CHEK2
CHEK2 (Checkpoint kinase 2) is a tumor suppressor gene that encodes the protein CHK2, a serine-threonine kinase. CHK2 is involved in DNA repair, cell cycle arrest or apoptosis in response to DNA damage. Mutations to the CHEK2 gene have been linked to a wide range of cancers.[5]


Gene location
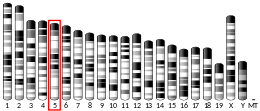
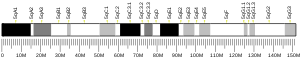
The CHEK2 gene is located on the long (q) arm of chromosome 22 at position 12.1. Its location on chromosome 22 stretches from base pair 28,687,742 to base pair 28,741,904.[5]
Protein structure
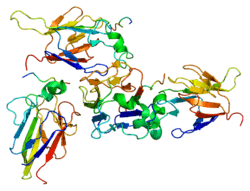
The CHK2 protein encoded by the CHEK2 gene is a serine threonine kinase. The protein consists of 543 amino acids and the following domains:
- N-terminal SQ/TQ cluster doman (SCD)
- Central forkhead-associated (FHA) domain
- C-terminal serine/threonine kinase domain (KD)
The SCD domain contains multiple SQ/TQ motifs that serve as sites for phosphorylation in response to DNA damage. The most notable and frequently phosphorylated site being Thr68.[6]
CHK2 appears as a monomer in its inactive state. However, in the event of DNA damage SCD phosphorylation causes CHK2 dimerization. The phosphorylated Thr68 (located on the SCD) interacts with the FHA domain to form the dimer. After the protein dimerizes the KD is activated via autophosphorylation. Once the KD is activated the CHK2 dimer dissociates.[6]
Function and mechanism
The CHEK2 gene encodes for checkpoint kinase 2 (CHK2), a protein that acts a tumor suppressor. CHK2 regulates cell division, and has the ability to prevent cells from dividing too rapidly or in an uncontrolled manner.[5]
When DNA undergoes a double-strand break, CHK2 is activated. Specifically, DNA damage-activated phosphatidylinositol kinase family protein (PIKK) ATM phosphorylates site Thr68 and activates CHK2.[6] Once activated, CHK2 phosphorylates downstream targets including CDC25 phosphatases, responsible for dephosphorylating and activating the cyclin-dependent kinases (CDKs). Thus, CHK2’s inhibition of the CDC25 phosphatases prevents entry of the cell into mitosis. Furthermore, the CHK2 protein interacts with several other proteins including p53 (p53). Stabilization of p53 by CHK2 leads to cell cycle arrest in phase G1. Furthermore, CHK2 is known to phosphorylate the cell-cycle transcription factor E2F1 and the promyelocytic leukemia protein (PML) involved in apoptosis (programmed cell death).[6]
Association with cancer
The CHK2 protein plays a critical role in the DNA damage checkpoint. Thus, mutations to the CHEK2 gene have been labeled as causes to a wide range of cancers.
In 1999, genetic variations of CHEK2 were found to correspond to inherited cancer susceptibility.[7]
Bell et al. (1999) discovered three CHEK2 germline mutations among four Li–Fraumeni syndrome (LFS) and 18 Li–Fraumeni-like (LFL) families. Since the time of this discovery, two of the three variants (a deletion in the kinase domain in exon 10 and a missense mutation in the FHA domain in exon 3) have been linked to inherited susceptibility to breast as well as other cancers.[8]
Beyond initial speculations, screening of LFS and LFL patients has revealed no or very rare individual missense variants in the CHEK2 gene. Additionally, the deletion in the kinase domain on exon 10 has been found rare among LFS/LFL patients. The evidence from these studies has suggests that CHEK2 is not a predisposition gene to Li–Fraumeni syndrome.[8]
Breast cancer
Inherited mutations in the CHEK2 gene have been linked to certain cases of breast cancer. Most notably, the deletion of a single DNA nucleotide at position 1100 in exon 10 (1100delC) produces a nonfunctional version of the CHK2 protein, truncated at the kinase domain. The loss of normal CHK2 protein function leads to unregulated cell division, accumulated damage to DNA and in many cases, tumor development.[5] The CHEK2*1100del mutation is most commonly seen in individuals of Eastern and Northern European descent. Within these populations the CHEK2*1100delC mutation is seen in 1 out of 100 to 1 out of 200 individuals. However, in North America the frequency drops to 1 out of 333 to 1 out of 500. The mutation is almost absent in Spain and India.[9] Studies show that a CHEK2 1100delC corresponds to a two-fold increased risk of breast cancer and a 10-fold increased risk of breast cancer in males.[10]
A CHEK2 mutation known as the I157T variant to the FHA domain in exon 3 has also been linked to breast cancer but at a lower risk than the CHEK2*1100delC mutation. The estimated fraction of breast cancer attributed to this variant is reported to be around 1.2% in the US.[8]
Two more CHEK2 gene mutations, CHEK2*S428F, an amino-acid substitution to the kinase domain in exon 11 and CHEK2*P85L, an amino-acid substitution in the N-terminal region (exon 1) have been found in the Ashkenazi Jewish population.[9] Suggestion of a Hispanic founder mutation has also been described.[11]
Other cancers
Mutations to CHEK2 have been found in hereditary and nonhereditary cases of cancer. Studies link the mutation to cases of prostate, lung, colon, kidney, and thyroid cancers. Links have also been drawn to certain brain tumors and osteosarcoma.[5]
Unlike BRCA1 and BRCA2 mutations, CHEK2 mutations do not appear to cause an elevated risk for ovarian cancer.[10]
Meiosis
CHEK2 regulates cell cycle progression and spindle assembly during mouse oocyte maturation and early embryo development.[12] Although CHEK2 is a down stream effector of the ATM kinase that responds primarily to double-strand breaks it can also be activated by ATR (ataxia-telangiectasia and Rad3 related) kinase that responds primarily to single-strand breaks. In mice, CHEK2 is essential for DNA damage surveillance in female meiosis. The response of oocytes to DNA double-strand break damage involves a pathway hierarchy in which ATR kinase signals to CHEK2 which then activates p53 and p63 proteins.[13]
In the fruit fly Drosophila, irradiation of germ line cells generates double-strand breaks that result in cell cycle arrest and apoptosis. The Drosophila CHEK2 ortholog mnk and the p53 ortholog dp53 are required for much of the cell death observed in early oogenesis when oocyte selection and meiotic recombination occur.[14]
Interactions
CHEK2 has been shown to interact with:
References
- GRCh38: Ensembl release 89: ENSG00000183765 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000029521 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "CHEK2". Genetics Home Reference. August 2007.
- Cai Z, Chehab NH, Pavletich NP (Sep 2009). "Structure and activation mechanism of the CHK2 DNA damage checkpoint kinase". Molecular Cell. 35 (6): 818–29. doi:10.1016/j.molcel.2009.09.007. PMID 19782031.
- Bell DW, Varley JM, Szydlo TE, Kang DH, Wahrer DC, Shannon KE, Lubratovich M, Verselis SJ, Isselbacher KJ, Fraumeni JF, Birch JM, Li FP, Garber JE, Haber DA (Dec 1999). "Heterozygous germ line hCHK2 mutations in Li-Fraumeni syndrome". Science. 286 (5449): 2528–31. doi:10.1126/science.286.5449.2528. PMID 10617473.
- Nevanlinna H, Bartek J (Sep 2006). "The CHEK2 gene and inherited breast cancer susceptibility". Oncogene. 25 (43): 5912–9. doi:10.1038/sj.onc.1209877. PMID 16998506.
- Offit K, Garber JE (Feb 2008). "Time to check CHEK2 in families with breast cancer?". Journal of Clinical Oncology. 26 (4): 519–20. doi:10.1200/JCO.2007.13.8503. PMID 18172189.
- Meijers-Heijboer H, van den Ouweland A, Klijn J, Wasielewski M, de Snoo A, Oldenburg R, Hollestelle A, Houben M, Crepin E, van Veghel-Plandsoen M, Elstrodt F, van Duijn C, Bartels C, Meijers C, Schutte M, McGuffog L, Thompson D, Easton D, Sodha N, Seal S, Barfoot R, Mangion J, Chang-Claude J, Eccles D, Eeles R, Evans DG, Houlston R, Murday V, Narod S, Peretz T, Peto J, Phelan C, Zhang HX, Szabo C, Devilee P, Goldgar D, Futreal PA, Nathanson KL, Weber B, Rahman N, Stratton MR (May 2002). "Low-penetrance susceptibility to breast cancer due to CHEK2(*)1100delC in noncarriers of BRCA1 or BRCA2 mutations". Nature Genetics. 31 (1): 55–9. doi:10.1038/ng879. PMID 11967536.
- Weitzel, Jeffrey N.; Neuhausen, Susan L.; Adamson, Aaron; Tao, Shu; Ricker, Charité; Maoz, Asaf; Rosenblatt, Margalit; Nehoray, Bita; Sand, Sharon (2019-06-17). "Pathogenic and likely pathogenic variants in PALB2, CHEK2, and other known breast cancer susceptibility genes among 1054 BRCA-negative Hispanics with breast cancer". Cancer. 125 (16): 2829–2836. doi:10.1002/cncr.32083. ISSN 1097-0142. PMID 31206626.
- Dai XX, Duan X, Liu HL, Cui XS, Kim NH, Sun SC (2014). "Chk2 regulates cell cycle progression during mouse oocyte maturation and early embryo development". Mol. Cells. 37 (2): 126–32. doi:10.14348/molcells.2014.2259. PMC 3935625. PMID 24598997.
- Bolcun-Filas E, Rinaldi VD, White ME, Schimenti JC (2014). "Reversal of female infertility by Chk2 ablation reveals the oocyte DNA damage checkpoint pathway". Science. 343 (6170): 533–6. Bibcode:2014Sci...343..533B. doi:10.1126/science.1247671. PMC 4048839. PMID 24482479.
- Shim HJ, Lee EM, Nguyen LD, Shim J, Song YH (2014). "High-dose irradiation induces cell cycle arrest, apoptosis, and developmental defects during Drosophila oogenesis". PLOS ONE. 9 (2): e89009. Bibcode:2014PLoSO...989009S. doi:10.1371/journal.pone.0089009. PMC 3923870. PMID 24551207.
- Lee JS, Collins KM, Brown AL, Lee CH, Chung JH (Mar 2000). "hCds1-mediated phosphorylation of BRCA1 regulates the DNA damage response". Nature. 404 (6774): 201–4. Bibcode:2000Natur.404..201L. doi:10.1038/35004614. PMID 10724175.
- Chabalier-Taste C, Racca C, Dozier C, Larminat F (Dec 2008). "BRCA1 is regulated by Chk2 in response to spindle damage". Biochimica et Biophysica Acta (BBA) - Molecular Cell Research. 1783 (12): 2223–33. doi:10.1016/j.bbamcr.2008.08.006. PMID 18804494.
- Matsuoka S, Ballif BA, Smogorzewska A, McDonald ER, Hurov KE, Luo J, Bakalarski CE, Zhao Z, Solimini N, Lerenthal Y, Shiloh Y, Gygi SP, Elledge SJ (May 2007). "ATM and ATR substrate analysis reveals extensive protein networks responsive to DNA damage". Science. 316 (5828): 1160–6. Bibcode:2007Sci...316.1160M. doi:10.1126/science.1140321. PMID 17525332. S2CID 16648052.
- Lou Z, Minter-Dykhouse K, Wu X, Chen J (Feb 2003). "MDC1 is coupled to activated CHK2 in mammalian DNA damage response pathways". Nature. 421 (6926): 957–61. Bibcode:2003Natur.421..957L. doi:10.1038/nature01447. PMID 12607004.
- Adamson AW, Beardsley DI, Kim WJ, Gao Y, Baskaran R, Brown KD (Mar 2005). "Methylator-induced, mismatch repair-dependent G2 arrest is activated through Chk1 and Chk2". Molecular Biology of the Cell. 16 (3): 1513–26. doi:10.1091/mbc.E04-02-0089. PMC 551512. PMID 15647386.
- Brown KD, Rathi A, Kamath R, Beardsley DI, Zhan Q, Mannino JL, Baskaran R (Jan 2003). "The mismatch repair system is required for S-phase checkpoint activation". Nature Genetics. 33 (1): 80–4. doi:10.1038/ng1052. PMID 12447371.
- Chen XB, Melchionna R, Denis CM, Gaillard PH, Blasina A, Van de Weyer I, Boddy MN, Russell P, Vialard J, McGowan CH (Nov 2001). "Human Mus81-associated endonuclease cleaves Holliday junctions in vitro". Molecular Cell. 8 (5): 1117–27. doi:10.1016/s1097-2765(01)00375-6. PMID 11741546.
- Tsvetkov L, Xu X, Li J, Stern DF (Mar 2003). "Polo-like kinase 1 and Chk2 interact and co-localize to centrosomes and the midbody". The Journal of Biological Chemistry. 278 (10): 8468–75. doi:10.1074/jbc.M211202200. PMID 12493754.
- Bahassi el M, Conn CW, Myer DL, Hennigan RF, McGowan CH, Sanchez Y, Stambrook PJ (Sep 2002). "Mammalian Polo-like kinase 3 (Plk3) is a multifunctional protein involved in stress response pathways". Oncogene. 21 (43): 6633–40. doi:10.1038/sj.onc.1205850. PMID 12242661.
Further reading
- Caspari T (Apr 2000). "How to activate p53". Current Biology. 10 (8): R315–7. doi:10.1016/S0960-9822(00)00439-5. PMID 10801407.
- McGowan CH (Jun 2002). "Checking in on Cds1 (Chk2): A checkpoint kinase and tumor suppressor". BioEssays. 24 (6): 502–11. doi:10.1002/bies.10101. PMID 12111733.
- Honrado E, Osorio A, Palacios J, Benitez J (Sep 2006). "Pathology and gene expression of hereditary breast tumors associated with BRCA1, BRCA2 and CHEK2 gene mutations". Oncogene. 25 (43): 5837–45. doi:10.1038/sj.onc.1209875. PMID 16998498.
- Nevanlinna H, Bartek J (Sep 2006). "The CHEK2 gene and inherited breast cancer susceptibility". Oncogene. 25 (43): 5912–9. doi:10.1038/sj.onc.1209877. PMID 16998506.
- Peng CY, Graves PR, Thoma RS, Wu Z, Shaw AS, Piwnica-Worms H (Sep 1997). "Mitotic and G2 checkpoint control: regulation of 14-3-3 protein binding by phosphorylation of Cdc25C on serine-216". Science. 277 (5331): 1501–5. doi:10.1126/science.277.5331.1501. PMID 9278512.
- Lykidis A, Jackson PD, Rock CO, Jackowski S (Dec 1997). "The role of CDP-diacylglycerol synthetase and phosphatidylinositol synthase activity levels in the regulation of cellular phosphatidylinositol content". The Journal of Biological Chemistry. 272 (52): 33402–9. doi:10.1074/jbc.272.52.33402. PMID 9407135.
- Lindsay HD, Griffiths DJ, Edwards RJ, Christensen PU, Murray JM, Osman F, Walworth N, Carr AM (Feb 1998). "S-phase-specific activation of Cds1 kinase defines a subpathway of the checkpoint response in Schizosaccharomyces pombe". Genes & Development. 12 (3): 382–95. doi:10.1101/gad.12.3.382. PMC 316487. PMID 9450932.
- Matsuoka S, Huang M, Elledge SJ (Dec 1998). "Linkage of ATM to cell cycle regulation by the Chk2 protein kinase". Science. 282 (5395): 1893–7. Bibcode:1998Sci...282.1893M. doi:10.1126/science.282.5395.1893. PMID 9836640.
- Martinho RG, Lindsay HD, Flaggs G, DeMaggio AJ, Hoekstra MF, Carr AM, Bentley NJ (Dec 1998). "Analysis of Rad3 and Chk1 protein kinases defines different checkpoint responses". The EMBO Journal. 17 (24): 7239–49. doi:10.1093/emboj/17.24.7239. PMC 1171070. PMID 9857181.
- Blasina A, de Weyer IV, Laus MC, Luyten WH, Parker AE, McGowan CH (Jan 1999). "A human homologue of the checkpoint kinase Cds1 directly inhibits Cdc25 phosphatase". Current Biology. 9 (1): 1–10. doi:10.1016/S0960-9822(99)80041-4. PMID 9889122.
- Brown AL, Lee CH, Schwarz JK, Mitiku N, Piwnica-Worms H, Chung JH (Mar 1999). "A human Cds1-related kinase that functions downstream of ATM protein in the cellular response to DNA damage". Proceedings of the National Academy of Sciences of the United States of America. 96 (7): 3745–50. Bibcode:1999PNAS...96.3745B. doi:10.1073/pnas.96.7.3745. PMC 22365. PMID 10097108.
- Chaturvedi P, Eng WK, Zhu Y, Mattern MR, Mishra R, Hurle MR, Zhang X, Annan RS, Lu Q, Faucette LF, Scott GF, Li X, Carr SA, Johnson RK, Winkler JD, Zhou BB (Jul 1999). "Mammalian Chk2 is a downstream effector of the ATM-dependent DNA damage checkpoint pathway". Oncogene. 18 (28): 4047–54. doi:10.1038/sj.onc.1202925. PMID 10435585.
- Ouyang B, Li W, Pan H, Meadows J, Hoffmann I, Dai W (Oct 1999). "The physical association and phosphorylation of Cdc25C protein phosphatase by Prk". Oncogene. 18 (44): 6029–36. doi:10.1038/sj.onc.1202983. PMID 10557092.
- Dunham I, Shimizu N, Roe BA, Chissoe S, Hunt AR, Collins JE, Bruskiewich R, Beare DM, Clamp M, Smink LJ, Ainscough R, Almeida JP, Babbage A, Bagguley C, Bailey J, Barlow K, Bates KN, Beasley O, Bird CP, Blakey S, Bridgeman AM, Buck D, Burgess J, Burrill WD, O'Brien KP (Dec 1999). "The DNA sequence of human chromosome 22". Nature. 402 (6761): 489–95. Bibcode:1999Natur.402..489D. doi:10.1038/990031. PMID 10591208.
- Bell DW, Varley JM, Szydlo TE, Kang DH, Wahrer DC, Shannon KE, Lubratovich M, Verselis SJ, Isselbacher KJ, Fraumeni JF, Birch JM, Li FP, Garber JE, Haber DA (Dec 1999). "Heterozygous germ line hCHK2 mutations in Li-Fraumeni syndrome". Science. 286 (5449): 2528–31. doi:10.1126/science.286.5449.2528. PMID 10617473.
- Chehab NH, Malikzay A, Appel M, Halazonetis TD (Feb 2000). "Chk2/hCds1 functions as a DNA damage checkpoint in G(1) by stabilizing p53". Genes & Development. 14 (3): 278–88. doi:10.1101/gad.14.3.278 (inactive 2020-05-25). PMC 316357. PMID 10673500.
- Hirao A, Kong YY, Matsuoka S, Wakeham A, Ruland J, Yoshida H, Liu D, Elledge SJ, Mak TW (Mar 2000). "DNA damage-induced activation of p53 by the checkpoint kinase Chk2". Science. 287 (5459): 1824–7. Bibcode:2000Sci...287.1824H. doi:10.1126/science.287.5459.1824. PMID 10710310.
- Lee JS, Collins KM, Brown AL, Lee CH, Chung JH (Mar 2000). "hCds1-mediated phosphorylation of BRCA1 regulates the DNA damage response". Nature. 404 (6774): 201–4. Bibcode:2000Natur.404..201L. doi:10.1038/35004614. PMID 10724175.
- Zhou BB, Chaturvedi P, Spring K, Scott SP, Johanson RA, Mishra R, Mattern MR, Winkler JD, Khanna KK (Apr 2000). "Caffeine abolishes the mammalian G(2)/M DNA damage checkpoint by inhibiting ataxia-telangiectasia-mutated kinase activity" (PDF). The Journal of Biological Chemistry. 275 (14): 10342–8. doi:10.1074/jbc.275.14.10342. PMID 10744722.
External links
- Educational resources
- Gene Reviews
- Gene Tests
- CHEK2+protein,+human at the US National Library of Medicine Medical Subject Headings (MeSH)
- Human CDS1 genome location and CDS1 gene details page in the UCSC Genome Browser.
- Human CHEK2 genome location and CHEK2 gene details page in the UCSC Genome Browser.
PDB gallery | |
|---|---|
|
This article incorporates text from the United States National Library of Medicine, which is in the public domain.