HSPA1L
Heat shock 70 kDa protein 1L is a protein that in humans is encoded by the HSPA1L gene on chromosome 6.[5][6][7] As a member of the heat shock protein 70 (Hsp70) family and a chaperone protein, it facilitates the proper folding of newly translated and misfolded proteins, as well as stabilize or degrade mutant proteins.[7][8] Its functions contribute to biological processes including signal transduction, apoptosis, protein homeostasis, and cell growth and differentiation.[8][9] It has been associated with an extensive number of cancers, neurodegenerative diseases, cell senescence and aging, and Graft-versus-host disease.[8][9][10]
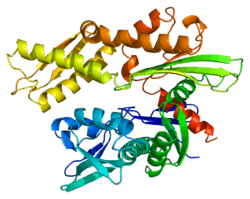



Structure
This gene encodes a 70kDa heat shock protein and is located in the major histocompatibility complex class III region, in a cluster with two closely related genes which also encode isoforms of the 70kDa heat shock protein.[7] The amino acid sequence of the encoded protein shares a 90% homology to the isoforms HSPA1A and HSPA1B.[11] As a Hsp70 protein, it has a C-terminal protein substrate-binding domain and an N-terminal ATP-binding domain.[12][13][14][15] The substrate-binding domain consists of two subdomains, a two-layered β-sandwich subdomain (SBDβ) and an α-helical subdomain (SBDα), which are connected by the loop Lα,β. SBDβ contains the peptide binding pocket while SBDα serves as a lid to cover the substrate binding cleft. The ATP binding domain consists of four subdomains split into two lobes by a central ATP/ADP binding pocket.[14] The two terminal domains are linked together by a conserved region referred to as loop LL,1, which is critical for allosteric regulation. The unstructured region at the very end of the C-terminal is believed to be the docking site for co-chaperones.[14][15]
Since a cDNA clone of this gene contains a 119 bp-region in the 5' UTR, it is likely that HSPA1L contains one or more introns in its own 5' UTR.[11]
Function
In general, HSPA1L is widely distributed across tissues at low abundances, but in particular, it is constitutively and abundantly expressed in the testis.[15][16]
Along with other heat shock proteins, this protein stabilizes existing proteins against aggregation and mediates the folding of newly translated proteins in the cytosol and in organelles.[8][9] In order to properly fold non-native proteins, this protein interacts with the hydrophobic peptide segments of proteins in an ATP-controlled fashion. Though the exact mechanism still remains unclear, there are at least two alternative modes of action: kinetic partitioning and local unfolding. In kinetic partitioning, Hsp70s repetitively bind and release substrates in cycles that maintain low concentrations of free substrate. This effectively prevents aggregation while allowing free molecules to fold to the native state. In local unfolding, the binding and release cycles induce localized unfolding in the substrate, which helps to overcome kinetic barriers for folding to the native state. Ultimately, its role in protein folding contributes to its function in signal transduction, apoptosis, protein homeostasis, and cell growth and differentiation.[8][9]
In addition to the process of protein folding, transport and degradation, this Hsp70 member can preserve the function of mutant proteins. Nonetheless, effects of these mutations can still manifest when Hsp70 chaperones are overwhelmed during stress conditions.[8] Furthermore, this protein enhances antigen-specific tumor immunity by facilitating more efficient antigen presentation to cytotoxic T cells.[9] Though it shares close homology to HSPA1A and HSPA1B, it is regulated differently and is not heat-inducible.[11]
Clinical significance
The Hsp70 member proteins are important apoptotic constituents. During a normal embryologic processes, or during cell injury (such as ischemia-reperfusion injury during heart attacks and strokes) or during developments and processes in cancer, an apoptotic cell undergoes structural changes including cell shrinkage, plasma membrane blebbing, nuclear condensation, and fragmentation of the DNA and nucleus. This is followed by fragmentation into apoptotic bodies that are quickly removed by phagocytes, thereby preventing an inflammatory response.[17] It is a mode of cell death defined by characteristic morphological, biochemical and molecular changes. It was first described as a "shrinkage necrosis", and then this term was replaced by apoptosis to emphasize its role opposite mitosis in tissue kinetics. In later stages of apoptosis the entire cell becomes fragmented, forming a number of plasma membrane-bounded apoptotic bodies which contain nuclear and or cytoplasmic elements. The ultrastructural appearance of necrosis is quite different, the main features being mitochondrial swelling, plasma membrane breakdown and cellular disintegration. Apoptosis occurs in many physiological and pathological processes. It plays an important role during embryonal development as programmed cell death and accompanies a variety of normal involutional processes in which it serves as a mechanism to remove "unwanted" cells.
Hsp70 member proteins, including Hsp72, inhibit apoptosis by acting on the caspase-dependent pathway and against apoptosis-inducing agents such as tumor necrosis factor-α (TNFα), staurosporine, and doxorubicin. This role leads to its involvement in many pathological processes, such as oncogenesis, neurodegeneration, and senescence. In particular, overexpression of HSP72 has been linked to the development some cancers, such as hepatocellular carcinoma, gastric cancers, colon cancers, breast cancers, and lung cancers, which led to its use as a prognostic marker for these cancers.[9] Elevated Hsp70 levels in tumor cells may increase malignancy and resistance to therapy by complexing, and hence, stabilizing, oncofetal proteins and products and transporting them into intracellular sites, thereby promoting tumor cell proliferation.[8][9] As a result, tumor vaccine strategies for Hsp70s have been highly successful in animal models and progressed to clinical trials.[9] Alternatively, overexpression of Hsp70 can mitigate the effects of neurodegenerative diseases, such as Alzheimer’s disease, Parkinson’s disease (PD), Huntington’s disease, and spinocerebellar ataxias, and aging and cell senescence, as observed in centenarians subjected to heat shock challenge.[8] HSPA1L may fight against PD by co-regulating the translocation of parkin to damaged mitochondria, thus facilitating their removal.[16]
HSPA1L is also involved in Graft-versus-host disease (GVHD) and has potential to serve as a diagnostic/prognostic biomarker.[10] Polymorphisms in the HSPA1L gene, especially those in the substrate binding domain, have been associated with disease.[15]
Interactions
HSPA1L has been shown to interact with PARK2.[16]
References
- ENSG00000226704, ENSG00000236251, ENSG00000204390, ENSG00000206383 GRCh38: Ensembl release 89: ENSG00000234258, ENSG00000226704, ENSG00000236251, ENSG00000204390, ENSG00000206383 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000007033 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Ito Y, Ando A, Ando H, Ando J, Saijoh Y, Inoko H, Fujimoto H (Aug 1998). "Genomic structure of the spermatid-specific hsp70 homolog gene located in the class III region of the major histocompatibility complex of mouse and man". Journal of Biochemistry. 124 (2): 347–53. doi:10.1093/oxfordjournals.jbchem.a022118. PMID 9685725.
- Ishihara M, Ohno S (Nov 1997). "Genetic influences on sarcoidosis". Eye. 11. 11 (2): 155–61. doi:10.1038/eye.1997.44. PMID 9349405.
- "Entrez Gene: HSPA1L heat shock 70kDa protein 1-like".
- Mayer MP, Bukau B (Mar 2005). "Hsp70 chaperones: cellular functions and molecular mechanism". Cellular and Molecular Life Sciences. 62 (6): 670–684. doi:10.1007/s00018-004-4464-6. PMC 2773841. PMID 15770419.
- Wang X, Wang Q, Lin H, Li S, Sun L, Yang Y (Feb 2013). "HSP72 and gp96 in gastroenterological cancers". Clinica Chimica Acta; International Journal of Clinical Chemistry. 417: 73–9. doi:10.1016/j.cca.2012.12.017. PMID 23266770.
- Atarod S, Turner B, Pearce KF, Ahmed SS, Norden J, Bogunia-Kubik K, Wang XN, Collin M, Dickinson AM (Feb 2015). "Elevated level of HSPA1L mRNA correlates with graft-versus-host disease". Transplant Immunology. 32 (3): 188–94. doi:10.1016/j.trim.2015.02.002. PMID 25680846.
- Ito Y, Ando A, Ando H, Ando J, Saijoh Y, Inoko H, Fujimoto H (Aug 1998). "Genomic structure of the spermatid-specific hsp70 homolog gene located in the class III region of the major histocompatibility complex of mouse and man". Journal of Biochemistry. 124 (2): 347–53. doi:10.1093/oxfordjournals.jbchem.a022118. PMID 9685725.
- Ravagnan L, Gurbuxani S, Susin SA, Maisse C, Daugas E, Zamzami N, Mak T, Jäättelä M, Penninger JM, Garrido C, Kroemer G (September 2001). "Heat-shock protein 70 antagonizes apoptosis-inducing factor". Nat. Cell Biol. 3 (9): 839–43. doi:10.1038/ncb0901-839. PMID 11533664.
- Zhang B, Rong R, Li H, Peng X, Xiong L, Wang Y, Yu X, Mao H (2015). "Heat shock protein 72 suppresses apoptosis by increasing the stability of X-linked inhibitor of apoptosis protein in renal ischemia/reperfusion injury". Mol Med Rep. 11 (3): 1793–9. doi:10.3892/mmr.2014.2939. PMC 4270332. PMID 25394481.
- Zhang P, Leu JI, Murphy ME, George DL, Marmorstein R (2014). "Crystal structure of the stress-inducible human heat shock protein 70 substrate-binding domain in complex with peptide substrate". PLOS ONE. 9 (7): e103518. doi:10.1371/journal.pone.0103518. PMC 4110032. PMID 25058147.
- Wisniewska M, Karlberg T, Lehtiö L, Johansson I, Kotenyova T, Moche M, Schüler H (11 January 2010). "Crystal structures of the ATPase domains of four human Hsp70 isoforms: HSPA1L/Hsp70-hom, HSPA2/Hsp70-2, HSPA6/Hsp70B', and HSPA5/BiP/GRP78". PLOS ONE. 5 (1): e8625. doi:10.1371/journal.pone.0008625. PMC 2803158. PMID 20072699.
- Hasson SA, Kane LA, Yamano K, Huang CH, Sliter DA, Buehler E, Wang C, Heman-Ackah SM, Hessa T, Guha R, Martin SE, Youle RJ (Dec 2013). "High-content genome-wide RNAi screens identify regulators of parkin upstream of mitophagy". Nature. 504 (7479): 291–5. doi:10.1038/nature12748. PMC 5841086. PMID 24270810.
- Kerr JF, Wyllie AH, Currie AR (Aug 1972). "Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics". British Journal of Cancer. 26 (4): 239–57. doi:10.1038/bjc.1972.33. PMC 2008650. PMID 4561027.
Further reading
- Milner CM, Campbell RD (1992). "Polymorphic analysis of the three MHC-linked HSP70 genes". Immunogenetics. 36 (6): 357–62. doi:10.1007/BF00218042. PMID 1356099.
- Milner CM, Campbell RD (1990). "Structure and expression of the three MHC-linked HSP70 genes". Immunogenetics. 32 (4): 242–51. doi:10.1007/BF00187095. PMID 1700760.
- Sargent CA, Dunham I, Trowsdale J, Campbell RD (Mar 1989). "Human major histocompatibility complex contains genes for the major heat shock protein HSP70". Proceedings of the National Academy of Sciences of the United States of America. 86 (6): 1968–72. doi:10.1073/pnas.86.6.1968. PMC 286826. PMID 2538825.
- Goate AM, Cooper DN, Hall C, Leung TK, Solomon E, Lim L (Feb 1987). "Localization of a human heat-shock HSP 70 gene sequence to chromosome 6 and detection of two other loci by somatic-cell hybrid and restriction fragment length polymorphism analysis". Human Genetics. 75 (2): 123–8. doi:10.1007/BF00591072. PMID 2880793.
- Harrison GS, Drabkin HA, Kao FT, Hartz J, Hart IM, Chu EH, Wu BJ, Morimoto RI (Mar 1987). "Chromosomal location of human genes encoding major heat-shock protein HSP70" (PDF). Somatic Cell and Molecular Genetics. 13 (2): 119–30. doi:10.1007/BF01534692. hdl:2027.42/45535. PMID 3470951.
- Voellmy R, Ahmed A, Schiller P, Bromley P, Rungger D (Aug 1985). "Isolation and functional analysis of a human 70,000-dalton heat shock protein gene segment". Proceedings of the National Academy of Sciences of the United States of America. 82 (15): 4949–53. doi:10.1073/pnas.82.15.4949. PMC 390475. PMID 3927293.
- Maruyama K, Sugano S (Jan 1994). "Oligo-capping: a simple method to replace the cap structure of eukaryotic mRNAs with oligoribonucleotides". Gene. 138 (1–2): 171–4. doi:10.1016/0378-1119(94)90802-8. PMID 8125298.
- Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (Oct 1997). "Construction and characterization of a full length-enriched and a 5'-end-enriched cDNA library". Gene. 200 (1–2): 149–56. doi:10.1016/S0378-1119(97)00411-3. PMID 9373149.
- Hartley JL, Temple GF, Brasch MA (Nov 2000). "DNA cloning using in vitro site-specific recombination". Genome Research. 10 (11): 1788–95. doi:10.1101/gr.143000. PMC 310948. PMID 11076863.
- Fourie AM, Peterson PA, Yang Y (Jul 2001). "Characterization and regulation of the major histocompatibility complex-encoded proteins Hsp70-Hom and Hsp70-1/2". Cell Stress & Chaperones. 6 (3): 282–95. doi:10.1379/1466-1268(2001)006<0282:CAROTM>2.0.CO;2. PMC 434410. PMID 11599570.
- Schröder O, Schulte KM, Ostermann P, Röher HD, Ekkernkamp A, Laun RA (Jan 2003). "Heat shock protein 70 genotypes HSPA1B and HSPA1L influence cytokine concentrations and interfere with outcome after major injury". Critical Care Medicine. 31 (1): 73–9. doi:10.1097/00003246-200301000-00011. PMID 12544996.
- Xie T, Rowen L, Aguado B, Ahearn ME, Madan A, Qin S, Campbell RD, Hood L (Dec 2003). "Analysis of the gene-dense major histocompatibility complex class III region and its comparison to mouse". Genome Research. 13 (12): 2621–36. doi:10.1101/gr.1736803. PMC 403804. PMID 14656967.
- Bouwmeester T, Bauch A, Ruffner H, Angrand PO, Bergamini G, Croughton K, Cruciat C, Eberhard D, Gagneur J, Ghidelli S, Hopf C, Huhse B, Mangano R, Michon AM, Schirle M, Schlegl J, Schwab M, Stein MA, Bauer A, Casari G, Drewes G, Gavin AC, Jackson DB, Joberty G, Neubauer G, Rick J, Kuster B, Superti-Furga G (Feb 2004). "A physical and functional map of the human TNF-alpha/NF-kappa B signal transduction pathway". Nature Cell Biology. 6 (2): 97–105. doi:10.1038/ncb1086. PMID 14743216.
- Wiemann S, Arlt D, Huber W, Wellenreuther R, Schleeger S, Mehrle A, Bechtel S, Sauermann M, Korf U, Pepperkok R, Sültmann H, Poustka A (Oct 2004). "From ORFeome to biology: a functional genomics pipeline". Genome Research. 14 (10B): 2136–44. doi:10.1101/gr.2576704. PMC 528930. PMID 15489336.
External links
- HSPA1AL+protein,+human at the US National Library of Medicine Medical Subject Headings (MeSH)
PDB gallery | |
|---|---|
|


