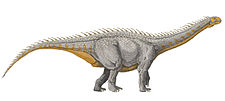
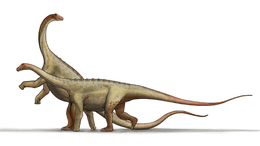Titanosauria
Titanosaurs (or titanosaurians; members of the group Titanosauria) were a diverse group of sauropod dinosaurs, including genera from Africa, Asia, South America, North America, Europe, Australia and Antarctica. The titanosaurians were the last surviving group of long-necked sauropods, with taxa still thriving at the time of the extinction event at the end of the Cretaceous. This group includes the largest land animals known to have ever existed, such as Patagotitan—estimated at 37 m (121 ft) long[1] with a weight of 69 tonnes (76 tons)[2]—and the comparably-sized Argentinosaurus and Puertasaurus from the same region.
| Titanosaurs | |
|---|---|
 | |
| Mounted Patagotitan on display at the Field Museum of Natural History, Chicago, IL | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Suborder: | †Sauropodomorpha |
| Clade: | †Sauropoda |
| Clade: | †Somphospondyli |
| Clade: | †Titanosauria Bonaparte & Coria, 1993 |
| Subgroups | |
| |
The group's name alludes to the mythological Titans of ancient Greek mythology, via the type genus (now considered a nomen dubium) Titanosaurus. Together with the brachiosaurids and relatives, titanosaurians make up the larger sauropod clade Titanosauriformes. Titanosaurs have long been a poorly-known group, and the relationships between titanosaur species are still not well-understood.
Description

Titanosaurs had small heads, even when compared with other sauropods. The head was also wide, similar to the heads of Camarasaurus and Brachiosaurus, though somewhat more elongated. Titanosaurian nostrils were large ("macronarian") and all had crests formed by the nasal bones. Their teeth were either somewhat spatulate (spoon-like) or like pegs or pencils, but were always very small.
Titanosaur necks were of average length for sauropods, and their tails were whip-like though not as long as in the diplodocids. While the pelvis was slimmer than some sauropods, the pectoral (chest) area was much wider, giving them a uniquely "wide-legged" stance. As a result, the fossilized trackways of titanosaurians are distinctly broader than other sauropods. Their forelimbs were also stocky, and often longer than their hind limbs. Unlike other sauropods, some titanosaurs had no digits, walking only on horseshoe-shaped "stumps" made up of the columnar metacarpal bones.[3][4] Their vertebrae (back bones) were solid (not hollowed-out), which may be a reversal to more basal saurischian characteristics. Their spinal column was relatively flexible, likely making them more agile than other sauropods and more able to rear onto their hind legs. One of the most characteristic features shared by most titanosaurs were their procoelous caudal vertebrae, with ball-and-socket articulations between the vertebral centra.
From skin impressions found with fossils, it has been determined that the skin of many titanosaurians was armored with a small mosaic of small, bead-like scales surrounding larger scales.[5] While most titanosaurs were very large animals, many were fairly average in size compared to other giant dinosaurs. Some island-dwelling dwarf titanosaurs, such as Magyarosaurus, were probably the result of allopatric speciation and insular dwarfism.
Body size
Titanosauria have the largest range of body size of any sauropod clade, and includes both the largest known sauropods and some of the smallest.[6] One of the largest titanosaurs, Patagotitan, had a body mass estimated to be 69 tonnes (76 tons), whereas one of the smallest, Magyarosaurus, had a body mass of approximately 900 kilograms (2,000 lb).[2][7] Even relatively closely related titanosaurs could be very different body sizes, as the small rinconsaurs were closely related to the gigantic lognkosaurs.[2] The smallest titanosaurs, such as Magyarosaurus, inhabited Europe, which was largely made up of islands during the Cretaceous, and were likely island dwarfs.
Skull
The heads of titanosaurians are poorly known. However, several different cranial morphologies are apparent. In some species, such as Sarmientosaurus, the head resembled that of brachiosaurids.[8] In others, such as Rapetosaurus and Nemegtosaurus, the head resembled that of diplodocids. In some titanosaurs, the skull was especially diplodocid-like due to square-shaped jaws;[9] the titanosaur Antarctosaurus is especially similar to the rebbachisaurid Nigersaurus.[10]
Vertebrae
The dorsal vertebrae of titanosaurs show multiple derived features among sauropods. Similarly to the Rebbachisauridae, titanosaurs lost the hyposphene-hypantrum articulations, a set of surfaces between vertebrae that prevent additional rotation of the bones. Andesaurus, one of the most basal titanosaurs, shows a normal hyposphene. The same area is reduced in Argentinosaurus to only two ridges, and is fully absent in taxa like Opisthocoelicaudia and Saltasaurus. Both Argentinosaurus and Epachthosaurus bear similar intermediate "hyposphenal ridges," which suggests they represent a more primitive form of dorsal vertebrae.[11]
Limbs

Sauropod hands already are highly derived from other dinosaurs, being reduced into columnar metacarpals and blocky phalanges with fewer claws. However, titanosaurs evolved the manus even further, completely losing the phalanges and heavily modifying the metacarpals. Argyrosaurus is the only titanosaur known to possess carpals. Other taxa like Epachthosaurus show a reduction of phalanges to one or two bones. Opisthoeoclicaudia shows even more reduction of the hand than other titanosaurs, with both carpals and phalanges completely absent.[12] However, Diamantinasaurus, while lacking carpals, preserves a manual formula of 2–1–1–1–1, including a thumb claw and phalanges on all other digits. This, coupled with the preservation of a single phalanx on digit IV of Epachthosaurus and potentially Opisthocoelicaudia (further study is necessary), show that preservation biases may be responsible for the lack of hand phalanges in these taxa. This suggests that Alamosaurus, Neuquensaurus, Saltasaurus and Rapetosaurus - all known from imperfect or disarticulated remains previously associated with a lack of phalanges - may have had phalanges but lost them after death.[13]
Titanosaurs have a poor fossil record of their pedes (feet), only being complete in five definitive titanosaurians. Among these, Notocolossus is the largest, and also has the most specialized pes: like all titanosaurs, its pes is composed of short, thick metatarsals of approximately the same lengths; however, metatarsals I and V are notably more robust than in other taxa.[14]
Osteoderms

Some titanosaurs had osteoderms. Osteoderms were first confirmed in the genus Saltasaurus but are now known to have been present in a variety of titanosaurs within the clade Lithostrotia.[15] The exact arrangement of osteoderms on the body of a titanosaur is not known, but some paleontologists consider it likely that the osteoderms were arranged in two parallel rows on the animal's back, an arrangement similar to the plates of stegosaurs.[16] Several other arrangements have been proposed, such as a single row along the midline, and it is possible that different species had different arrangements. The osteoderms were certainly far more sparse than those of ankylosaurs, and did not completely cover the back in scutes. Because of their sparse arrangement, it was unlikely that they served a significant role in defense. However, they may have played an important role in nutrient storage for titanosaurs living in highly seasonal climates and for female titanosaurs laying eggs.[17][18] Osteoderms were present on both large and small species, so they were not solely used by smaller species as protection against predators.[19]
Classification
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Phylogenetic position of Titanosauria within Eusauropoda[20] |
Titanosaurs are classified as sauropod dinosaurs. They are a highly diverse group form the dominant clade of Cretaceous sauropods.[21] Within Sauropoda, titanosaurs were once classified as close relatives of Diplodocidae due to their shared characteristic of narrow teeth, but this is now known to be the result of convergent evolution.[22] Titanosaurs are now known to be most closely related to euhelopodids and brachiosaurids; together they form a clade named Titanosauriformes.[23]
For much of the 20th century, most known species of titanosaurs were classified in the family Titanosauridae, which is no longer in widespread use.[24] Titanosauria was first proposed in 1993 as a taxon to encompass titanosaurids and their close relatives.[25] It has been phylogenetically defined as the clade composed of the most recent common ancestor of Saltasaurus and Andesaurus and all of its descendants.[22][24][26][27][15][23] The relationships of species within Titanosauria remain largely unresolved, and it is considered one of the most poorly-understood areas of dinosaur classification. One of the few areas of agreement is that the majority of titanosaurs except Andesaurus and some other basal species form a clade called Lithostrotia, which some researchers consider equivalent to the deprecated Titanosauridae.[24][28][27] Lithostrotians include titanosaurs such as Alamosaurus, Isisaurus, Malawisaurus, Rapetosaurus, and Saltasaurus.[28]
Early history
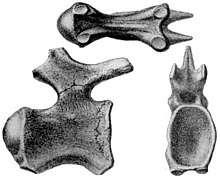
Titanosaurus indicus was first named by British paleontologist Richard Lydekker in 1877, as a new taxon of dinosaur based on two caudals and a femur collected on different occasions at the same location in India.[29] While it was later given a position as a sauropod within Cetiosauridae by Lydekker in 1888,[30] he named the new sauropod family Titanosauridae for the genus in 1893, which included only Titanosaurus and Argyrosaurus, united by procoelous caudals, opisthocoelous presacrals, a lack of pleurocoels and open chevrons.[31] Following this, Austro-Hungarian paleontologist Franz Nopcsa reviewed reptile genera in 1928, and provided a short classification of Sauropoda, where he placed the Titanosaurinae (a reranking of Lydekker's Titanosauridae) in Morosauridae, and included the genera Titanosaurus, Hypselosaurus and Macrurosaurus because they all had strongly procoelous caudals.[32] German paleontologist Friedrich von Huene provided a significant revision of Titanosauridae the following year in 1929, where he reviewed the dinosaurs of Cretaceous Argentina, and named multiple new genera. Huene included multiple species of Titanosaurus from India, England, France, Romania, Madagascar and Argentina, Hypselosaurus and Aepisaurus from France, Macrurosaurus from England, Alamosaurus from United States, and Argyrosaurus, Antarctosaurus, and Laplatasaurus from Argentina. The material between them represented almost all regions of the skeleton, which showed they were derived sauropods Huene interpreted as closest to Pleurocoelus of the various non-titanosaurid genera.[33]

For his 1986 thesis, Argentinian paleontologist Jaime Powell described and classified many new genera of South American titanosaurs. Using the family Titanosauridae to include them all, he grouped the genera into Titanosaurinae, Saltasaurinae, Antarctosaurinae, Argyrosaurinae and Titanosauridae indet. Titanosaurinae included Titanosaurus and the new genus Aeolosaurus, united by multiple features of the caudal vertebrae; the new clade Saltasaurinae was created to include Saltasaurus and the new genus Neuquensaurus, united by very distinct dorsals, caudals, and ilia; the new clade Antarctosaurinae was created to include Antarctosaurus, distinguished by large size, a different form of braincase, more elongate girdle bones, and more robust limb bones; and Argyrosaurinae was created for Argyrosaurus, bearing a more robust forelimb and hand and more primitive dorsals. The new genus Epachthosaurus was named for a more basal titanosaurid classified as Titanosauridae indet. along with unnamed specimens, Clasmodosaurus and Campylodoniscus.[34]
John Stanton McIntosh provided a synopsis of sauropod relationships in 1990, using Titanosauridae as the group to contain all taxa like previous authors. Opisthocoelicaudia was placed in Opisthocoelicaudiinae within Camarasauridae, following its original description and not later works, and Nemegtosaurus and Quaesitosaurus were placed within Dicraeosaurinae. Titanosauridae included many previously named genera, plus taxa like Tornieria and Janenschia.[35] Saltasaurus included the species previously known as Titanosaurus australis and T. robustus, which were named Neuquensaurus by Powell in 1986.[24] McIntosh provided a large diagnosis of the family: "dorsals with irregularly shaped pleurocoels and spines directed strongly backward; transverse processes directed dorsally as well as laterally, very robust in shoulder region; a second dorsosacral, its rib fused to ilium; caudals strongly procoelous with a prominent ball on distal end of centrum throughout tail; caudal arches on front half of centrum; sternal plates large; preacetabular process of ilium swept outward to become almost horizontal," but stressed that the relationships of titanosaurids to other sauropod groups couldn't be determined due to a lack of cranial material.[35]
A brief review of putative titanosaurids from Europe was authored by Jean Le Loeuff in 1993, and covered the supposed genera known so far. The Barremian (middle Early Cretaceous) species Titanosaurus valdensis, named decades previous by Huene, was kept as the oldest of the titanosaurid and given the new genus name Iuticosaurus. The French taxon Aepisaurus was removed from the family and placed in undetermined Sauropoda. Macrurosaurus was considered a chimaera of titanosaurid and non-titanosaurid material because of the presence of both procoelous and amphicoelous caudals. Huene's species Titanosaurus lydekkeri was left as a nomen dubium, but left within Titanosauridae. Maastrichtian fossils from France and Spain were removed from Hypselosaurus and Titanosaurus, with Hypselosaurus being declared dubious like T. lydekkeri. The variety of Romanian fossils named as Magyarosaurus by Huene were also moved into the same species again, M. dacus as originally named by Nopcsa.[36]
Titanosauria named

José Bonaparte and Rodolfo Coria in 1993 concluded that a new clade of derived sauropods was necessary because Argentinosaurus, Andesaurus and Epachthosaurus were distinct from Titanosauridae as they possessed hyposphene-hypantrum articulations, but were still very closely related to the titanosaurids. The taxa that possessed the articulations were united within the new family Andesauridae, and the two families were grouped together within the new clade Titanosauria. The titanosaurs were diagnosed by possessing small pleurocoels centered within an anteroposteriorly elongate depression and the presence of two well defined depressions on the posterior face of the neural arch. The entire group was compared favourably with cetiosaurids like Patagosaurus and Volkheimeria.[25]
Overlooking the naming of Titanosauria, Paul Upchurch in 1995 named the clade Titanosauroidea, to include Opisthocoelicaudia and the more derived Titanosauridae (Malawisaurus, Alamosaurus and Saltasaurus). United by: caudals with anteriorly-shifted neural spines, extremely robust forearm bones, a prominent concavity on the ulna for articulation with the humerus, a laterally flared and flattened ilium, and a less robust pubis; Upchurch considered the clade sister taxon to Diplodocoidea, because of their shared dental anatomy, although he noted that peg-like teeth might have been independently evolved.[37] This was followed up by Upchurch's 1998 study on sauropod phylogenetics, which additionally recovered Phuwiangosaurus and Andesaurus within Titanosauroidea and resolved Opisthocoelicaudia as the sister of Saltasaurus instead of the most basal titanosauroid. This result places Titanosauroidea in a group with Camarasaurus and Brachiosaurus, although Nemegtosauridae (Nemegtosaurus and Quaesitosaurus) was still classified as the basalmost family of diplodocoids. Upchurch chose to use Titanosauroidea as a replacement name for Titanosauria due to the recommended use of Linnean taxonomy and ranks.[38]
In 1997, Leonardo Salgado et al. published a phylogenetic study on Titanosauriformes, including relationships within Titanosauria. They provided a definition for the clade of "including the most recent common ancestor of Andesaurus delgadoi and Titanosauridae and all of its descendants". Titanosauria resolved including the same two subclades as Bonaparte & Coria (1993), where Andesauridae was monotypic, only including the name genus, and Titanosauridae was all other titanosaurs. Titanosauria was additionally rediagnosed, with eye-shaped pleurocoels, forked infradiapophyseal laminae, centro-parapophyseal laminae, procoelous anterior caudals, and a significantly longer pubis than ischium. Titanosauridae was less strongly defined because of the polytomy between Malawisaurus and Epachthosaurus, so some diagnostic eatures couldn't be resolved. Saltasaurinae was defined as the most recent ancestor of Neuquensaurus, Saltasaurus and its descendants, and diagnosed by short cervical prezygapophyses, vertically compressed anterior caudals, and a posteriorly shifted anterior caudal neural spine.[22]

| Titanosauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
Contributing additional work to the systematics of titanosaurs, Spanish paleontologist José Sanz et al. published an additional study in 1999, utilizing both the names Titanosauria and Titanosauroidea in displaying their results. Similar to Upchurch (1995), Sanz et al. recovered Opisthocoelicaudia as a titanosauroid outside Titanosauria, while Titanosauria was redefined to include only the taxa classified by their study. Eutitanosauria was proposed as a name for the titanosaurs more derived than Epachthosaurus, and noted the presence of osteoderms as a probable synapomorphy of this clade. Aeolosaurus, Alamosaurus, Ampelosaurus and Magyarosaurus were looked at using their character list, but were considered too incomplete to add to the final study.[39]
Argentinian paleontologist Jaime Powell published his 1986 thesis in 2003, with revisions to bring his old work up to date, including the addition of more phylogenetics and the recognition of Titanosauria as a clade name. Using the datamatrix of Sanz et al. (1999) and modifying it to include additional taxa and some character changes, Powell found that titanosaurs formed mostly a single gradual radiation beginning with Epachthosaurus as the most basal titanosaur, and Ampelosaurus and Isisaurus as the most derived. Titanosauroidea (following Upchurch 1995), was distinguished by pre- and post-spinal laminae in anterior caudals, a laterally flared ilium, a lateral expansion of the upper femur, and strongly opisthocoelous posterior dorsals. Less inclusive, Titanosauria was diagnosed by horizontally facing dorsal diapophyses, prominent procoelous anterioy caudals, and a ridge on the sternal plates. Within Titanosauria, Eutitanosauria was characterized by the absence of a hyposphene-hypantrum, no femoral fourth trochanter, and osteoderms. A small clade of Alamosaurus, Lirainosaurus and the "Peirópolis titanosaur" (Trigonosaurus) was resolved, and diagnosed by only a rotation of the tibia so the proximal end is perpendicular to the distal end. More derived clades, while resolved, were only weakly supported, or characterized by reversions of diagnostic traits of larger groups (below and left).[40]
|
Powell (2003)
|
Curry-Rogers & Forster (2001)
|
Rapetosaurus was described in 2001 by Kristina Curry-Rogers and Catherine Forster, who additionally provided a new phylogenetic analysis of Titanosauriformes (above and right). Titanosauria was strongly supported, distinguished by up to 20 characters depending on unknown traits in basal taxa. Similarly, Saltasaurinae was characterised by up to 16 traits, and the clade of Rapetosaurus and related taxa possessed four unique features. Nemegtosaurus and Quaesitosaurus were resolved within Titanosauria for the first time, after being placed in Diplodocoidea by multiple other analyses, because Rapetosaurus provided the first significant titanosaur cranial material with associated postcrania. All three genera were resolved in a clade together, although Curry-Rogers & Forster noted that it was possible the group was only resolved because no other titanosaurs had comparable cranial material. Opisthocoelicaudia was also nested deeply in Saltasaurinae, though a further investigation of titanosaur interrelationships was proposed.[41]

American paleontologist Jeff Wilson presented another revision of overall sauropod phylogeny in 2002, resolving strong support for most groups, and a similar result to Upchurch (1998) although with Euhelopus closest to titanosaurs instead of outside Neosauropoda. More internal clades were resolved for Titanosauria, with Nemegtosaurus and Rapetosaurus united within Nemegtosauridae, and Saltasauridae including two subfamilies, Opisthocoelicaudiinae and Saltasaurinae. Saltasauridae was defined as a node-stem triplet, where everything descended from the common ancestor of Opisthocoelicaudia and Saltasaurus was within Saltasauridae, and the subfamilies Saltasaurinae and Opisthocoelicaudiinae were for every taxon on one branch of the saltasaurid tree or the other.[10]
Wilson and Paul Upchurch followed this study up in 2003 with a significant revision of the type genus Titanosaurus, and revisited all the material that had been assigned to the genus while reviewing titanosaur inter-relationships. Because they found Titanosaurus to be a dubious name, they proposed that Linnaean-named groups Titanosauridae and Titanosauroidea should be considered invalid as well. Wilson & Upchurch (2003) supported the definition of Salgado et al. (1997) for Titanosauria, since it was oldest and most similar to the original content of the group when named by Bonaparte & Coria (1993). Lithostrotia (Upchurch et al. 2004) was defined to be Malawisaurus and all more derived titanosaurs, and the clade Eutitanosauria (Sanz et al. 1999) was considered a possible synonym of Saltasauridae. Wilson & Upchurch (2003) presented a reduced cladogram of Titanosauria, including only the most commonly-analyzed taxa from previous studies, resulting in a tree similar to that of Wilson (2002) but with Rapetosaurus and Nemegtosaurus excluded and Epachthosaurus included. Alamosaurus and Opisthocoelicaudia were united within Opisthocoelicaudiinae, Neuquensaurus and Saltasaurus formed Saltasaurinae, and Isisaurus placed as the next most derived titanosaurid.[24]

At the same time as Wilson & Upchurch redescribing the species of Titanosaurus, Saldago (2003) looked over the potential invalidity of the family Titanosauridae and redefined the internal clades of Titanosauria.[24][42] Titanosauria was defined as more inclusive than Titanosauroidea, contrasting with earlier used by Upchurch (1995) and Sanz et al. (1999), as all taxa in Somphospondyli closer to Saltasaurus than Euhelopus. In order to create additional stability, Saldago also defined Andesauroidea for only Andesaurus, as every titanosaur closer to that genus than Saltasaurus, and also it's opposite Titanosauroidea as every titanosaur closer to Saltasaurus than Andesaurus. Next most inclusive, Salgado revitalised Titanosauridae to include everything descended from the ancestor of Epachthosaurus and Saltasaurus, and to replace the node-stem triplet of Saltasauridae, defined the clades Epachthosaurinae and Eutitanosauria as Epachthosaurus>Saltasaurus and Saltasaurus<Epachthosaurus respectively. Saltasaurinae and Opisthocoelicaudiinae were retained with their original definitions, but Lithostrotia was considered a synonym of Titanosauridae, and Titanosaurinae was considered a paraphyletic clade of unrelated titanosaurids.[42]

Following the clade definitions proposed in previous Salgado studies, Bernardo González-Riga published two papers in 2003 describing new taxa in Titanosauria: Mendozasaurus, and Rinconsaurus (with Jorge O. Calvo). In both studies, the new taxa formed clades within Titanosauridae, although neither were named, and new diagnostic features were proposed for the family.[43][44] For Mendozasaurus, the new genus grouped with Malawisaurus as basal within Titanosauridae, but because of the features of caudal vertebrae in these basal taxa, González-Riga recommended revising the diagnosis of the family, instead of changing the content.[43] The situation of caudals in Rinconsaurus also suggested procoelous caudals were no longer diagnostic, because in the tail of Rinconsaurus the vertebrae regularly changed their articular surfaces, being from procoelous caudals interspersed with amphicoelous, opisthocoelous and biconvex vertebrae.[44] Rinconsaurus was then included in Aeolosaurini, a clade named the following year by Aldirene Franco-Rosas et al. containing everything closer to Aeolosaurus and Gondwanatitan than Saltasaurus or Opisthocoelicaudia. Only the three genera and various intermediate specimens were included in Aeolosaurini in their 2004 paper, with the tribe being considered to be within Saltasaurinae.[45]
The second edition of The Dinosauria, published in 2004, included newly described titanosaurs and other taxa reidentified as titanosaurs. Written by Upchurch, Paul Barrett and Peter Dodson, a review of Sauropoda included a more expansive Titanosauria for sauropods more derived than brachiosaurids. Titanosauria, defined as everything closer to Saltasaurus than Brachiosaurus, included a very large variety of taxa, and the new clade Lithostrotia was named for a large number of more derived taxa, although Nemegtosauridae was placed in Diplodocoidea following earlier publications of Upchurch.[28] Lithostrotia adopted the distinguishing feature of strongly procoelous caudals, previously used for Titanosauria.[24][28]
New phylogenetic frameworks
In 2005, Curry-Rogers proposed a new phylogenetic analysis that focused on the inter-relationships of Titanosauria and included the most expansive character and taxon list of any study before it. 364 characters were selected from all previous phylogenetic analyses and scored across 29 probable titanosaurs, ranging from the Late Jurassic African Janenschia to the large variety of Late Cretaceous global genera. Proposing her analysis as the basis for a new phylogenetic framework of Titanosauria, Curry-Rogers recommended only using named for clades that were very strongly supported. For the strict consensus, every taxon more derived than Brachiosaurus was in an unresolved polytomy except for a clade of Rapetosaurus and Nemegtosaurus, and one of Saltasaurinae. Within the recommended results, she only named Titanosauria, Lithostrotia, Saltasauridae, Saltasaurinae and Opisthocoelicaudiinae, because of the weakness of support (below and left).[26]
|
Curry-Rogers (2005)
|
Carballido et al. (2017)
|
Another form of composite matrix was created by Calvo, González-Riga and Juan Porfiri in 2007, based upon multiple previous studies between 1997 and 2003. The final analysis included 15 titanosaurs and 65 characters, and the typical titanosaur subclades were resolved, Titanosauridae being used over Lithostrotia following Salgado (2003), and the new clade Rinconsauria for the clade of Rinconsaurus and Muyelensaurus. The new clade (defined as Rinconsaurus and Muyelensaurus) was placed as the sister taxon of Aeolosaurini, which together grouped with Rapetosaurus as sister to Saltasauridae.[27] In the same year, Calvo et al. published another paper, describing the basal titanosaur Futalognkosaurus. The only difference in the resulting phylogeny, based on the matrix of the Calvo, González-Riga & Porfiri (2007), was the addition of Futalognkosaurus as the sister taxon to Mendozasaurus in a clade Calvo et al. named Lognkosauria, defined by the two genera classified within it.[46] A very similar result was also recovered by González-Riga et al. in 2009 in a phylogenetic analysis based partially on that of Calvo et al. (2007), although Epachthosaurus was nested with Rapetosaurus outside the clades of aeolosaurines.[47] Further updates and modifications were then made by Palbo Gallina & Apesteguía in 2011, with the additions of Ligabuesaurus, Antarctosaurus, Nemegtosaurus and Bonitasaura and character updates to match, bringing the total to 77 characters and 22 taxa. Significantly contrasting the earlier results, internal relationships of Titanosauria were rearranged. Malawisaurus nested with Andesaurus in a clade of the basalmost titanosaurs outside Titanosauroidea, where Lirainosaurus, instead of being the basal member of the saltasaur-branch was instead basalmost titanosauroid. Lognkosauria moved to be within rinconsaurs, while Nemegtosauridae was resolved as the sister of Aeolosaurus and Gondwanatitan, and the rinconsaur-lognkosaur branch. Antarctosaurus was unstable, but placed in a polytomy with the lognkosaurs and rinconsaurs before being excluded. Saltasaurinae and its relationship with Opisthocoelicaudia remained the same.[48]
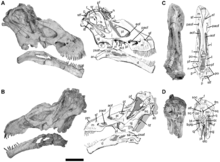
Nemegtosauridae was additionally revised by Hussam Zaher et al. (2011) with the description of Tapuiasaurus, which nested closer to Rapetosaurus than Nemegtosaurus, with all three forming a clade of derived lithostrotians. Using the matrix of Wilson (2002), following the additions of a few cranial characters and Diamantinasaurus, Tangvayosaurus and Phuwiangosaurus, remained the same as originally found by Wilson but with Diamantinasaurus sister to Saltasauridae and the other two genera as basal titanosaurs outside Lithostrotia, since Titanosauria, while undefined, was labelled to include all taxa closer to Saltasaurus than Euhelopus.[49] Following a revision of the skull of Tapuiasaurus, Wilson et al. (2016) rescored the analysis of Zaher et al. and recovered similar results for everything but Nemegtosauridae, where the family dissolved into a more basal Tapuiasaurus outside Lithostrota and Nemegtosaurus outside Saltasauridae. While non-titanosaur phylogeny remained identical in every single result, the topology within Titanosauria was very labile and prone to change with minor adjustments.[50]

Also following the 2002 analysis of Wilson, José Carballido and colleagues published a redescription of Chubutisaurus in 2011, and utilized an updated Wilson matrix, expanded to 289 characters across 41 taxa, including 15 titanosaurs. The primary focus of the analysis was on the basal titanosauriform taxa, but Titanosauria was defined, as the most recent common ancestor of Andesaurus delgadoi and Saltasaurus loricatus, and all its descendants, although the only autapomorphy of the group recovered was the absence of a prominent ventral process on the scapula.[51] This same matrix and basis of characters was further utilized and expanded for analyses on Tehuelchesaurus, Comahuesaurus and related rebbachisaurs, Europasaurus, and Padillasaurus, before being expanded upon once again in 2017 by Carballido et al. during the description of Patagotitan to 405 characters and 87 taxa, including 28 titanosaurs (above and right).[52][53][54][55][56] The definition of Titanosauria was preserved following Salgado et al. (1997) as Andesaurus plus Saltasaurus. Eutitanosauria (closer to Saltasaurus than Epachthosaurus) was resolved as a very inclusive clade composed of two distinct branches, one leading to the larger-bodied lognkosaurs and the other to the smaller-bodied saltasaurs. On the lognkosaur branch of Eutitanosauria, there is a branch of lognkosaurs and one of Rinconsauria. Following Calvo, González-Riga and Porfiri (2007), Rinconsauria was defined as Muyelensaurus plus Rinconsaurus, and Lognkosauria was defined as Mendozasaurus plus Futalognkosaurus. Rinconsauria included taxa typically found within Aeolosaurini as well, so Aeolosaurini was redefined as Aeolosaurus rionegrinus plus Gondwanatitan to preserve the original restricted content, otherwise the entire rinconsaur-lognkosaur branch would be classified within Aeolosaurini. Lithostrotia, Saltasauridae and Saltasaurinae had their definitions preserved from earlier studies, and included their typical content.[56]
Philip Mannion and colleagues redescribed Lusotitan in 2013, creating a new analysis of 279 characters drawn from significant previous analyses by Upchurch and Wilson supplemented by other studies. 63 sauropods were included, focusing on non-titanosaurian sauropods, although 14 probable titanosaurs were included. Unique to Mannion et al., continuous characters were distinguished in a run of the matrix, which resolved almost all of Somphospondyli within Titanosauria because of Andesaurus placing very basal in a large group of Andesauroidea. Titanosauroidea was tentatively retained as the opposite clade of titanosaurs, which included all other traditional titanosaurs, although it was noted because of the invalidity of Titanosaurus, Titanosauroidea should be considered an invalid name as well.[23] While the original analysis didn't focus on titanosaurs, it was utilised during the descriptions of Savannasaurus and Diamantinasaurus, Yongjinglong, an osteology of Mendozasaurus, and redescribing Tendaguria.[13][57][58][59][20] From these updates, an analysis of 548 characters and 124 taxa was published by Mannion et al. in 2019 for a redescription of Jiangshanosaurus and Dongyangosaurus, and additional revisions of Ruyangosaurus were made. No differentiation between continuous and discrete characters was made like performed by Mannion et al. (2013), but a large clade of Andesauroidea was still resolved with implied weights. Both redescribed Asian taxa, as well as Yongjinglong, previously considered derived titanosaurs related to Saltasauridae, were removed to outside the clade.[60]
| Titanosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
In the description of Mansourasaurus, Sallam et al. (2017) published a phylogenetic analysis of Titanosauria including the most taxa of any analysis of the clade.[61] In an updated version of the analysis, with the taxon Mnyamawamtuka added, Gorscak & O'Connor (2019) got similar results, with slightly different relationships within small clades.[62]
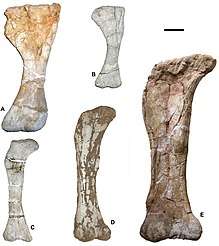
| Titanosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiology
Diet
Fossilized dung associated with late Cretaceous titanosaurids has revealed phytoliths, silicified plant fragments, that offer clues to a broad, unselective plant diet. Besides the plant remains that might have been expected, such as cycads and conifers, discoveries published in 2005[63] revealed an unexpectedly wide range of monocotyledons, including palms and grasses (Poaceae), including ancestors of rice and bamboo, which has given rise to speculation that herbivorous dinosaurs and grasses co-evolved.
Nesting
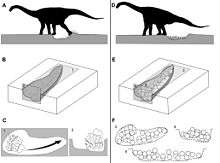
A large titanosaurid nesting ground was discovered in Auca Mahuevo, in Patagonia, Argentina and another colony has reportedly been discovered in Spain. Several hundred female saltasaurs dug holes with their back feet, laid eggs in clutches averaging around 25 eggs each, and buried the nests under dirt and vegetation. The small eggs, about 11–12 centimetres (4.3–4.7 in) in diameter, contained fossilised embryos, complete with skin impressions. The impressions showed that titanosaurs were covered in a mosaic armour of small bead-like scales.[5] The huge number of individuals gives evidence of herd behavior, which, along with their armor, could have helped provide protection against large predators such as Abelisaurus.[64]
Range
The titanosaurs were the last great group of sauropods, which existed from about 136[65] to 66 million years ago, before the Cretaceous–Paleogene extinction event, and were the dominant herbivores of their time. The fossil evidence suggests they replaced the other sauropods, like the diplodocids and the brachiosaurids, which died out between the late Jurassic and the mid-Cretaceous Periods.
Titanosaurs were widespread. In December 2011, Argentine scientists announced titanosaur fossils had been found on Antarctica—meaning that titanosaur fossils have been found on all continents. They are especially numerous in the southern continents (then part of the supercontinent of Gondwana). Australia had titanosaurs around 96 million years ago: fossils have been discovered in Queensland of a creature around 25 metres (82 ft) long.[66][67] Remains have also been discovered in New Zealand.[68] One of the largest ever titanosaur footprints was discovered in the Gobi desert in 2016.[69] One of the oldest remains of this group was found from the Valley of the Dinosaurs, Paraíba state of Brazil, representing a 136-million-year-old subadult individual.[65]
Exhibits
- The Field Museum of Natural History, Chicago, has a permanent exhibit of a 37.2 metres (122 ft) titanosaur skeleton named Maximo.[70] The specimen belongs to one of the largest known dinosaurs, Patagotitan.[71][72] The exhibit unseated the famous Tyrannosaurus rex skeleton named Sue. The Field Museum also has a mount of a real skeleton of a juvenile Rapetosaurus from Madagascar, and a 6.6 feet (2.0 m) right femur of Argyrosaurus from Argentina.
- The Naturmuseum Senckenberg in Germany Argentinosaurus huinculensis
- Royal Ontario Museum, Toronto, Canada Futalognkosaurus
References
- "Giant dinosaur slims down... a bit". BBC News. 10 August 2017. Retrieved 8 April 2020.
- Carballido, J.L.; Pol, D.; Otero, A.; Cerda, I.A.; Salgado, L.; Garrido, A.C.; Ramezani, J.; Cúneo, N.R.; Krause, J.M. (2017). "A new giant titanosaur sheds light on body mass evolution among sauropod dinosaurs". Proceedings of the Royal Society B: Biological Sciences. 284 (1860): 20171219. doi:10.1098/rspb.2017.1219. PMC 5563814. PMID 28794222.
- Apesteguía, S. (2005). "Evolution of the titanosaur metacarpus". Pp. 321–345 in Tidwell, V. and Carpenter, K. (eds.) Thunder-Lizards: The Sauropodomorph Dinosaurs. Indianapolis: Indiana University Press.
- Day, J.J.; Norman, D.B.; Gale, A.S.; Upchurch, P.; Powell, H.P. (2004). "A Middle Jurassic dinosaur trackway site from Oxfordshire, UK". Palaeontology. 47 (2): 319–348. doi:10.1111/j.0031-0239.2004.00366.x.
- Coria R.A., Chiappe L.M. (2007). "Embryonic Skin From Late Cretaceous Sauropods (Dinosauria) of Auca Mahuevo, Patgonia, Argentina". Journal of Paleontology. 81 (6): 1528–1532. doi:10.1666/05-150.1.
- Wilson, J. A. (2006). "An overview of titanosaur evolution and phylogeny". Actas de las III Jornadas sobre Dinosaurios y su Entorno. Salas de los Infantes, Burgos, Spain. pp. 169–190.
- Stein, K.; Csiki, Z.; Rogers, K. C.; Weishampel, D. B.; Redelstorff, R.; Carballido, J. L.; Sander, P. M. (30 April 2010). "Small body size and extreme cortical bone remodeling indicate phyletic dwarfism in Magyarosaurus dacus (Sauropoda: Titanosauria)". Proceedings of the National Academy of Sciences. 107 (20): 9258–9263. Bibcode:2010PNAS..107.9258S. doi:10.1073/pnas.1000781107. PMC 2889090. PMID 20435913.
- Rubén D. F. Martínez, Matthew C. Lamanna, Fernando E. Novas, Ryan C. Ridgely, Gabriel A. Casal, Javier E. Martínez, Javier R. Vita and Lawrence M. Witmer (2016). "A Basal Lithostrotian Titanosaur (Dinosauria: Sauropoda) with a Complete Skull: Implications for the Evolution and Paleobiology of Titanosauria". PLOS ONE. 11 (4): e0151661. Bibcode:2016PLoSO..1151661M. doi:10.1371/journal.pone.0151661. PMC 4846048. PMID 27115989.CS1 maint: multiple names: authors list (link)
- Apesteguía, Sebastián (10 September 2004). "Bonitasaura salgadoi gen. et sp. nov.: a beaked sauropod from the Late Cretaceous of Patagonia". Naturwissenschaften. 91 (10): 493–497. Bibcode:2004NW.....91..493A. doi:10.1007/s00114-004-0560-6. PMID 15729763.
- Wilson, J.A. (2002). "Sauropod dinosaur phylogeny: critique and cladistic analysis". Zoological Journal of the Linnean Society. 136 (2): 215–275. doi:10.1046/j.1096-3642.2002.00029.x.
- Apesteguía, S. (2005). "Evolution of the Hyposphene-Hypantrum Complex within Sauropoda". In Tidwell, V.; Carpenter, K. (eds.). Thunder Lizards: The Sauropodomorph Dinosaurs. Indiana University Press. pp. 248–267. ISBN 0-253-34542-1.
- Apesteguía, S. (2005). "Evolution of the Titanosaur Metacarpus". In Tidwell, V.; Carpenter, K. (eds.). Thunder Lizards: The Sauropodomorph Dinosaurs. Indiana University Press. pp. 321–345. ISBN 0-253-34542-1.
- Poropat, S.F.; Upchurch, P.; Mannion, P.D.; Hocknull, S.A.; Kear, B.P.; Sloan, T.; Sinapius, G.H.K.; Elliot, D.A. (2014). "Revision of the sauropod dinosaur Diamantinasaurus matildae Hocknull et al. 2009 from the mid-Cretaceous of Australia: Implications for Gondwanan titanosauriform dispersal". Gondwana Research. 27 (3): 995–1033. doi:10.1016/j.gr.2014.03.014.
- González-Riga, B.J.; Lamanna, M.C.; David, L.O.; Calvo, J.O.; Coria, J.P. (2016). "A gigantic new dinosaur from Argentina and the evolution of the sauropod hind foot". Scientific Reports. 6: 19165. Bibcode:2016NatSR...619165G. doi:10.1038/srep19165. PMC 4725985. PMID 26777391.
- D'Emic, Michael D. (2012). "The early evolution of titanosauriform sauropod dinosaurs" (PDF). Zoological Journal of the Linnean Society. 166 (3): 624–671. doi:10.1111/j.1096-3642.2012.00853.x.
- Vidal, Daniel; Ortega, Francisco; Sanz, José Luis (13 August 2014). Peter Dodson (ed.). "Titanosaur Osteoderms from the Upper Cretaceous of Lo Hueco (Spain) and Their Implications on the Armor of Laurasian Titanosaurs". PLOS ONE. 9 (8): –102488. Bibcode:2014PLoSO...9j2488V. doi:10.1371/journal.pone.0102488. ISSN 1932-6203. PMC 4131861. PMID 25118985.
- Curry Rogers, Kristina; D'Emic, Michael; Rogers, Raymond; Vickaryous, Matthew; Cagan, Amanda (29 November 2011). "Sauropod dinosaur osteoderms from the Late Cretaceous of Madagascar". Nature Communications. 2: 564. Bibcode:2011NatCo...2..564C. doi:10.1038/ncomms1578. ISSN 2041-1723. PMID 22127060.
- Vidal, Daniel; Ortega, Francisco; Gascó, Francisco; Serrano-Martínez, Alejandro; Sanz, José Luis (7 February 2017). "The internal anatomy of titanosaur osteoderms from the Upper Cretaceous of Spain is compatible with a role in oogenesis". Scientific Reports. 7: 42035. Bibcode:2017NatSR...742035V. doi:10.1038/srep42035. ISSN 2045-2322. PMC 5294579. PMID 28169348.
- Carrano, Matthew T.; D’Emic, Michael D. (3 February 2015). "Osteoderms of the titanosaur sauropod dinosaur Alamosaurus sanjuanensis Gilmore, 1922". Journal of Vertebrate Paleontology. 35 (1): e901334. doi:10.1080/02724634.2014.901334.
- Mannion, P.D.; Upchurch, P.; Schwarz, D.; Wings, O. (2019). "Taxonomic affinities of the putative titanosaurs from the Late Jurassic Tendaguru Formation of Tanzania: phylogenetic and biogeographic implications for eusauropod dinosaur evolution". Zoological Journal of the Linnean Society. 185 (3): 784–909. doi:10.1093/zoolinnean/zly068.
- González Riga, Bernardo J.; Lamanna, Matthew C.; Otero, Alejandro; Ortiz David, Leonardo D.; Kellner, Alexander W. A.; Ibiricu, Lucio M. (2019). "An overview of the appendicular skeletal anatomy of South American titanosaurian sauropods, with definition of a newly recognized clade". Anais da Academia Brasileira de Ciências. 91 (suppl 2): e20180374. doi:10.1590/0001-3765201920180374. PMID 31340217.
- Salgado, L.; Coria, R.A.; Calvo, J.O. (1997). "Evolution of titanosaurid sauropods. I: Phylogenetic analysis based on the postcranial evidence". Ameghiniana. 34 (1): 3–32.
- Mannion, P.D.; Upchurch, P.; Barnes, R.N.; Mateus, O. (2013). "Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms". Zoological Journal of the Linnean Society. 168: 98–206. doi:10.1111/zoj.12029.
- Wilson, J.A. and Upchurch, P. (2003). "A revision of Titanosaurus Lydekker (Dinosauria – Sauropoda), the first dinosaur genus with a 'Gondwanan' distribution" (PDF). Journal of Systematic Palaeontology. 1 (3): 125–160. doi:10.1017/S1477201903001044.CS1 maint: multiple names: authors list (link)
- Bonaparte, J.F.; Coria, R.A. (1993). "Un nuevo y gigantesco sauropodo titanosaurio de la Formacion Rio Limay (Albiano-Cenomaniano) de la Provincia del Neuquen, Argentina". Ameghiniana. 30 (3): 271–282.
- Curry-Rogers, K. (2005). "Titanosauria: a phylogenetic overview". In Curry-Rogers, K.; Wilson, J. (eds.). The Sauropods: Evolution and Paleobiology. Indiana University Press. pp. 50–103. ISBN 0-520-24623-3.
- Calvo, J.O.; González-Riga, B.J.; Porfiri, J.D. (2007). "A new titanosaur sauropod from the Late Cretaceous of Neuquén, Patagonia, Argentina". Arquivos do Museu Nacional, Rio de Janeiro. 65 (4): 485–504.
- Upchurch, P.; Barrett, P.M.; Dodson, P. (2004). "Sauropoda". In Weishampel, D.B.; Dodson, P.; Osmólska, H. (eds.). The Dinosauria (2nd ed.). University of California Press. pp. 259–322. ISBN 0-520-24209-2.
- Lydekker, R. (1877). "Notice of new and other Vertebrata from Indian Tertiary and Secondary rocks". Records of the Geological Survey of India. 10 (1): 30–43.
- Lydekker, R. (1888). "Part I. Containing the orders Ornithosauria, Crocodilia, Dinosauria, Squamata, Rhynchocephalia and Proterosauria". Catalogue of the fossil Reptilia and Amphibia in the British Museum (Natural History). British Museum of Natural History. pp. 134–136.
- Lydekker, R. (1893). "Part I. The dinosaurs of Patagonia". Contributions to a knowledge of the fossil vertebrates of Argentina. 2. Anales del Museo de La Plata. pp. 1–14.
- Nopcsa, F. (1928). "The genera of reptiles". Palaeobiologica. 1: 184.
- Huene, F. von (1929). "Los Saurisquios y Ornitisquios del Cretáceo Argentino". Anales del Museo de la Plata. 3 (2): 1–196.
- Powell, J.E. (1986). "Revision de los Titanosauridos de America del Sur". PHD Thesis. Universidad Nacional de Tucuman Facultad de Ciencas Naturales: 1–340.
- McIntosh, J.S. (1990). "Sauropoda". In Weishampel, D.B.; Dodson, P.; Osmólska, H. (eds.). The Dinosauria (1st ed.). University of California Press. pp. 345–402. ISBN 0-520-06726-6.
- Le Loeuff, J. (1993). "European titanosaurids". Revue de Paléobiologie. Spécial (7): 105–117.
- Upchurch, P. (1995). "The evolutionary history of sauropod dinosaurs". Philosophical Transactions of the Royal Society of London B. 349 (1330): 365–390. Bibcode:1995RSPTB.349..365U. doi:10.1098/rstb.1995.0125.
- Upchurch, P. (1998). "The phylogenetic relationships of sauropod dinosaurs". Zoological Journal of the Linnean Society. 124 (1): 43–103. doi:10.1006/zjls.1997.0138.
- Sanz, J.L.; Powell, J.E.; Le Loeuff, J.; Martinez, R.; Pereda-Suberbiola, X. (1999). "Sauropod remains from the Upper Cretaceous of Laño (Northcentral Spain). Titanosaur phylogenetic relationships". Estudios del Museo de Ciencias Naturales de Alava. 14 (1): 235–255.
- Powell, J.E. (2003). "Revision of South American Titanosaurid dinosaurs: palaeobiological, palaeobiogeographical and phylogenetic aspects". Records of the Queen Victoria Museum. 111: 1–173.
- Curry-Rogers, K.; Forster, C.A. (2001). "The last of the dinosaur titans: a new sauropod from Madagascar". Nature. 412 (6846): 530–534. Bibcode:2001Natur.412..530C. doi:10.1038/35087566. PMID 11484051.
- Salgado, L. (2003). "Should we abandon the name Titanosauridae? Some comments on the taxonomy of Titanosaurian Sauropods (Dinosauria)". Revista Española de Paleontología. 18 (1): 15–21.
- González-Riga, B.J. (2003). "A new titanosaur (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza Province, Argentina". Ameghiniana. 40 (2): 155–172.
- Calvo, J.O.; González-Riga, B.J. (2003). "Rinconsaurus caudamirus gen. et sp. nov., a new titanosaurid (Dinosauria, Sauropoda) from the Late Cretaceous of Patagonia, Argentina". Revista Geológica de Chile. 30 (2): 333–353. doi:10.4067/S0716-02082003000200011.
- Franco-Rosas, A.C.; Salgado, L.; Rosas, C.F.; Carvalho, I.S. (2004). "Nuevos materiales de Titanosaurios (Sauropoda) en el Cretácico Superior de Mato Grosso, Brazil". Revista Brasileira de Paleontologia. 7 (3): 329–336. doi:10.4072/rbp.2004.3.04.
- Calvo, J.O.; Porfiri, J.D.; González-Riga, B.J.; Kellner, A.W. (2007). "A new Cretaceous terrestrial ecosystem from Gondwana with the description of a new sauropod dinosaur". Anais da Academia Brasileira de Ciências. 79 (3): 529–541. doi:10.1590/S0001-37652007000300013. PMID 17768539.
- González-Riga, B.J.; Previtera, E.; Pirrone, C.A. (2009). "Malarguesaurus florenciae gen. et sp. nov., a new titanosauriform (Dinosauria, Sauropoda) from the Upper Cretaceous of Mendoza, Argentina". Cretaceous Research. 30 (1): 135–148. doi:10.1016/j.cretres.2008.06.006.
- Gallina, P.A.; Apesteguía, S. (2011). "Cranial anatomy and phylogenetic position of the titanosaurian sauropod Bonitasaura salgadoi". Acta Palaeontologica Polonica. 56 (1): 45–60. doi:10.4202/app.2010.0011.
- Zaher, H.; Pol, D.; Carvalho, A.B.; Nascimento, P.M.; Riccomini, C.; Larson, P.; Juarez-Valieri, R.; Pires-Dominigues, R.; da Silva Jr, N.J.; Campos, D.A. (2011). "A Complete Skull of an Early Cretaceous Sauropod and the Evolution of Advanced Titanosaurians". PLOS ONE. 6 (2): e16663. Bibcode:2011PLoSO...616663Z. doi:10.1371/journal.pone.0016663. PMC 3034730. PMID 21326881.
- Wilson, J.A.; Pol, D.; Carvalho, A.B.; Zaher, H. (2016). "The skull of the titanosaur Tapuiasaurus macedoi (Dinosauria: Sauropoda), a basal titanosaur from the Lower Cretaceous of Brazil". Zoological Journal of the Linnean Society. 178 (3): 611–662. doi:10.1111/zoj.12420.
- Carballido, José L.; Pol, Diego; Cerda, Ignacio; Salgado, Leonardo (10 February 2011). "The osteology of Chubutisaurus insignis del Corro, 1975 (Dinosauria: Neosauropoda) from the 'middle' Cretaceous of central Patagonia, Argentina". Journal of Vertebrate Paleontology. 31 (1): 93–110. doi:10.1080/02724634.2011.539651.
- Carballido, J.L.; Rauhut, O.W.M.; Pol, D.; Salgado, L. (2011). "Osteology and phylogenetic relationships of Tehuelchesaurus benitezii (Dinosauria, Sauropoda) from the Upper Jurassic of Patagonia". Zoological Journal of the Linnean Society. 163 (2): 605–662. doi:10.1111/j.1096-3642.2011.00723.x.
- Carballido, J.L.; Salgado, L.; Pol, D.; Canudo, J.I.; Garrido, A. (2012). "A new basal rebbachisaurid (Sauropoda, Diplodocoidea) from the Early Cretaceous of the Neuquén Basin; evolution and biogeography of the group". Historical Biology. 24 (6): 631–654. doi:10.1080/08912963.2012.672416.
- Carballido, J.L.; Sander, M.P. (2014). "Postcranial axial skeleton of Europasaurus holgeri (Dinosauria, Sauropoda) from the Upper Jurassic of Germany: implications for sauropod ontogeny and phylogenetic relationships of basal Macronaria". Journal of Systematic Palaeontology. 12 (3): 335–387. doi:10.1080/14772019.2013.764935.
- Carballido, J.L.; Pol, D.; Parra-Ruge, M.L.; Bernal, S.P.; Páramo-Fonseca, M.E.; Etayo-Serna, F. (2015). "A new Early Cretaceous brachiosaurid (Dinosauria, Neosauropoda) from northwestern Gondwana (Villa de Leiva, Colombia)". Journal of Vertebrate Paleontology. e980505 (5): 1–12. doi:10.1080/02724634.2015.980505.
- Carballido, J.L.; Pol, D.; Otero, A.; Cerda, I.A.; Salgado, L.; Garrido, A.C.; Ramezani, J.; Cúneo, N.R.; Krause, J.R. (2017). "A new giant titanosaur sheds light on body mass evolution among sauropod dinosaurs". Proceedings of the Royal Society B. 284 (1860): 20171219. doi:10.1098/rspb.2017.1219. PMC 5563814. PMID 28794222.
- Poropat, S.F.; Mannion, P.D.; Upchurch, P.; Hocknull, S.A.; Kear, B.P.; Kundrát, M.; Tischler, T.R.; Sloan, T.; Sinapius, G.H.K.; Elliott, J.A.; Elliott, D.A. (2016). "New Australian sauropods shed light on Cretaceous dinosaur palaeobiogeography". Scientific Reports. 6: 34467. Bibcode:2016NatSR...634467P. doi:10.1038/srep34467. PMC 5072287. PMID 27763598.
- Li, L.G.; Li, D.Q.; You, H.L.; Dodson, P. (2014). "A New Titanosaurian Sauropod from the Hekou Group (Lower Cretaceous) of the Lanzhou-Minhe Basin, Gansu Province, China". PLOS ONE. 9 (1): e85979. Bibcode:2014PLoSO...985979L. doi:10.1371/journal.pone.0085979. PMC 3906019. PMID 24489684.
- González-Riga, B.J.; Mannion, P.D.; Poropat, S.F.; Ortiz David, L.; Coria, J.P. (2018). "Osteology of the Late Cretaceous Argentinean sauropod dinosaur Mendozasaurus neguyelap: implications for basal titanosaur relationships". Zoological Journal of the Linnean Society. 184 (1): 136–181. doi:10.1093/zoolinnean/zlx103. hdl:10044/1/53967.
- Mannion, P.D.; Upchurch, P.; Jin, X.; Zheng, W. (2019). "New information on the Cretaceous sauropod dinosaurs of Zhejiang Province, China: impact on Laurasian titanosauriform phylogeny and biogeography". Royal Society Open Science. 6 (8): 191057. Bibcode:2019RSOS....691057M. doi:10.1098/rsos.191057. PMC 6731702. PMID 31598266.
- Sallam, H.; Gorscak, E.; O'Connor, P.; El-Dawoudi, I.; El-Sayed, S.; Saber, S. (26 June 2017). "New Egyptian sauropod reveals Late Cretaceous dinosaur dispersal between Europe and Africa". Nature. 2 (3): 445–451. doi:10.1038/s41559-017-0455-5. PMID 29379183.
- Gorscak, E.; O'Connor, P. (2019). "A new African Titanosaurian Sauropod Dinosaur from the middle Cretaceous Galula Formation (Mtuka Member), Rukwa Rift Basin, Southwestern Tanzania". PLOS ONE. 14 (2): e0211412. Bibcode:2019PLoSO..1411412G. doi:10.1371/journal.pone.0211412. PMC 6374010. PMID 30759122.
- Prasad, Vandana; Strömberg, Caroline A. E.; Alimohammadian, Habib; Sahni, Ashok (18 November 2005). "Dinosaur Coprolites and the Early Evolution of Grasses and Grazers". Science. 310 (5751): 1177–1180. Bibcode:2005Sci...310.1177P. doi:10.1126/science.1118806. PMID 16293759. S2CID 1816461.
- Vila, Bernat; Jackson, Frankie D.; Fortuny, Josep; Sellés, Albert G.; Galobart, Àngel (2010). "3-D Modelling of Megaloolithid Clutches: Insights about Nest Construction and Dinosaur Behaviour". PLOS ONE. 5 (5): e10362. Bibcode:2010PLoSO...510362V. doi:10.1371/journal.pone.0010362. PMC 2864735. PMID 20463953.
- Ghilardi, Aline M.; Aureliano, Tito; Duque, Rudah R. C.; Fernandes, Marcelo A.; Barreto, Alcina M. F.; Chinsamy, Anusuya (1 December 2016). "A new titanosaur from the Lower Cretaceous of Brazil". Cretaceous Research. 67: 16–24. doi:10.1016/j.cretres.2016.07.001.
- Roberts, Greg (3 May 2007). "Bones reveal Queensland's prehistoric titans". The Australian. Retrieved 4 May 2007.
- Molnar, R. E.; Salisbury, S. W. (2005). "Observations on Cretaceous Sauropods from Australia". In Carpenter, Kenneth; Tidswell, Virginia (eds.). Thunder Lizards: The Sauropodomorph Dinosaurs. Indiana University Press. pp. 454–465. ISBN 978-0-253-34542-4.
- "Bone discovery confirms big dinosaur roamed NZ". The New Zealand Herald. 24 June 2008. Retrieved 18 January 2009.
- "Giant footprint could shed light on titanosaurus behaviour". BBC News Online. 5 October 2016. Retrieved 5 October 2016.
- Bentle, Kyle. "How big is new Field Museum dinosaur? See for yourself". chicagotribune.com. Retrieved 22 June 2018.
- Geggel, Laura (15 January 2016). "122-Foot Titanosaur: Staggeringly Big Dino Barely Fits into Museum". Scientific American.
- "The Titanosaur". AMNH. Retrieved 16 January 2016.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||