SUMO protein
Small Ubiquitin-like Modifier (or SUMO) proteins are a family of small proteins that are covalently attached to and detached from other proteins in cells to modify their function. SUMOylation is a post-translational modification involved in various cellular processes, such as nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress, and progression through the cell cycle.[1]
SUMO proteins are similar to ubiquitin and are considered members of the ubiquitin-like protein family. SUMOylation is directed by an enzymatic cascade analogous to that involved in ubiquitination. In contrast to ubiquitin, SUMO is not used to tag proteins for degradation. Mature SUMO is produced when the last four amino acids of the C-terminus have been cleaved off to allow formation of an isopeptide bond between the C-terminal glycine residue of SUMO and an acceptor lysine on the target protein.
SUMO family members often have dissimilar names; the SUMO homologue in yeast, for example, is called SMT3 (suppressor of mif two 3). Several pseudogenes have been reported for this gene.

Function
SUMO modification of proteins has many functions. Among the most frequent and best studied are protein stability, nuclear-cytosolic transport, and transcriptional regulation. Typically, only a small fraction of a given protein is SUMOylated and this modification is rapidly reversed by the action of deSUMOylating enzymes. SUMOylation of target proteins has been shown to cause a number of different outcomes including altered localization and binding partners. The SUMO-1 modification of RanGAP1 (the first identified SUMO substrate) leads to its trafficking from cytosol to nuclear pore complex.[2][3] The SUMO modification of hNinein leads to its movement from the centrosome to the nucleus.[4] In many cases, SUMO modification of transcriptional regulators correlates with inhibition of transcription.[5] One can refer to the GeneRIFs of the SUMO proteins, e.g. human SUMO-1,[6] to find out more.
There are 4 confirmed SUMO isoforms in humans; SUMO-1, SUMO-2, SUMO-3 and SUMO-4. At the amino acid level, SUMO1 is about 50% identical with the nearly identical SUMO2. SUMO-2/3 show a high degree of similarity to each other and are distinct from SUMO-1. SUMO-4 shows similarity to SUMO-2/3 but differs in having a Proline instead of Glutamine at position 90. As a result, SUMO-4 isn't processed and conjugated under normal conditions, but is used for modification of proteins under stress-conditions like starvation.[7] During mitosis, SUMO-2/3 localize to centromeres and condensed chromosomes, whereas SUMO-1 localizes to the mitotic spindle and spindle midzone, indicating that SUMO paralogs regulate distinct mitotic processes in mammalian cells.[8] One of the major SUMO conjugation products associated with mitotic chromosomes arose from SUMO-2/3 conjugation of topoisomerase II, which is modified exclusively by SUMO-2/3 during mitosis.[9] SUMO-2/3 modifications seem to be involved specifically in the stress response.[10] SUMO-1 and SUMO-2/3 can form mixed chains, however, because SUMO-1 does not contain the internal SUMO consensus sites found in SUMO-2/3, it is thought to terminate these poly-SUMO chains.[11] Serine 2 of SUMO-1 is phosphorylated, raising the concept of a 'modified modifier'.[12]
DNA damage response
Cellular DNA is regularly exposed to DNA damaging agents. A DNA damage response (DDR) that is well regulated and intricate is usually employed to deal with the potential deleterious effects of the damage. When DNA damage occurs, SUMO protein has been shown to act as a molecular glue to facilitate the assembly of large protein complexes in repair foci.[13] Also, SUMOylation can alter a protein's biochemical activities and interactions. SUMOylation plays a role in the major DNA repair pathways of base excision repair, nucleotide excision repair, non-homologous end joining and homologous recombinational repair.[13] SUMOylation also facilitates error prone translesion synthesis.
Structure
SUMO proteins are small; most are around 100 amino acids in length and 12 kDa in mass. The exact length and mass varies between SUMO family members and depends on which organism the protein comes from. Although SUMO has very little sequence identity with ubiquitin at the amino acid level, it has a nearly identical structural fold.
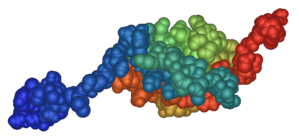
The structure of human SUMO1 is depicted on the right. It shows SUMO1 as a globular protein with both ends of the amino acid chain (shown in red and blue) sticking out of the protein's centre. The spherical core consists of an alpha helix and a beta sheet. The diagrams shown are based on an NMR analysis of the protein in solution.
Prediction of SUMO attachment
Most SUMO-modified proteins contain the tetrapeptide consensus motif Ψ-K-x-D/E where Ψ is a hydrophobic residue, K is the lysine conjugated to SUMO, x is any amino acid (aa), D or E is an acidic residue. Substrate specificity appears to be derived directly from Ubc9 and the respective substrate motif. Currently available prediction programs are:
- SUMOplot - online free access software developed to predict the probability for the SUMO consensus sequence (SUMO-CS) to be engaged in SUMO attachment.[14] The SUMOplot score system is based on two criteria: 1) direct amino acid match to the SUMO-CS observed and shown to bind Ubc9, and 2) substitution of the consensus amino acid residues with amino acid residues exhibiting similar hydrophobicity. SUMOplot has been used in the past to predict Ubc9 dependent sites.
- seeSUMO - uses random forests and support vector machines trained on the data collected from the literature[15]
- SUMOsp - uses PSSM to score potential SUMOylation peptide stites. It can predict sites followed the ψKXE motif and unusual SUMOylation sites contained other non-canonical motifs.[16]
- JASSA - online free access predictor of SUMOylation sites (classical and inverted consensus) and SIMs (SUMO interacting motif). JASSA uses a scoring system based on a Position Frequency Matrix derived from the alignment of experimental SUMOylation sites or SIMs. Novel features were implemented towards a better evaluation of the prediction, including identification of database hits matching the query sequence and representation of candidate sites within the secondary structural elements and/or the 3D fold of the protein of interest, retrievable from deposited PDB files.[17]
SUMO attachment
SUMO attachment to its target is similar to that of ubiquitin (as it is for the other ubiquitin-like proteins such as NEDD 8). The SUMO precursor has some extra amino acids that need to be removed, therefore a C-terminal peptide is cleaved from the SUMO precursor by a protease (in human these are the SENP proteases or Ulp1 in yeast) to reveal a di-glycine motif. The obtained SUMO then becomes bound to an E1 enzyme (SUMO Activating Enzyme (SAE)) which is a heterodimer. It is then passed to an E2 which is a conjugating enzyme (Ubc9). Finally, one of a small number of E3 ligating proteins attaches it to the protein. In yeast, there are four SUMO E3 proteins, Cst9,[18] Mms21, Siz1 and Siz2. While in ubiquitination an E3 is essential to add ubiquitin to its target, evidence suggests that the E2 is sufficient in SUMOylation as long as the consensus sequence is present. It is thought that the E3 ligase promotes the efficiency of SUMOylation and in some cases has been shown to direct SUMO conjugation onto non-consensus motifs. E3 enzymes can be largely classed into PIAS proteins, such as Mms21 (a member of the Smc5/6 complex) and Pias-gamma and HECT proteins. On Chromosome 17 of the human genome, SUMO2 is near SUMO1+E1/E2 and SUMO2+E1/E2, among various others. Some E3's, such as RanBP2, however, are neither.[19] Recent evidence has shown that PIAS-gamma is required for the SUMOylation of the transcription factor yy1 but it is independent of the zinc-RING finger (identified as the functional domain of the E3 ligases). SUMOylation is reversible and is removed from targets by specific SUMO proteases. In budding yeast, the Ulp1 SUMO protease is found bound at the nuclear pore, whereas Ulp2 is nucleoplasmic. The distinct subnuclear localisation of deSUMOylating enzymes is conserved in higher eukaryotes.[20]
Role in protein purification
Recombinant proteins expressed in E. coli may fail to fold properly, instead forming aggregates and precipitating as inclusion bodies.[21] This insolubility may be due to the presence of codons read inefficiently by E. coli, differences in eukaryotic and prokaryotic ribosomes, or lack of appropriate molecular chaperones for proper protein folding.[22] In order to purify such proteins it may be necessary to fuse the protein of interest with a solubility tag such as SUMO or MBP (maltose-binding protein) to increase the protein's solubility.[22] SUMO can later be cleaved from the protein of interest using a SUMO-specific protease such as Ulp1 peptidase.[22]
Human SUMO proteins
References
- Hay RT (April 2005). "SUMO: a history of modification". Molecular Cell. 18 (1): 1–12. doi:10.1016/j.molcel.2005.03.012. PMID 15808504.
- Matunis MJ, Coutavas E, Blobel G (December 1996). "A novel ubiquitin-like modification modulates the partitioning of the Ran-GTPase-activating protein RanGAP1 between the cytosol and the nuclear pore complex". The Journal of Cell Biology. 135 (6 Pt 1): 1457–70. doi:10.1083/jcb.135.6.1457. PMC 2133973. PMID 8978815.
- Mahajan R, Delphin C, Guan T, Gerace L, Melchior F (January 1997). "A small ubiquitin-related polypeptide involved in targeting RanGAP1 to nuclear pore complex protein RanBP2". Cell. 88 (1): 97–107. doi:10.1016/S0092-8674(00)81862-0. PMID 9019411.
- Cheng TS, Chang LK, Howng SL, Lu PJ, Lee CI, Hong YR (February 2006). "SUMO-1 modification of centrosomal protein hNinein promotes hNinein nuclear localization". Life Sciences. 78 (10): 1114–20. doi:10.1016/j.lfs.2005.06.021. PMID 16154161.
- Gill G (October 2005). "Something about SUMO inhibits transcription". Current Opinion in Genetics & Development. 15 (5): 536–41. doi:10.1016/j.gde.2005.07.004. PMID 16095902.
- SUMO1 SMT3 suppressor of mif two 3 homolog 1 (S. cerevisiae)
- Wei W, Yang P, Pang J, Zhang S, Wang Y, Wang MH, Dong Z, She JX, Wang CY (October 2008). "A stress-dependent SUMO4 SUMOylation of its substrate proteins". Biochemical and Biophysical Research Communications. 375 (3): 454–9. doi:10.1016/j.bbrc.2008.08.028. PMID 18708028.
- Zhang XD, Goeres J, Zhang H, Yen TJ, Porter AC, Matunis MJ (March 2008). "SUMO-2/3 modification and binding regulate the association of CENP-E with kinetochores and progression through mitosis". Molecular Cell. 29 (6): 729–41. doi:10.1016/j.molcel.2008.01.013. PMC 2366111. PMID 18374647.
- Azuma Y, Arnaoutov A, Dasso M (November 2003). "SUMO-2/3 regulates topoisomerase II in mitosis". The Journal of Cell Biology. 163 (3): 477–87. doi:10.1083/jcb.200304088. PMC 2173648. PMID 14597774.
- Saitoh H, Hinchey J (March 2000). "Functional heterogeneity of small ubiquitin-related protein modifiers SUMO-1 versus SUMO-2/3". The Journal of Biological Chemistry. 275 (9): 6252–8. doi:10.1074/jbc.275.9.6252. PMID 10692421.
- Matic I, van Hagen M, Schimmel J, Macek B, Ogg SC, Tatham MH, Hay RT, Lamond AI, Mann M, Vertegaal AC (January 2008). "In vivo identification of human small ubiquitin-like modifier polymerization sites by high accuracy mass spectrometry and an in vitro to in vivo strategy". Molecular & Cellular Proteomics. 7 (1): 132–44. doi:10.1074/mcp.M700173-MCP200. PMC 3840926. PMID 17938407.
- Matic I, Macek B, Hilger M, Walther TC, Mann M (September 2008). "Phosphorylation of SUMO-1 occurs in vivo and is conserved through evolution". Journal of Proteome Research. 7 (9): 4050–7. doi:10.1021/pr800368m. PMID 18707152.
- Jalal D, Chalissery J, Hassan AH (2017). "Genome maintenance in Saccharomyces cerevisiae: the role of SUMO and SUMO-targeted ubiquitin ligases". Nucleic Acids Res. 45 (5): 2242–2261. doi:10.1093/nar/gkw1369. PMC 5389695. PMID 28115630.
- Gramatikoff K. et al. In Frontiers of Biotechnology and Pharmaceuticals, Science Press (2004) 4: pp.181-210.
- Teng S, Luo H, Wang L (July 2012). "Predicting protein SUMOylation sites from sequence features". Amino Acids. 43 (1): 447–55. doi:10.1007/s00726-011-1100-2. PMID 21986959.
- Ren, Jian; Gao, Xinjiao; Jin, Changjiang; Zhu, Mei; Wang, Xiwei; Shaw, Andrew; Wen, Longping; Yao, Xuebiao; Xue, Yu (2009). "Systematic study of protein SUMOylation: Development of a site-specific predictor of SUMOsp 2.0". Proteomics. 9 (12): 3409–3412. doi:10.1002/pmic.200800646. PMID 19504496.
- Beauclair G, Bridier-Nahmias A, Zagury JF, Saïb A, Zamborlini A (November 2015). "JASSA: a comprehensive tool for prediction of SUMOylation sites and SIMs". Bioinformatics. 31 (21): 3483–91. doi:10.1093/bioinformatics/btv403. PMID 26142185.
- Cheng CH, Lo YH, Liang SS, Ti SC, Lin FM, Yeh CH, Huang HY, Wang TF (August 2006). "SUMO modifications control assembly of synaptonemal complex and polycomplex in meiosis of Saccharomyces cerevisiae". Genes & Development. 20 (15): 2067–81. doi:10.1101/gad.1430406. PMC 1536058. PMID 16847351.
- Pichler A, Knipscheer P, Saitoh H, Sixma TK, Melchior F (October 2004). "The RanBP2 SUMO E3 ligase is neither HECT- nor RING-type". Nature Structural & Molecular Biology. 11 (10): 984–91. doi:10.1038/nsmb834. PMID 15378033.
- Mukhopadhyay D, Dasso M (June 2007). "Modification in reverse: the SUMO proteases". Trends in Biochemical Sciences. 32 (6): 286–95. doi:10.1016/j.tibs.2007.05.002. PMID 17499995.
- Burgess, Richard; Deutscher, Murray (2009). "Guide to Protein Purification". Methods in Enzymology (2nd ed.). 463: 259–282. doi:10.1016/S0076-6879(09)63017-2. PMID 19892177.
- Kuo, Dennis; Nie, Minghua; Courey, Albert (2014). Protein Affinity Tags. Methods in Molecular Biology (Methods and Protocols). New York, NY: Humana Press. pp. 71–80. ISBN 978-1-4939-1034-2.
Further reading
- Ulrich HD (October 2005). "Mutual interactions between the SUMO and ubiquitin systems: a plea of no contest". Trends in Cell Biology. 15 (10): 525–32. doi:10.1016/j.tcb.2005.08.002. PMID 16125934.
- Gill G (October 2005). "Something about SUMO inhibits transcription". Current Opinion in Genetics & Development. 15 (5): 536–41. doi:10.1016/j.gde.2005.07.004. PMID 16095902.
- Li M, Guo D, Isales CM, Eizirik DL, Atkinson M, She JX, Wang CY (July 2005). "SUMO wrestling with type 1 diabetes". Journal of Molecular Medicine. 83 (7): 504–13. doi:10.1007/s00109-005-0645-5. PMID 15806321.
- Verger A, Perdomo J, Crossley M (February 2003). "Modification with SUMO. A role in transcriptional regulation". EMBO Reports. 4 (2): 137–42. doi:10.1038/sj.embor.embor738. PMC 1315836. PMID 12612601.
- Peroutka Iii RJ, Orcutt SJ, Strickler JE, Butt TR (2011). "SUMO fusion technology for enhanced protein expression and purification in prokaryotes and eukaryotes". Heterologous Gene Expression in E.coli. Methods in Molecular Biology. 705. pp. 15–30. doi:10.1007/978-1-61737-967-3_2. ISBN 978-1-61737-966-6. PMID 21125378.
External links
- LifeSensors' SUMO-based Protein and Peptide Expression Systems
- Proteins Expressed With SUMO
- Boston Biochem overview of SUMO reagents and the SUMOylation Cycle
- SUMO1 homology group from HomoloGene
- human SUMO proteins on ExPASy: SUMO1 SUMO2 SUMO3 SUMO4
- UniProt entry for rat Sumo1
Programs for prediction SUMOylation:
- SUMOplot Analysis Program — predicts and scores SUMOylation sites in your protein (by Abgent)
- seeSUMO - prediction of sumoulation sites
- SUMOsp - prediction of sumoulation sites
- JASSA - Predicts and scores SUMOylation sites and SIM (SUMO interacting motif)