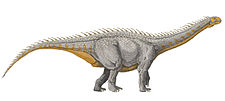
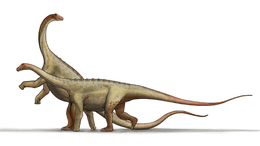Diplodocidae
Diplodocids, or members of the family Diplodocidae ("double beams"), are a group of sauropod dinosaurs. The family includes some of the longest creatures ever to walk the Earth, including Diplodocus and Supersaurus, some of which may have reached lengths of up to 34 metres (112 ft).[2][3]
| Diplodocidae | |
|---|---|
 | |
| Mounted A. louisae (specimen CM 3018), Carnegie Museum of Natural History | |
 | |
| Mounted D. carnegii type specimen, Carnegie Museum of Natural History | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Suborder: | †Sauropodomorpha |
| Clade: | †Sauropoda |
| Clade: | †Flagellicaudata |
| Family: | †Diplodocidae Marsh, 1884 |
| Type species | |
| †Diplodocus longus Marsh, 1878 | |
| Subgroups[1] | |
| Synonyms | |
| |
Description
While the diplodocids were massive sauropods, they were relatively slender when compared to the titanosaurs and brachiosaurids, although they were also extremely long. They had short legs, making them the "dachshund" of giant dinosaurs, and their rear legs were longer than front legs, giving their back a distinctive downward slope towards the neck.
Their necks were also extremely long. According to recent computer simulations, they may not have been able to lift their necks like other sauropods. However, these simulations do not take vertebral cartilage into account, which would likely allow a greater range of motion. Instead of reaching up into trees, they may have used their necks to graze over a broad area. They may also have used their necks to reach into dense stands of conifers, or over marshy ground.
Their heads, like those of other sauropods, were tiny with the nasal openings on the top of the head (though in life the nostrils themselves would have been close to the tip of the snout[4]).
The heads of diplodocids have been widely depicted with the nostrils on top due to the position of the nasal openings at the apex of the skull. There has been speculation over whether such a configuration meant that diplodocids may have had a trunk.[5] A 2006 study[6] surmised there was no paleoneuroanatomical evidence for a trunk. It noted that the facial nerve in an animal with a trunk, such as an elephant, is large as it innervates the trunk. The evidence suggests that the facial nerve is very small in diplodocids. Studies by Lawrence Witmer (2001) indicated that, while the nasal openings were high on the head, the actual, fleshy nostrils were situated much lower down on the snout.[7]
Diplodocids also had long, whip-like tails, which were thick at the base and tapered off to be very thin at the end. Computer simulations have shown that the diplodocids could have easily snapped their tails, like a bullwhip. This could generate a sonic boom in excess of 200 decibels, and may have been used in mating displays, or to drive off predators. There is some circumstantial evidence supporting this as well: a number of diplodocids have been found with fused or damaged tail vertebrae, which may be a symptom of cracking their tails.[8]
Diet and feeding
Their teeth were only present in the front of the mouth, and looked like pencils or pegs. They probably used their teeth to crop off food, without chewing, and relied on gastroliths (gizzard stones) to break down tough plant fibers (similar to modern birds). Diplodocines have highly unusual teeth compared to other sauropods. The crowns are long and slender, and elliptical in cross-section, while the apex forms a blunt, triangular point.[9] The most prominent wear facet is on the apex, though unlike all other wear patterns observed within sauropods, diplodocine wear patterns are on the labial (cheek) side of both the upper and lower teeth.[9] This implies that the feeding mechanism of Diplodocus and other diplodocids was radically different from that of other sauropods. Unilateral branch stripping is the most likely feeding behavior of Diplodocus,[10][11][12] as it explains the unusual wear patterns of the teeth (coming from tooth–food contact). In unilateral branch stripping, one tooth row would have been used to strip foliage from the stem, while the other would act as a guide and stabilizer. With the elongated preorbital (in front of the eyes) region of the skull, longer portions of stems could be stripped in a single action.[9] Also, the palinal (backwards) motion of the lower jaws could have contributed two significant roles to feeding behaviour: 1) an increased gape, and 2) allowed fine adjustments of the relative positions of the tooth rows, creating a smooth stripping action.[9]
Young et al. (2012) used biomechanical modelling to examine the performance of the diplodocine skull. It was concluded that the proposal that its dentition was used for bark-stripping was not supported by the data, which showed that under that scenario, the skull and teeth would undergo extreme stresses. The hypotheses of branch-stripping and/or precision biting were both shown to be biomechanically plausible feeding behaviors.[13] Diplodocine teeth were also continually replaced throughout their lives, usually in less than 35 days, as was discovered by Michael D'Emic et al. Within each tooth socket, as many as five replacement teeth were developing to replace the next one. Studies of the teeth also reveal that it preferred different vegetation from the other sauropods of the Morrison, such as Camarasaurus. This may have better allowed the various species of sauropods to exist without competition.[14]
Skin
_2.jpg)
Few skin impressions of diplodocids have been found. However, at least one significant find was reported by Stephen Czerkas in 1992. Fossils from the Howe Quarry in Shell, Wyoming preserved portions of the skin from around the tip of the tail, or "whiplash".[15] Czerkas noted that the skin preserved a sequence of conical spines, and that other, larger spines were found scattered around larger tail vertebrae. The spines appeared to be oriented in a single row along the mid-line of the tail, and Czerkas speculated that this midline row may have continued over the animal's entire back and neck.[16]
Growth
Long-bone histology enables researchers to estimate the age that a specific individual reached. A study by Griebeler et al. (2013) examined long bone histological data and concluded that the diplodocid MfN.R.2625 weighed 4,753 kilograms (5.2 short tons), reached sexual maturity at 23 years and died at age 24. The same growth model indicated that the diplodocid MfN.R.NW4 weighed 18,463 kilograms (20.4 short tons), and died at age 23, before reaching sexual maturity.[17]
Classification
.jpg)
Diplodocidae was the third name given to what is now recognized as the single family of long-necked, whip-tailed sauropods. Edward Drinker Cope named the family Amphicoeliidae in 1878 for his genus Amphicoelias, sometimes considered a diplodocid.[18] However, the name Amphicoeliidae did not come into wider use and was not used in the scientific literature after 1899, making it a nomen oblitum ("forgotten name") according to the ICZN, preventing it from displacing the name Diplodocidae as a senior synonym.[19] More recent studies have also shown that Amphicoelias itself does not belong to this family, but is instead a more primitive diplodocoid.[20] A similar situation occurred for the family name Atlantosauridae, named by Othniel Charles Marsh in 1877, and which Hay argued had priority over Amphicoelidae.[21] George Olshevsky declared Atlantosauridae a nomen oblitum in 1991, though scientists such as Steel and Nowinski had treated Atlantosauridae as a valid name as late as 1971, and the former even added a subfamily, Atlantosaurinae.[19][22]
Some dinosaurs have been considered diplodocids in the past but have not been found to be members of that group in later, larger analyses of the family's relationships. Australodocus, for example, was initially described as a diplodocid, but may actually have been a Macronarian.[20][23] Amphicoelias was traditionally considered a diplodocid due to its similar anatomy, but phylogenetic studies showed it to be a more basal member of the Diplodocoidea.[20]
The relationships of species within Diplodocidae has also been subject to frequent revision. A study by Lovelace, Hartman and Wahl in 2008 found that Suuwassea and Supersaurus were relatives of Apatosaurus, within the subfamily Apatosaurinae. However, a subsequent analysis by Whitlock in 2011 showed that Supersaurus is slightly closer to Diplodocus than to Apatosaurus, and that Suuwassea is actually a primitive dicraeosaurid.[20]

The subfamily Diplodocinae, was erected to include Diplodocus and its closest relatives, including Barosaurus.[24][25] The Portuguese Dinheirosaurus and the African Tornieria have also been identified as close relatives of Diplodocus by some authors.[26][27]
Cladogram of the Diplodocidae after Tschopp, Mateus, and Benson (2015).[1]
| Diplodocidae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Distinguishing anatomical features

A diagnosis is a statement of the anatomical features of an organism (or group) that collectively distinguish it from all other organisms. Some, but not all, of the features in a diagnosis are also autapomorphies - distinctive anatomical features that are unique to a given organism or group.
The clade Diplodocidae is distinguished based on the following characteristics:[28]
- nares: the external nares face dorsally; and the internarial bar is absent
- jugal: the jugal forms a substantial part of the caudoventral margin of the antorbital fenestra
- quadratojugal processes: the angle between the rostral quadratojugal process and the dorsal quadratojugal process is approximately 130°
- paroccipital process: the distal end of the paroccipital process is rounded and tongue-like in shape
- parasphenoid: the parasphenoid rostrum is a laterally compressed, thin spike and is lacking the longitudinal dorsal groove
- pterygoid: the ectopterygoid process of the pterygoid is located below the antorbital fenestra, and is reduced, such that it is not visible below the ventral margin of the skull when examined in lateral view; also the breadth of the main body of the pterygoid at least 33% of the length of the pterygoid
- teeth: at least 5-6 replacement teeth occur per alveolus (as observed in Nigersaurus)
- dorsal vertebrae: no more than 10 dorsal vertebrae are present
- caudal vertebrae: 70-80 caudal vertebrae are present
- pedal phalanges: pedal phalanx I-1 has a proximoventral margin drawn out into a thin plate or heel that underlies the distal end of metatarsal I; also pedal phalanx II-2 is reduced in craniocaudal length and has an irregular shape
References
- Tschopp, E.; Mateus, O. V.; Benson, R. B. J. (2015). "A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda)". PeerJ. 3: e857. doi:10.7717/peerj.857. PMC 4393826. PMID 25870766.
- Lovelace, David M.; Hartman, Scott A.; Wahl, William R. (2007). "Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny". Arquivos do Museu Nacional. 65 (4): 527–544.
- Tschopp, E.; Mateus, O.; Benson, R. B. J. (2015). "A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda)". PeerJ. 3: e857. doi:10.7717/peerj.857. PMC 4393826. PMID 25870766.
- Science, 03 Aug 2001: Vol. 293, Issue 5531, pp. 850-853, DOI: 10.1126/science.1062681, Nostril Position in Dinosaurs and Other Vertebrates and Its Significance for Nasal Function, Lawrence M. Witmer
- Bakker, Robert T. (1986) The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction. New York: Morrow.
- Knoll, F.; Galton, P.M.; López-Antoñanzas, R. (2006). "Paleoneurological evidence against a proboscis in the sauropod dinosaur Diplodocus". Geobios. 39 (2): 215–221. doi:10.1016/j.geobios.2004.11.005.
- Lawrence M. Witmer; et al. (2001). "Nostril Position in Dinosaurs and other Vertebrates and its Significance for Nasal Function". Science. 293 (5531): 850–853. CiteSeerX 10.1.1.629.1744. doi:10.1126/science.1062681. PMID 11486085.
- Myhrvold, Nathan P. (1997). "Supersonic sauropods? Tail dynamics in the diplodocids". Paleobiology. 23 (4): 393–409. doi:10.1017/S0094837300019801.
- Upchurch, P.; Barrett, P.M. (2000). "The evolution of sauropod feeding mechanism". In Sues, Hans Dieter (ed.). Evolution of Herbivory in Terrestrial Vertebrates. Cambridge University Press. ISBN 978-0-521-59449-3.
- Norman, D.B. (1985). The illustrated Encyclopedia of Dinosaurs. London: Salamander Books Ltd
- Dodson, P. (1990). "Sauropod paleoecology". In Weishampel, D.B.; Dodson, P.; Osmólska, H. (eds.). The Dinosauria" 1st Edition. University of California Press. ASIN B008UBRHZM.
- Barrett, P.M.; Upchurch, P. (1994). "Feeding mechanisms of Diplodocus". Gaia. 10: 195–204.
- Young, Mark T.; Rayfield, Emily J.; Holliday, Casey M.; Witmer, Lawrence M.; Button, David J.; Upchurch, Paul; Barrett, Paul M. (August 2012). "Cranial biomechanics of Diplodocus (Dinosauria, Sauropoda): testing hypotheses of feeding behaviour in an extinct megaherbivore". Naturwissenschaften. 99 (8): 637–643. Bibcode:2012NW.....99..637Y. doi:10.1007/s00114-012-0944-y. ISSN 1432-1904. PMID 22790834.
- D’Emic, M. D.; Whitlock, J. A.; Smith, K. M.; Fisher, D. C.; Wilson, J. A. (2013). Evans, A. R. (ed.). "Evolution of high tooth replacement rates in sauropod dinosaurs". PLOS ONE. 8 (7): e69235. Bibcode:2013PLoSO...869235D. doi:10.1371/journal.pone.0069235. PMC 3714237. PMID 23874921.
- Czerkas, S.A. (1993). "The new look of sauropods". Journal of Vertebrate Paleontology. 13: 26A. doi:10.1080/02724634.1993.10011533.
- Czerkas, S.A. (1993). "Discovery of dermal spines reveals a new look for sauropod dinosaurs". Geology. 20 (12): 1068–1070. Bibcode:1992Geo....20.1068C. doi:10.1130/0091-7613(1992)020<1068:dodsra>2.3.co;2.
- Griebeler, EM; Klein, N; Sander, PM (2013). "Aging, Maturation and Growth of Sauropodomorph Dinosaurs as Deduced from Growth Curves Using Long Bone Histological Data: An Assessment of Methodological Constraints and Solutions". PLoS ONE. 8 (6): e67012. Bibcode:2013PLoSO...867012G. doi:10.1371/journal.pone.0067012. PMC 3686781. PMID 23840575.
- Carpenter, K. (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus." In Foster, J.R. and Lucas, S.G., eds., 2006, Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 131-138.
- Olshevsky, G. (1991). "A Revision of the Parainfraclass Archosauria Cope, 1869, Excluding the Advanced Crocodylia." Mesozoic Meanderings, 2.
- Whitlock, J.A. (2011). "A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda)". Zoological Journal of the Linnean Society. 161: 872–915. doi:10.1111/j.1096-3642.2010.00665.x.
- Hay, O.P. (1902). "Bibliography and Catalogue of the Fossil Vertebrata of North America". Bulletin of the United States Geological Survey. 179: 1–868.
- Nowinski, A (1971). "Nemegtosaurus mongoliensis n. gen., n. sp. (Sauropoda) from the uppermost Cretaceous of Mongolia". Palaeontologia Polonica. 25: 57–81.
- Remes, Kristian (2007). "A second Gondwanan diplodocid dinosaur from the Upper Jurassic Tendaguru Beds of Tanzania, East Africa". Palaeontology. 50 (3): 653–667. doi:10.1111/j.1475-4983.2007.00652.x.
- Taylor, M.P.; Naish, D. (2005). "The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda)". PaleoBios. 25 (2): 1–7. ISSN 0031-0298.
- Harris, J.D. (2006). "The significance of Suuwassea emiliae (Dinosauria: Sauropoda) for flagellicaudatan intrarelationships and evolution" (PDF). Journal of Systematic Palaeontology. 4 (2): 185–198. doi:10.1017/S1477201906001805.
- Bonaparte, J.F.; Mateus, O. (1999). "A new diplodocid, Dinheirosaurus lourinhanensis gen. et sp. nov., from the Late Jurassic beds of Portugal". Revista del Museo Argentino de Ciencias Naturales. 5 (2): 13–29. Archived from the original on 2012-02-19. Retrieved 2013-06-13.
- Rauhut, O.W.M.; Remes, K.; Fechner, R.; Cladera, G. & Puerta, P. (2005). "Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia". Nature. 435 (7042): 670–672. Bibcode:2005Natur.435..670R. doi:10.1038/nature03623. PMID 15931221.
- Upchurch P, Barrett PM, Dodson P (2004). "Sauropoda". In Weishampel DB, Dodson P, Osmólska H. The Dinosauria (2nd Edition). University of California Press.
External links
- Cracking the whip, from the NewScientist.
- Sauropod dinosaur may have whipped its tail like Indiana Jones (says Nathan Myhrvold), from the NewScientist.
- Ivan Peterson's Whips and Dinosaur Tails.
- Skeletal restorations of several diplodocids, from Scott Hartman's Skeletal Drawing website.
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||