.[5]


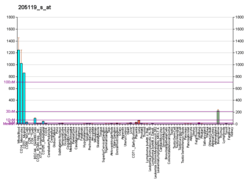
Humans also express two paralogs of FPR1 vis., FPR2 and FPR3. Mice express no fewer than 7 Fpr receptors and encoding genes that are homologous to FPR1 although no single one of these FPRs appears to perform exactly the same functions as any one of the human FPRs.[6]
Function
FPR1 binds with and is activated by:
- bacterial and mitochondrial N-formyl peptides and thereby initiates innate host immune responses.
- various synthetic N-formyl and non-formylated peptides that show distinguishing differences from those that interact with FPR2 and FPR3.
- T20/DP178 & T21/DP107, N-acetylated polypeptides derived from the gp41 HIV-1 envelope protein. This interaction is of unknown physiological significance although peptide T20/DP178 is a licensed anti-retrovirus agent (pentafuside) termed Enfuvirtide which acts at the level of HIV-target cell fusion and is used clinically to treat HIV-1 infection).
- Annexin A1 (also termed ANXA1 and lipocortin 1) and its N-terminal peptides (Ac2–26 and Ac9–25). At low concentrations, these agents stimulate neutrophils to raise cytosolic Ca2+ levels and thereby activate Ca2+-dependent signaling pathways; however, they do not fully activate the MAPK pathway but rather leave the neutrophil desensitized (i.e. unresponsive) to chemokine IL-8. At high concentrations, in contrast, the agents fully activate neutrophils and are potent pro-inflammatory stimulants.[7]
History
Studies conducted in the 1970s found that a series of N-formylmethionine-containing oligopeptides, including the most potent and best known member of this series, N-Formylmethionine-leucyl-phenylalanine (FMLP or fMet-Leu-Phe), stimulated rabbit and human neutrophils by an apparent receptor-dependent mechanism to migrate in a directional pattern in classical laboratory assays of chemotaxis. Since these oligopeptides were produced by bacteria or synthetic analogs of such products, it was suggested that the N-formyl oligopeptides are important chemotatic factors and their receptors are important chemotactic factor receptors that act respectively as signaling and signal-recognizing elements to initiate Inflammation responses in order to defend against bacterial invasion. Further studies cloned a receptor for these N-formyl oligopeptides, FPR1. Two receptors where thereafter discovered and named FPR2 and FPR3 based on the similarity of their genes' predicted amino acid sequence to that of FPR1 rather than on any ability to bind or be activated by the formyl oligopeptides. The latter two receptors were subsequently found to have very different specificities for the formyl oligopeptides and very different functions than those for FPR1. FPR1 is the premiere receptor for the pro-inflammatory actions of formyl peptides.[5][8][9]
Nomenclature
Confusingly, there are two nomenclatures for FPR receptors and their genes, the first one used, FPR, FPR1, and FPR2, and its replacement (which corresponds directly to these three respective receptors and their genes), FPR1, FPR2, and FPR3. The latter nomenclature was recommended by the International Union of Basic and Clinical Pharmacology[7] and is used here. Other previously used names for FPR1 are NFPR, and FMLPR; for FPR2 are FPRH1, FPRL1, RFP, LXA4R, ALXR, FPR2/ALX, HM63, FMLPX, FPR2A, and ALX/FPR2 (most recently, ALX/FPR2 is commonly used for FPR2); and for FPR3 are FPRH2, FPRL2, and FMLPY.[7]
Gene
Human
In early studies, cultured human HL-60 promyelocytes purposely differentiated to granulocytes were used to partially purify[10] and in a series of experiments clone FPR1;[11][12][13][14][15] an apparent homolog of FPR1, Fpr was also cloned from rabbit neutrophils.[16] The studies indicated that FPR1 is a G protein-coupled receptor that activates cells though a linkage to the pertussis toxin-sensitive Gαi subclass of G proteins, that FPR1 is located on chromosome 19q.13.3, and that this gene consists of two exons, the first of which encodes a 66 base pair 5'-untranslated sequence, the second of which has an intronless open reading frame coding for a protein containing ~354 amino acids; the studies also indicated that cells express multiple formyl peptide receptor mRNA transcripts due to Allelic heterogeneity, alternate Polyadenylation sites, and possibly products of other genes with homology to FPR1. Subsequent studies cloned two other genes with homology to FPR1 viz., FPL2 (originally termed FPR1, FPRH1, or FPRL1) and FPR3 (originally termed FPR2, FPRH2, or FPRL2).[17][18][19] FPR2 and FPR3 are composed of 351 and 352 amino acids, respectively, and similar to FPR1 have intronless open reading frames which encode G protein coupled receptors; FPR1 and FPR2 have 66% and 56% amino acid sequence identity with FPR1 and 72% homology to each other.[7] All three genes localize to chromosome 19q.13.3 in the order of FPR1, FPR2, and FPR3 to form a cluster which also includes the gene for another G protein-coupled chemotactic factor receptor, the C5a receptor (also termed CD88), which binds and is activated by complement component 5a (C5a) and GPR77, a second C5a anaphylatoxin chemotactic receptor C5a2 (C5L2), a second C5a receptor of debated function which has the structure of a G protein coupled receptor but fails to couple to G proteins.[20] These points are of interest because C5a is generated by the interaction of bacteria with blood plasma components to activate the complement cascade which then cleave C5a from Complement component 5. Thus, bacteria produce a family of oligopeptide chemotactic factors plus activate host complement pathways to generate C5a, which, like the formylated oligopeptides, is a neutrophil chemotactic factor that operates through receptors whose genes cluster with those for the three formly peptide receptors.[21] Furthermore, bacteria-induced complement activation also causes the formation of complement component 3a (C3a) by cleavage from complement component 3; C3a is a neutrophil chemotactic factor which operates through a G protein coupled chemotactic factor receptor, the C3a receptor, whose gene is located at chromosome 12p13; C3a also acts through C5L2.[20][22]
Mouse
Mouse formyl peptide receptor genes localize to chromosome 17A3.2 in the following order: Fpr1, Fpr-rs2 (or fpr2), Fpr-rs1 (or Lxa4R), Fpr-rs4, Fpr-rs7, Fpr-rs6, and Fpr-rs3; Pseudogenes ψFpr-rs2 and ψFpr-rs3 (or ψFpr-rs5) lie just after Fpr-rs2 and Fpr-rs1, respectively. All of the active mouse FPR receptors have ≥50% amino acid sequence identity with each other as well as with the three human FPR receptors.[6] Studies find that: a) mouse Fpr1 is an ortholog of human FPR1, responding to many bacterial- and mitochondrial-derived formyl peptides but only minimally to FMLP and having certain pharmacologic properties in common with human FPR2/ALX; b) mouse Fpr2 and mFpr-rs1 bind with high affinity and respond to lipoxins but have little affinity for or responsiveness to formyl peptides and therefore share key properties with human FPR2/ALX; and c) based on its predominantly intracellular distribution, mFpr-rs1 correlates, and therefore may share functionally, with human FPR3;[23][24][25]
The ψFpr-rs2 gene contains a deletion and frame shift which renders its protein 186 nucleotides shorter but 98% identical to the protein encoded by its closest paralog gene, Fpr-rs2. Since ψFpr-rs2 transcripts are expressed and inducible in multiple mouse tissues and since gene knockout studies ascribe functionality to it, ψFpr-rs2 may not a true pseudogene and, it is suggested, should be renamed Fpr-rs8.[26]
Fpr-rs1, Fpr-rs3, Fpr-rs4, Fpr-rs6, and Fpr-rs7 receptors are expressed in the olfactory bulb sensory neurons of the Vomeronasal organ where they have been shown to respond to their known ligands, FMLP and lipoxin A4. Isolated mouse Olfactory bulb neurons also respond to a range of other fpr agonists. These results suggest that the cited receptors function to allow the olfactory-based detection of various contaminated compounds such as spoiled food and/or their many inflammation-regulating and other agonists in bodily secretions.[27]
Gene knockout studies
The large number of mouse compared to human FPR receptors makes it difficult to extrapolate human FPR1 functions based on genetic (e.g. gene knockout or forced overexpression) or other experimental manipulations of FPR receptors in mice. In any event, targeted disruption of the Fpr1 gene reduced the ability of mice to survive intravenous injection of the bacterial pathogen, listeria monocytogenes;[28] disruption of the Fpr2 gene in mice produce a similar effect while disruption of both genes further lowered the survival of mice to the listeria challenge.[29] The effect of these gene knockouts appeared due to faulty leukocyte function and other causes leading to a breakdown in the innate immune response. The functions of the human FPR1 receptor may be equivalent to the overlapping functions of the mouse Fpr1 and Fpr2 functions and therefore be critical in the defense against at least certain bacteria. Targeted disruption of FPR-rs1 produced a 33% reduction in the lifetime of mice; there was no specific pathology associated with this reduction.[26]
Other species
FPR receptors are widely distributed throughout mammalian species with the FPR1, FPR2, and FPR3 paralogs, based on phylogenetic analysis, originating from a common ancestor, early duplication of FPR1, and FPR2/FPR3 splitting with FPR3 originating from the latest duplication event near the origin of primates.[30] Rabbits express an ortholog of FPR1 (78% amino acid sequence identity) with high binding affinity for FMLP; rats express an ortholog of FPR2 (74% amino acid sequence identity) with high affinity for lipoxin A4.[6]
Cellular and tissue distribution
FPR1 is widely expressed by circulating blood neutrophils, eosinophils, basophils, monocytes, and platelets; tissue-bound macrophages, fibroblasts, and immature dendritic cells; vascular endothelial and smooth muscle cells; various types of epithelial cells, liver hepatocytes, neural tissue glial cells, astrocytes and malignant neuroblastoma cells; skin keratinocytes; and virtually all types of multicellular tissues.[6][31][32][33]
See also
References
- GRCh38: Ensembl release 89: ENSG00000171051 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000045551 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Entrez Gene: Formyl peptide receptor 1".
- Migeotte I, Communi D, Parmentier M (Dec 2006). "Formyl peptide receptors: a promiscuous subfamily of G protein-coupled receptors controlling immune responses". Cytokine & Growth Factor Reviews. 17 (6): 501–19. doi:10.1016/j.cytogfr.2006.09.009. PMID 17084101.
- Ye RD, Boulay F, Wang JM, Dahlgren C, Gerard C, Parmentier M, Serhan CN, Murphy PM (Jun 2009). "International Union of Basic and Clinical Pharmacology. LXXIII. Nomenclature for the formyl peptide receptor (FPR) family". Pharmacological Reviews. 61 (2): 119–61. doi:10.1124/pr.109.001578. PMC 2745437. PMID 19498085.
- Boulay F, Tardif M, Brouchon L, Vignais P (May 1990). "Synthesis and use of a novel N-formyl peptide derivative to isolate a human N-formyl peptide receptor cDNA". Biochemical and Biophysical Research Communications. 168 (3): 1103–9. doi:10.1016/0006-291X(90)91143-G. PMID 2161213.
- Zhang Y, Syed R, Uygar C, Pallos D, Gorry MC, Firatli E, Cortelli JR, VanDyke TE, Hart PS, Feingold E, Hart TC (Jan 2003). "Evaluation of human leukocyte N-formylpeptide receptor (FPR1) SNPs in aggressive periodontitis patients". Genes and Immunity. 4 (1): 22–9. doi:10.1038/sj.gene.6363900. PMID 12595898.
- Polakis PG, Uhing RJ, Snyderman R (Apr 1988). "The formylpeptide chemoattractant receptor copurifies with a GTP-binding protein containing a distinct 40-kDa pertussis toxin substrate". The Journal of Biological Chemistry. 263 (10): 4969–76. PMID 2832415.
- Boulay F, Tardif M, Brouchon L, Vignais P (May 1990). "Synthesis and use of a novel N-formyl peptide derivative to isolate a human N-formyl peptide receptor cDNA". Biochemical and Biophysical Research Communications. 168 (3): 1103–9. doi:10.1016/0006-291x(90)91143-g. PMID 2161213.
- Boulay F, Tardif M, Brouchon L, Vignais P (Dec 1990). "The human N-formylpeptide receptor. Characterization of two cDNA isolates and evidence for a new subfamily of G-protein-coupled receptors". Biochemistry. 29 (50): 11123–33. doi:10.1021/bi00502a016. PMID 2176894.
- Murphy PM, Gallin EK, Tiffany HL, Malech HL (Feb 1990). "The formyl peptide chemoattractant receptor is encoded by a 2 kilobase messenger RNA. Expression in Xenopus oocytes". FEBS Letters. 261 (2): 353–7. doi:10.1016/0014-5793(90)80590-f. PMID 1690150.
- Coats WD, Navarro J (Apr 1990). "Functional reconstitution of fMet-Leu-Phe receptor in Xenopus laevis oocytes". The Journal of Biological Chemistry. 265 (11): 5964–6. PMID 2156834.
- Perez HD, Holmes R, Kelly E, McClary J, Chou Q, Andrews WH (Nov 1992). "Cloning of the gene coding for a human receptor for formyl peptides. Characterization of a promoter region and evidence for polymorphic expression". Biochemistry. 31 (46): 11595–9. doi:10.1021/bi00161a044. PMID 1445895.
- Thomas KM, Pyun HY, Navarro J (Nov 1990). "Molecular cloning of the fMet-Leu-Phe receptor from neutrophils". The Journal of Biological Chemistry. 265 (33): 20061–4. PMID 1700779.
- Bao L, Gerard NP, Eddy RL, Shows TB, Gerard C (Jun 1992). "Mapping of genes for the human C5a receptor (C5AR), human FMLP receptor (FPR), and two FMLP receptor homologue orphan receptors (FPRH1, FPRH2) to chromosome 19". Genomics. 13 (2): 437–40. doi:10.1016/0888-7543(92)90265-t. PMID 1612600.
- Murphy PM, Ozçelik T, Kenney RT, Tiffany HL, McDermott D, Francke U (Apr 1992). "A structural homologue of the N-formyl peptide receptor. Characterization and chromosome mapping of a peptide chemoattractant receptor family". The Journal of Biological Chemistry. 267 (11): 7637–43. PMID 1373134.
- Ye RD, Cavanagh SL, Quehenberger O, Prossnitz ER, Cochrane CG (Apr 1992). "Isolation of a cDNA that encodes a novel granulocyte N-formyl peptide receptor". Biochemical and Biophysical Research Communications. 184 (2): 582–9. doi:10.1016/0006-291x(92)90629-y. PMID 1374236.
- Li R, Coulthard LG, Wu MC, Taylor SM, Woodruff TM (Mar 2013). "C5L2: a controversial receptor of complement anaphylatoxin, C5a". FASEB Journal. 27 (3): 855–64. doi:10.1096/fj.12-220509. PMID 23239822.
- Klos A, Wende E, Wareham KJ, Monk PN (Jan 2013). "International Union of Basic and Clinical Pharmacology. [corrected]. LXXXVII. Complement peptide C5a, C4a, and C3a receptors". Pharmacological Reviews. 65 (1): 500–43. doi:10.1124/pr.111.005223. PMID 23383423.
- Paral D, Sohns B, Crass T, Grove M, Köhl J, Klos A, Bautsch W (Aug 1998). "Genomic organization of the human C3a receptor". European Journal of Immunology. 28 (8): 2417–23. doi:10.1002/(SICI)1521-4141(199808)28:08<2417::AID-IMMU2417>3.0.CO;2-B. PMID 9710219.
- He HQ, Liao D, Wang ZG, Wang ZL, Zhou HC, Wang MW, Ye RD (Feb 2013). "Functional characterization of three mouse formyl peptide receptors". Molecular Pharmacology. 83 (2): 389–98. doi:10.1124/mol.112.081315. PMC 4170117. PMID 23160941.
- Takano T, Fiore S, Maddox JF, Brady HR, Petasis NA, Serhan CN (May 1997). "Aspirin-triggered 15-epi-lipoxin A4 (LXA4) and LXA4 stable analogues are potent inhibitors of acute inflammation: evidence for anti-inflammatory receptors". The Journal of Experimental Medicine. 185 (9): 1693–704. doi:10.1084/jem.185.9.1693. PMC 2196289. PMID 9151906.
- Vaughn MW, Proske RJ, Haviland DL (Sep 2002). "Identification, cloning, and functional characterization of a murine lipoxin A4 receptor homologue gene". Journal of Immunology. 169 (6): 3363–9. doi:10.4049/jimmunol.169.6.3363. PMID 12218158.
- Tiffany HL, Gao JL, Roffe E, Sechler JM, Murphy PM (2011). "Characterization of Fpr-rs8, an atypical member of the mouse formyl peptide receptor gene family". Journal of Innate Immunity. 3 (5): 519–29. doi:10.1159/000327718. PMC 3186714. PMID 21691049.
- Rivière S, Challet L, Fluegge D, Spehr M, Rodriguez I (May 2009). "Formyl peptide receptor-like proteins are a novel family of vomeronasal chemosensors". Nature. 459 (7246): 574–7. Bibcode:2009Natur.459..574R. doi:10.1038/nature08029. PMID 19387439.
- Gao JL, Lee EJ, Murphy PM (Feb 1999). "Impaired antibacterial host defense in mice lacking the N-formylpeptide receptor". The Journal of Experimental Medicine. 189 (4): 657–62. doi:10.1084/jem.189.4.657. PMC 2192926. PMID 9989980.
- Liu M, Chen K, Yoshimura T, Liu Y, Gong W, Wang A, Gao JL, Murphy PM, Wang JM (2012). "Formylpeptide receptors are critical for rapid neutrophil mobilization in host defense against Listeria monocytogenes". Scientific Reports. 2: 786. Bibcode:2012NatSR...2E.786L. doi:10.1038/srep00786. PMC 3493074. PMID 23139859.
- Muto Y, Guindon S, Umemura T, Kőhidai L, Ueda H (Feb 2015). "Adaptive evolution of formyl peptide receptors in mammals". Journal of Molecular Evolution. 80 (2): 130–41. Bibcode:2015JMolE..80..130M. doi:10.1007/s00239-015-9666-z. PMID 25627928.
- de Paulis A, Prevete N, Fiorentino I, Walls AF, Curto M, Petraroli A, Castaldo V, Ceppa P, Fiocca R, Marone G (Jun 2004). "Basophils infiltrate human gastric mucosa at sites of Helicobacter pylori infection, and exhibit chemotaxis in response to H. pylori-derived peptide Hp(2-20)". Journal of Immunology. 172 (12): 7734–43. doi:10.4049/jimmunol.172.12.7734. PMID 15187157.
- Svensson L, Redvall E, Björn C, Karlsson J, Bergin AM, Rabiet MJ, Dahlgren C, Wennerås C (Jul 2007). "House dust mite allergen activates human eosinophils via formyl peptide receptor and formyl peptide receptor-like 1". European Journal of Immunology. 37 (7): 1966–77. doi:10.1002/eji.200636936. PMID 17559171.
- Scanzano A, Schembri L, Rasini E, Luini A, Dallatorre J, Legnaro M, Bombelli R, Congiu T, Cosentino M, Marino F (Feb 2015). "Adrenergic modulation of migration, CD11b and CD18 expression, ROS and interleukin-8 production by human polymorphonuclear leukocytes". Inflammation Research. 64 (2): 127–35. doi:10.1007/s00011-014-0791-8. PMID 25561369.
Further reading
- Graves V, Gabig T, McCarthy L, Strour EF, Leemhuis T, English D (Aug 1992). "Simultaneous mobilization of Mac-1 (CD11b/CD18) and formyl peptide chemoattractant receptors in human neutrophils". Blood. 80 (3): 776–87. doi:10.1182/blood.V80.3.776.776. PMID 1322204.
- Murphy PM, Ozçelik T, Kenney RT, Tiffany HL, McDermott D, Francke U (Apr 1992). "A structural homologue of the N-formyl peptide receptor. Characterization and chromosome mapping of a peptide chemoattractant receptor family". The Journal of Biological Chemistry. 267 (11): 7637–43. PMID 1373134.
- Perez HD, Holmes R, Kelly E, McClary J, Chou Q, Andrews WH (Nov 1992). "Cloning of the gene coding for a human receptor for formyl peptides. Characterization of a promoter region and evidence for polymorphic expression". Biochemistry. 31 (46): 11595–9. doi:10.1021/bi00161a044. PMID 1445895.
- Bao L, Gerard NP, Eddy RL, Shows TB, Gerard C (Jun 1992). "Mapping of genes for the human C5a receptor (C5AR), human FMLP receptor (FPR), and two FMLP receptor homologue orphan receptors (FPRH1, FPRH2) to chromosome 19". Genomics. 13 (2): 437–40. doi:10.1016/0888-7543(92)90265-T. PMID 1612600.
- Murphy PM, McDermott D (Jul 1991). "Functional expression of the human formyl peptide receptor in Xenopus oocytes requires a complementary human factor". The Journal of Biological Chemistry. 266 (19): 12560–7. PMID 1712023.
- Boulay F, Tardif M, Brouchon L, Vignais P (Dec 1990). "The human N-formylpeptide receptor. Characterization of two cDNA isolates and evidence for a new subfamily of G-protein-coupled receptors". Biochemistry. 29 (50): 11123–33. doi:10.1021/bi00502a016. PMID 2176894.
- Wahl SM, Allen JB, Gartner S, Orenstein JM, Popovic M, Chenoweth DE, Arthur LO, Farrar WL, Wahl LM (May 1989). "HIV-1 and its envelope glycoprotein down-regulate chemotactic ligand receptors and chemotactic function of peripheral blood monocytes". Journal of Immunology. 142 (10): 3553–9. PMID 2541200.
- Jesaitis AJ, Naemura JR, Painter RG, Sklar LA, Cochrane CG (Dec 1982). "Intracellular localization of N-formyl chemotactic receptor and Mg2+ dependent ATPase in human granulocytes". Biochimica et Biophysica Acta. 719 (3): 556–68. doi:10.1016/0304-4165(82)90246-x. PMID 6129903.
- Prossnitz ER, Kim CM, Benovic JL, Ye RD (Jan 1995). "Phosphorylation of the N-formyl peptide receptor carboxyl terminus by the G protein-coupled receptor kinase, GRK2". The Journal of Biological Chemistry. 270 (3): 1130–7. doi:10.1074/jbc.270.3.1130. PMID 7836371.
- Klotz KN, Jesaitis AJ (Sep 1994). "Physical coupling of N-formyl peptide chemoattractant receptors to G protein is unaffected by desensitization". Biochemical Pharmacology. 48 (6): 1297–300. doi:10.1016/0006-2952(94)90168-6. PMID 7945424.
- Bommakanti RK, Dratz EA, Siemsen DW, Jesaitis AJ (Nov 1994). "Characterization of complex formation between Gi2 and octyl glucoside solubilized neutrophil N-formyl peptide chemoattractant receptor by sedimentation velocity". Biochimica et Biophysica Acta (BBA) - Protein Structure and Molecular Enzymology. 1209 (1): 69–76. doi:10.1016/0167-4838(94)90138-4. PMID 7947984.
- Murphy PM, Tiffany HL, McDermott D, Ahuja SK (Nov 1993). "Sequence and organization of the human N-formyl peptide receptor-encoding gene". Gene. 133 (2): 285–90. doi:10.1016/0378-1119(93)90653-K. PMID 8224916.
- Jesaitis AJ, Erickson RW, Klotz KN, Bommakanti RK, Siemsen DW (Nov 1993). "Functional molecular complexes of human N-formyl chemoattractant receptors and actin". Journal of Immunology. 151 (10): 5653–65. PMID 8228254.
- Särndahl E, Bokoch GM, Boulay F, Stendahl O, Andersson T (Jun 1996). "Direct or C5a-induced activation of heterotrimeric Gi2 proteins in human neutrophils is associated with interaction between formyl peptide receptors and the cytoskeleton". The Journal of Biological Chemistry. 271 (25): 15267–71. doi:10.1074/jbc.271.25.15267. PMID 8663057.
- Maestes DC, Potter RM, Prossnitz ER (Oct 1999). "Differential phosphorylation paradigms dictate desensitization and internalization of the N-formyl peptide receptor". The Journal of Biological Chemistry. 274 (42): 29791–5. doi:10.1074/jbc.274.42.29791. PMID 10514456.
- Liang TS, Wang JM, Murphy PM, Gao JL (Apr 2000). "Serum amyloid A is a chemotactic agonist at FPR2, a low-affinity N-formylpeptide receptor on mouse neutrophils". Biochemical and Biophysical Research Communications. 270 (2): 331–5. doi:10.1006/bbrc.2000.2416. PMID 10753626.
- Luu NT, Rainger GE, Nash GB (Jun 2000). "Differential ability of exogenous chemotactic agents to disrupt transendothelial migration of flowing neutrophils". Journal of Immunology. 164 (11): 5961–9. doi:10.4049/jimmunol.164.11.5961. PMID 10820279.
- Bennett TA, Maestas DC, Prossnitz ER (Aug 2000). "Arrestin binding to the G protein-coupled N-formyl peptide receptor is regulated by the conserved "DRY" sequence". The Journal of Biological Chemistry. 275 (32): 24590–4. doi:10.1074/jbc.C000314200. PMID 10823817.
- Ayala JM, Goyal S, Liverton NJ, Claremon DA, O'Keefe SJ, Hanlon WA (Jun 2000). "Serum-induced monocyte differentiation and monocyte chemotaxis are regulated by the p38 MAP kinase signal transduction pathway". Journal of Leukocyte Biology. 67 (6): 869–75. doi:10.1002/jlb.67.6.869. PMID 10857861.
This article incorporates text from the United States National Library of Medicine, which is in the public domain.