Triassic
The Triassic (/traɪˈæs.ɪk/ try-ASS-ik)[5] is a geologic period and system which spans 50.6 million years from the end of the Permian Period 251.9 million years ago (Mya), to the beginning of the Jurassic Period 201.3 Mya.[6] The Triassic is the first and shortest period of the Mesozoic Era. Both the start and end of the period are marked by major extinction events.[7] The Triassic period is subdivided into three epochs: Early Triassic, Middle Triassic and Late Triassic.
| Triassic Period 251.902–201.3 million years ago | |
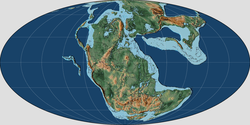 A map of the world as it appeared during the Late Triassic. (220 ma) | |
| Mean atmospheric O 2 content over period duration |
c. 16 vol % (80 % of modern level) |
| Mean atmospheric CO 2 content over period duration |
c. 1750 ppm (6 times pre-industrial level) |
| Mean surface temperature over period duration | c. 17 °C (3 °C above modern level) |
The Triassic began in the wake of the Permian–Triassic extinction event, which left the Earth's biosphere impoverished; it was well into the middle of the Triassic before life recovered its former diversity. Therapsids and archosaurs were the chief terrestrial vertebrates during this time. A specialized subgroup of archosaurs, called dinosaurs, first appeared in the Late Triassic but did not become dominant until the succeeding Jurassic Period.[8]
The first true mammals, themselves a specialized subgroup of therapsids, also evolved during this period, as well as the first flying vertebrates, the pterosaurs, who, like the dinosaurs, were a specialized subgroup of archosaurs. The vast supercontinent of Pangaea existed until the mid-Triassic, after which it began to gradually rift into two separate landmasses, Laurasia to the north and Gondwana to the south.
The global climate during the Triassic was mostly hot and dry,[9] with deserts spanning much of Pangaea's interior. However, the climate shifted and became more humid as Pangaea began to drift apart. The end of the period was marked by yet another major mass extinction, the Triassic–Jurassic extinction event, that wiped out many groups and allowed dinosaurs to assume dominance in the Jurassic.
The Triassic was named in 1834 by Friedrich von Alberti, after the three distinct rock layers (tri meaning "three") that are found throughout Germany and northwestern Europe—red beds, capped by marine limestone, followed by a series of terrestrial mud- and sandstones—called the "Trias".[10]
Dating and subdivisions
The Triassic is usually separated into Early, Middle, and Late Triassic Epochs, and the corresponding rocks are referred to as Lower, Middle, or Upper Triassic. The faunal stages from the youngest to oldest are:
| Upper/Late Triassic (Tr3) | |
| Rhaetian | (208.5–201.3 Mya) |
| Norian | (227–208.5 Mya) |
| Carnian | (237–227 Mya) |
| Middle Triassic (Tr2) | |
| Ladinian | (242–237 Mya) |
| Anisian | (247.2–242 Mya) |
| Lower/Early Triassic (Scythian) | |
| Olenekian | (251.2–247.2 Mya) |
| Induan | (251.902–251.2 Mya) |
Paleogeography
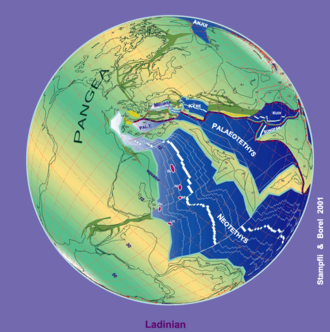
Saunders_Quarry-1.jpg)
During the Triassic, almost all the Earth's land mass was concentrated into a single supercontinent centered more or less on the equator and spanning from pole to pole, called Pangaea ("all the land"). From the east, along the equator, the Tethys sea penetrated Pangaea, causing the Paleo-Tethys Ocean to be closed.
Later in the mid-Triassic a similar sea penetrated along the equator from the west. The remaining shores were surrounded by the world-ocean known as Panthalassa ("all the sea"). All the deep-ocean sediments laid down during the Triassic have disappeared through subduction of oceanic plates; thus, very little is known of the Triassic open ocean.
The supercontinent Pangaea was rifting during the Triassic—especially late in that period—but had not yet separated. The first nonmarine sediments in the rift that marks the initial break-up of Pangaea, which separated New Jersey from Morocco, are of Late Triassic age; in the U.S., these thick sediments comprise the Newark Group.[12]
Because a super-continental mass has less shoreline compared to one broken up, Triassic marine deposits are globally relatively rare, despite their prominence in Western Europe, where the Triassic was first studied. In North America, for example, marine deposits are limited to a few exposures in the west. Thus Triassic stratigraphy is mostly based on organisms that lived in lagoons and hypersaline environments, such as Estheria crustaceans.
Africa
At the beginning of the Mesozoic Era, Africa was joined with Earth's other continents in Pangaea.[13] Africa shared the supercontinent's relatively uniform fauna which was dominated by theropods, prosauropods and primitive ornithischians by the close of the Triassic period.[13] Late Triassic fossils are found throughout Africa, but are more common in the south than north.[13] The time boundary separating the Permian and Triassic marks the advent of an extinction event with global impact, although African strata from this time period have not been thoroughly studied.[13]
Scandinavia
During the Triassic peneplains are thought to have formed in what is now Norway and southern Sweden.[14][15][16] Remnants of this peneplain can be traced as a tilted summit accordance in the Swedish West Coast.[14] In northern Norway Triassic peneplains may have been buried in sediments to be then re-exposed as coastal plains called strandflats.[15] Dating of illite clay from a strandflat of Bømlo, southern Norway, have shown that landscape there became weathered in Late Triassic times (c. 210 million years ago) with the landscape likely also being shaped during that time.[17]
South America
At Paleorrota geopark, located in Rio Grande do Sul, Brazil, the Santa Maria Formation and Caturrita Formations are exposed. In these formations, one of the earliest dinosaurs, Staurikosaurus, as well as the mammal ancestors Brasilitherium and Brasilodon have been discovered.
Climate
The Triassic continental interior climate was generally hot and dry, so that typical deposits are red bed sandstones and evaporites. There is no evidence of glaciation at or near either pole; in fact, the polar regions were apparently moist and temperate, providing a climate suitable for forests and vertebrates, including reptiles. Pangaea's large size limited the moderating effect of the global ocean; its continental climate was highly seasonal, with very hot summers and cold winters.[18] The strong contrast between the Pangea supercontinent and the global ocean triggered intense cross-equatorial monsoons.[18]
The Triassic may have mostly been a dry period, but evidence exists that it was punctuated by several episodes of increased rainfall in tropical and subtropical latitudes of the Tethys Sea and its surrounding land.[19] Sediments and fossils suggestive of a more humid climate are known from the Anisian to Ladinian of the Tethysian domain, and from the Carnian and Rhaetian of a larger area that includes also the Boreal domain (e.g., Svalbard Islands), the North American continent, the South China block and Argentina.
The best studied of such episodes of humid climate, and probably the most intense and widespread, was the Carnian Pluvial Event. A 2020 study found bubbles of carbon dioxide in basaltic rocks dating back to the end of the Triassic, and concluded that volcanic activity helped trigger climate change in that period.[20]
Life

Three categories of organisms can be distinguished in the Triassic record: survivors from the Permian–Triassic extinction event, new groups which flourished briefly, and other new groups which went on to dominate the Mesozoic Era.
Flora
On land, the surviving vascular plants included the lycophytes, the dominant cycadophytes, ginkgophyta (represented in modern times by Ginkgo biloba), ferns, horsetails and glossopterids. The spermatophytes, or seed plants, came to dominate the terrestrial flora: in the northern hemisphere, conifers, ferns and bennettitales flourished. Glossopteris (a seed fern) was the dominant southern hemisphere tree during the Early Triassic period.
Plankton
Before the Permian extinction, Archaeplastida (red and green algae) had been the major marine phytoplanktons since about 659–645 million years ago,[21] when they replaced marine planktonic cyanobacteria, which first appeared about 800 million years ago, as the dominant phytoplankton in the oceans.[22] In the Triassic, secondary endosymbiotic algae became the most important plankton.[23]
Marine fauna


In marine environments, new modern types of corals appeared in the Early Triassic, forming small patches of reefs of modest extent compared to the great reef systems of Devonian or modern times. Serpulids appeared in the Middle Triassic.[25] Microconchids were abundant. The shelled cephalopods called ammonites recovered, diversifying from a single line that survived the Permian extinction.
The fish fauna was remarkably uniform, suggesting that very few families survived the Permian extinction. There were also many types of marine reptiles. These included the Sauropterygia, which featured pachypleurosaurus and nothosaurs (both common during the Middle Triassic, especially in the Tethys region), placodonts, and the first plesiosaurs. The first of the lizardlike Thalattosauria (askeptosaurs) and the highly successful ichthyosaurs, which appeared in Early Triassic seas soon diversified, and some eventually developed to huge size during the Late Triassic. Subequatorial saurichthyids and birgeriids have also been described in Early Triassic strata.[26]
Terrestrial and freshwater fauna
Groups of terrestrial fauna, which appeared in the Triassic period or achieved a new level of evolutionary success during it include:[27][28]
- Lungfish: the lakes and rivers were populated by lungfish (Dipnoi), such as Ceratodus, which are mainly known from the dental plates, abundant in the fossils record.[29]
- Temnospondyls: one of the largest groups of early amphibians, temnospondyls originated during the Carboniferous and were still significant. Once abundant in both terrestrial and aquatic environments, the terrestrial species had mostly been replaced by reptiles. The Triassic survivors were aquatic or semi-aquatic, and were represented by Tupilakosaurus, Thabanchuia, Branchiosauridae and Micropholis, all of which died out in Early Triassic, and the successful Stereospondyli, with survivors into the Cretaceous era. The largest of these, such as the Mastodonsaurus were up to 13 feet in length.[30][31]
- Rhynchosaurs, barrel-gutted herbivores which thrived for only a short period of time, becoming extinct about 220 million years ago. They were exceptionally abundant in Triassic, the primary large herbivores in many ecosystems. They sheared plants with their beaks and several rows of teeth on the roof of the mouth.
- Phytosaurs: archosaurs that prospered during the Late Triassic. These long-snouted and semiaquatic predators resemble living crocodiles and probably had a similar lifestyle, hunting for fish and small reptiles around the water's edge. However this resemblance is only superficial and is a prime-case of convergent evolution.
- Aetosaurs: heavily armored archosaurs that were common during the last 30 million years of the Late Triassic but died out at the Triassic-Jurassic extinction. Most aetosaurs were herbivorous, and fed on low-growing plants but some may have eaten meat.
- Rauisuchians, another group of archosaurs, which were the keystone predators of most Triassic terrestrial ecosystems. Over 25 species have been found, and include giant quadrupedal hunters, sleek bipedal omnivores, and lumbering beasts with deep sails on their backs. They probably occupied the large-predator niche later filled by theropods.
- Theropods: dinosaurs that first evolved in the Triassic period but did not evolve into large sizes until the Jurassic. Most Triassic theropods, such as the Coelophysis, were only around 1–2 meters long and hunted small prey in the shadow of the giant Rauisuchians.
- Cynodonts, a large group that includes true mammals. The first cynodonts evolved in the Permian, but many groups prospered during the Triassic. Their characteristic mammalian features included hair, a large brain, and upright posture. Many were small but several forms were enormous and filled a large herbivore niche before the evolution of sauropodomorph dinosaurs, as well as large-sized carnivorous niches.
The Permian–Triassic extinction devastated terrestrial life. Biodiversity rebounded as the surviving species repopulated empty terrain, but these were short-lived. Diverse communities with complex food-web structures took 30 million years to reestablish.[7]
Temnospondyl amphibians were among those groups that survived the Permian–Triassic extinction; some lineages (e.g. trematosaurs) flourished briefly in the Early Triassic, while others (e.g. capitosaurs) remained successful throughout the whole period, or only came to prominence in the Late Triassic (e.g. Plagiosaurus, metoposaurs). As for other amphibians, the first Lissamphibia, progenitors of first frogs, are known from the Early Triassic, but the group as a whole did not become common until the Jurassic, when the temnospondyls had become very rare.
Most of the Reptiliomorpha, stem-amniotes that gave rise to the amniotes, disappeared in the Triassic, but two water-dwelling groups survived: Embolomeri that only survived into the early part of the period, and the Chroniosuchia, which survived until the end of the Triassic.
Archosauromorph reptiles, especially archosaurs, progressively replaced the synapsids that had dominated the previous Permian period. The Cynognathus was the characteristic top predator in earlier Triassic (Olenekian and Anisian) on Gondwana. Both kannemeyeriid dicynodonts and gomphodont cynodonts remained important herbivores during much of the period, and ecteniniids played a role as large-sized, cursorial predators in the Late Triassic. During the Carnian (early part of the Late Triassic), some advanced cynodonts gave rise to the first mammals. At the same time the Ornithodira, which until then had been small and insignificant, evolved into pterosaurs and a variety of dinosaurs. The Crurotarsi were the other important archosaur clade, and during the Late Triassic these also reached the height of their diversity, with various groups including the phytosaurs, aetosaurs, several distinct lineages of Rauisuchia, and the first crocodylians (the Sphenosuchia). Meanwhile, the stocky herbivorous rhynchosaurs and the small to medium-sized insectivorous or piscivorous Prolacertiformes were important basal archosauromorph groups throughout most of the Triassic.
Among other reptiles, the earliest turtles, like Proganochelys and Proterochersis, appeared during the Norian Age (Stage) of the Late Triassic Period. The Lepidosauromorpha, specifically the Sphenodontia, are first found in the fossil record of the earlier Carnian Age. The Procolophonidae were an important group of small lizard-like herbivores.
During the Triassic, archosaurs displaced therapsids as the dominant amniotes. This "Triassic Takeover" may have contributed to the evolution of mammals by forcing the surviving therapsids and their mammaliaform successors to live as small, mainly nocturnal insectivores. Nocturnal life may have forced the mammaliaforms to develop fur and a higher metabolic rate.[32]
 Postosuchus An apex predator of its time which preyed on anything smaller than itself
Postosuchus An apex predator of its time which preyed on anything smaller than itself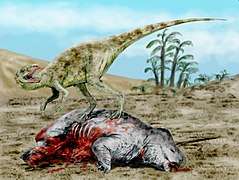 Staurikosaurus feeding on a dicynodont, in geopark Paleorrota
Staurikosaurus feeding on a dicynodont, in geopark Paleorrota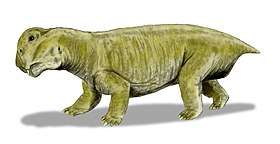 Lystrosaurus was the most common land vertebrate during the Early Triassic, when animal life had been greatly diminished
Lystrosaurus was the most common land vertebrate during the Early Triassic, when animal life had been greatly diminished Reconstruction of Proterosuchus, a genus of crocodile-like carnivorous reptile that existed in the Early Triassic
Reconstruction of Proterosuchus, a genus of crocodile-like carnivorous reptile that existed in the Early Triassic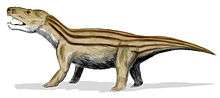 Cynognathus was a mammal-like cynodont from the Early Triassic. The first true mammals evolved during this period
Cynognathus was a mammal-like cynodont from the Early Triassic. The first true mammals evolved during this period Plateosaurus was an early sauropodomorph, or "prosauropod", of the Late Triassic
Plateosaurus was an early sauropodomorph, or "prosauropod", of the Late Triassic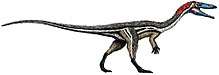 Coelophysis, one of the first dinosaurs, appeared in the Late Triassic
Coelophysis, one of the first dinosaurs, appeared in the Late Triassic Life reconstruction of Tanystropheus longobardicus
Life reconstruction of Tanystropheus longobardicus
Coal

- No known coal deposits date from the start of the Triassic period. This is known as the "coal gap" and can be seen as part of the Permian–Triassic extinction event.[34] Sharp drops in sea level at the time of the Permo-Triassic boundary may be the best explanation for the coal gap. However, there is still speculation as to why it is missing.[35] During the preceding Permian period the arid desert conditions contributed to the evaporation of many inland seas and the subsequent inundation of those areas, perhaps by a number of tsunami events that may have been responsible for the drop in sea level.[36] Large salt basins in the southwest United States and a very large basin in central Canada are evidence of this.[37] Coal of Triassic age is made of the fossilized remains of Triassic plants and trees.
- Conifers became common in Eurasia and Australia which contained a few species with a lycopod herbaceous under story. These groups of conifers arose from endemic species because of the ocean barriers that prevented seeds from crossing for over one hundred million years. For instance, Podocarpis was located south, and pines, junipers, and sequoias were located north. The dividing line ran through the Amazon Valley, across the Sahara, and north of Arabia, India, Thailand, and Australia.[38][39] It has been suggested that there was a climate barrier for the conifers,[40] although water barriers are more plausible. If so, something that could cross at least minor water barriers must have been involved in producing the coal hiatus. Hot climate could have been an important auxiliary factor across Antarctica or the Bering Strait, however. There was a spike of fern and lycopod spores immediately after the close of the Permian.[41] There was also a spike of fungal spores immediately after the Permian–Triassic boundary.[42] This spike may have lasted 50,000 years in Italy and 200,000 years in China and must have contributed to the climate warmth.
- An event excluding a catastrophe must have been involved to cause the coal hiatus because fungi would have removed all dead vegetation and coal-forming detritus in a few decades in most tropical places. In addition, fungal spores rose gradually and declined similarly along with a prevalence of woody debris. Each phenomenon would hint at widespread vegetative death. Whatever the cause of the coal hiatus, it must have started in North America approximately 25 million years sooner.[43]

Lagerstätten
The Monte San Giorgio lagerstätte, now in the Lake Lugano region of northern Italy and Switzerland, was in Triassic times a lagoon behind reefs with an anoxic bottom layer, so there were no scavengers and little turbulence to disturb fossilization, a situation that can be compared to the better-known Jurassic Solnhofen Limestone lagerstätte.
The remains of fish and various marine reptiles (including the common pachypleurosaur Neusticosaurus, and the bizarre long-necked archosauromorph Tanystropheus), along with some terrestrial forms like Ticinosuchus and Macrocnemus, have been recovered from this locality. All these fossils date from the Anisian/Ladinian transition (about 237 million years ago).
Triassic–Jurassic extinction event
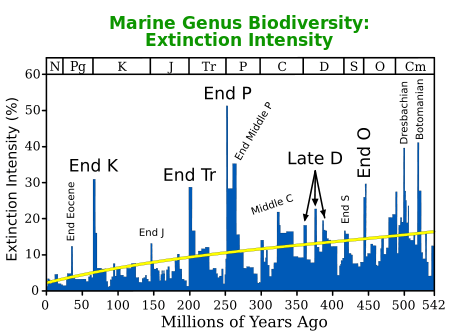
The Triassic period ended with a mass extinction, which was particularly severe in the oceans; the conodonts disappeared, as did all the marine reptiles except ichthyosaurs and plesiosaurs. Invertebrates like brachiopods, gastropods, and molluscs were severely affected. In the oceans, 22% of marine families and possibly about half of marine genera went missing.
Though the end-Triassic extinction event was not equally devastating in all terrestrial ecosystems, several important clades of crurotarsans (large archosaurian reptiles previously grouped together as the thecodonts) disappeared, as did most of the large labyrinthodont amphibians, groups of small reptiles, and some synapsids (except for the proto-mammals). Some of the early, primitive dinosaurs also became extinct, but more adaptive ones survived to evolve into the Jurassic. Surviving plants that went on to dominate the Mesozoic world included modern conifers and cycadeoids.
The cause of the Late Triassic extinction is uncertain. It was accompanied by huge volcanic eruptions that occurred as the supercontinent Pangaea began to break apart about 202 to 191 million years ago (40Ar/39Ar dates),[44] forming the Central Atlantic Magmatic Province (CAMP),[45] one of the largest known inland volcanic events since the planet had first cooled and stabilized. Other possible but less likely causes for the extinction events include global cooling or even a bolide impact, for which an impact crater containing Manicouagan Reservoir in Quebec, Canada, has been singled out. However, the Manicouagan impact melt has been dated to 214±1 Mya. The date of the Triassic-Jurassic boundary has also been more accurately fixed recently, at 201.3 Mya. Both dates are gaining accuracy by using more accurate forms of radiometric dating, in particular the decay of uranium to lead in zircons formed at time of the impact. So, the evidence suggests the Manicouagan impact preceded the end of the Triassic by approximately 10±2 Ma. It could not therefore be the immediate cause of the observed mass extinction.[46]

The number of Late Triassic extinctions is disputed. Some studies suggest that there are at least two periods of extinction towards the end of the Triassic, separated by 12 to 17 million years. But arguing against this is a recent study of North American faunas. In the Petrified Forest of northeast Arizona there is a unique sequence of late Carnian-early Norian terrestrial sediments. An analysis in 2002 found no significant change in the paleoenvironment.[47] Phytosaurs, the most common fossils there, experienced a change-over only at the genus level, and the number of species remained the same. Some aetosaurs, the next most common tetrapods, and early dinosaurs, passed through unchanged. However, both phytosaurs and aetosaurs were among the groups of archosaur reptiles completely wiped out by the end-Triassic extinction event.
It seems likely then that there was some sort of end-Carnian extinction, when several herbivorous archosauromorph groups died out, while the large herbivorous therapsids—the kannemeyeriid dicynodonts and the traversodont cynodonts—were much reduced in the northern half of Pangaea (Laurasia).
These extinctions within the Triassic and at its end allowed the dinosaurs to expand into many niches that had become unoccupied. Dinosaurs became increasingly dominant, abundant and diverse, and remained that way for the next 150 million years. The true "Age of Dinosaurs" is during the following Jurassic and Cretaceous periods, rather than the Triassic.
See also
- Geologic time scale
- List of fossil sites (with link directory)
- Phylloceratina
- Dinosaurs
Notes
- Widmann, Philipp; Bucher, Hugo; Leu, Marc; Vennemann, Torsten; Bagherpour, Borhan; Schneebeli-Hermann, Elke; Goudemand, Nicolas; Schaltegger, Urs (2020). "Dynamics of the Largest Carbon Isotope Excursion During the Early Triassic Biotic Recovery". Frontiers in Earth Science. 8 (196): 1–16. doi:10.3389/feart.2020.00196.
- McElwain, J. C.; Punyasena, S. W. (2007). "Mass extinction events and the plant fossil record". Trends in Ecology & Evolution. 22 (10): 548–557. doi:10.1016/j.tree.2007.09.003. PMID 17919771.
- |note5-nudge-down=1 Retallack, G. J.; Veevers, J.; Morante, R. (1996). "Global coal gap between Permian–Triassic extinctions and middle Triassic recovery of peat forming plants". GSA Bulletin. 108 (2): 195–207. doi:10.1130/0016-7606(1996)108<0195:GCGBPT>2.3.CO;2. Retrieved 2007-09-29.
- Payne, J. L.; Lehrmann, D. J.; Wei, J.; Orchard, M. J.; Schrag, D. P.; Knoll, A. H. (2004). "Large Perturbations of the Carbon Cycle During Recovery from the End-Permian Extinction". Science. 305 (5683): 506–9. doi:10.1126/science.1097023. PMID 15273391.
- "Triassic". Dictionary.com Unabridged. Random House.
- "ICS - Chart/Time Scale". www.stratigraphy.org.
- Sahney, S. & Benton, M.J. (2008). "Recovery from the most profound mass extinction of all time" (PDF). Proceedings of the Royal Society B: Biological Sciences. 275 (1636): 759–65. doi:10.1098/rspb.2007.1370. PMC 2596898. PMID 18198148.
- Brusatte, S. L.; Benton, M. J.; Ruta, M.; Lloyd, G. T. (2008-09-12). "Superiority, Competition, and Opportunism in the Evolutionary Radiation of Dinosaurs" (PDF). Science. 321 (5895): 1485–1488. Bibcode:2008Sci...321.1485B. doi:10.1126/science.1161833. PMID 18787166. Archived from the original (PDF) on 2014-06-24. Retrieved 2012-01-14.
- ""Lethally Hot" Earth Was Devoid of Life—Could It Happen Again?". nationalgeographic.com. 19 October 2012.
- Friedrich von Alberti, Beitrag zu einer Monographie des bunten Sandsteins, Muschelkalks und Keupers, und die Verbindung dieser Gebilde zu einer Formation [Contribution to a monograph on the colored sandstone, shell limestone and mudstone, and the joining of these structures into one formation] (Stuttgart and Tübingen, (Germany): J. G. Cotta, 1834). Alberti coined the term "Trias" on page 324 :
"… bunter Sandstein, Muschelkalk und Keuper das Resultat einer Periode, ihre Versteinerungen, um mich der Worte E. de Beaumont’s zu bedeinen, die Thermometer einer geologischen Epoche seyen, … also die bis jezt beobachtete Trennung dieser Gebilde in 3 Formationen nicht angemessen, und es mehr dem Begriffe Formation entsprechend sey, sie zu einer Formation, welche ich vorläufig Trias nennen will, zu verbinden."
( … colored sandstone, shell limestone, and mudstone are the result of a period; their fossils are, to avail myself of the words of E. de Beaumont, the thermometer of a geologic epoch; … thus the separation of these structures into 3 formations, which has been maintained until now, isn't appropriate, and it is more consistent with the concept of "formation" to join them into one formation, which for now I will name "trias".) - Herbert, Chris; Helby, Robin (1980). A Guide to the Sydney basin. Maitland, NSW: Geological Survey of NSW. p. 582. ISBN 978-0-7240-1250-3.
- "Lecture 10 - Triassic: Newark, Chinle". rainbow.ldeo.columbia.edu.
- Jacobs, Louis, L. (1997). "African Dinosaurs." Encyclopedia of Dinosaurs. Edited by Phillip J. Currie and Kevin Padian. Academic Press. p. 2-4.
- Lidmar-Bergström, Karna (1993). "Denudation surfaces and tectonics in the southernmost part of the Baltic Shield". Precambrian Research. 64 (1–4): 337–345. Bibcode:1993PreR...64..337L. doi:10.1016/0301-9268(93)90086-h.
- Olesen, Odleiv; Kierulf, Halfdan Pascal; Brönner, Marco; Dalsegg, Einar; Fredin, Ola; Solbakk, Terje (2013). "Deep weathering, neotectonics and strandflat formation in Nordland, northern Norway". Norwegian Journal of Geology. 93: 189–213.
- Japsen, Peter; Green, Paul F; Bonow, Johan M; Erlström, Mikael (2016). "Episodic burial and exhumation of the southern Baltic Shield: Epeirogenic uplifts during and after break-up of Pangaea". Gondwana Research. 35: 357–377. Bibcode:2016GondR..35..357J. doi:10.1016/j.gr.2015.06.005.
- Fredin, Ola; Viola, Giulio; Zwingmann, Horst; Sørlie, Ronald; Brönner, Marco; Lie, Jan-Erik; Margrethe Grandal, Else; Müller, Axel; Margeth, Annina; Vogt, Christoph; Knies, Jochen (2017). "The inheritance of a Mesozoic landscape in western Scandinavia". Nature. 8: 14879. Bibcode:2017NatCo...814879F. doi:10.1038/ncomms14879. PMC 5477494. PMID 28452366.
- Stanley, 452-3.
- Preto, N.; Kustatscher, E.; Wignall, P. B. (2010). "Triassic climates — State of the art and perspectives". Palaeogeography, Palaeoclimatology, Palaeoecology. 290 (1–4): 1–10. doi:10.1016/j.palaeo.2010.03.015.
- Manfredo Capriolo; et al. (2020). "Deep CO2 in the end-Triassic Central Atlantic Magmatic Province". 11 (1670). Nature Communications. doi:10.1038/s41467-020-15325-6.
- How snowball Earth gave rise to complex life - Cosmos Magazine
- December: Phytoplankton | News | University of Bristol
- The rise of algae in Cryogenian oceans and the emergence of animals - ResearchGate
- Scheyer et al. (2014): Early Triassic Marine Biotic Recovery: The Predators' Perspective. PLoS ONE https://doi.org/10.1371/journal.pone.0088987
- Vinn, O.; Mutvei, H. (2009). "Calcareous tubeworms of the Phanerozoic" (PDF). Estonian Journal of Earth Sciences. 58 (4): 286–296. doi:10.3176/earth.2009.4.07. Retrieved 2012-09-16.
- Romano, Carlo; Jenks, James F.; Jattiot, Romain; Scheyer, Torsten M. (2017). "Marine Early Triassic Actinopterygii from Elko County (Nevada, USA): implications for the Smithian equatorial vertebrate eclipse". Journal of Paleontology. 91 (5): 1–22. doi:10.1017/jpa.2017.36.
- Prehistoric Life: The Definitive Visual History of Life On Earth. London: Dorling Kindersley. 2009. pp. 206, 207. ISBN 9780756655730.
- Douglas Palmer & Peter Barrett (2009). Evolution: The Story of Life. London, Britain: The Natural History Museum. ISBN 9781845333393.CS1 maint: uses authors parameter (link)
- Agnolin, F. L., Mateus O., Milàn J., Marzola M., Wings O., Adolfssen J. S., & Clemmensen L. B. (2018). Ceratodus tunuensis, sp. nov., a new lungfish (Sarcopterygii, Dipnoi) from the Upper Triassic of central East Greenland. Journal of Vertebrate PaleontologyJournal of Vertebrate Paleontology. e1439834
- Wells, Kentwood D. (15 February 2010). The Ecology and Behavior of Amphibians. University of Chicago Press. ISBN 9780226893334 – via Google Books.
- Benton, Michael (5 February 2009). Vertebrate Palaeontology. John Wiley & Sons. ISBN 9781405144490 – via Google Books.
- Ruben, J.A. & Jones, T.D. (2000). "Selective Factors Associated with the Origin of Fur and Feathers". American Zoologist. 40 (4): 585–596. doi:10.1093/icb/40.4.585.
- Hosher, WT Magaritz M Clark D (1987). "Events near the time of the Permian-Triassic boundary". Mod. Geol. 11: 155–180 [173–174].
- Retallack, G. J.; Veevers, J. J.; Morante, R. (1996). "Global coal gap between Permian-Triassic extinction and Middle Triassic recovery of peat-forming plants". Bulletin of the Geological Society of America. 108 (2): 195–207. Bibcode:1996GSAB..108..195R. doi:10.1130/0016-7606(1996)108<0195:GCGBPT>2.3.CO;2.
- Holser, WT; Schoenlaub, H-P; Klein, P; Attrep, M; Boeckelmann, Klaus; et al. (1989). "A unique geochemical record at the Permian/Triassic boundary". Nature. 337 (6202): 39 [42]. Bibcode:1989Natur.337...39H. doi:10.1038/337039a0.
- Knauth, L. Paul (1998). "Salinity history of the Earth's early ocean". Nature. 395 (6702): 554–5. Bibcode:1998Natur.395..554K. doi:10.1038/26879. PMID 11542867.
- Dott, R.H. and Batten, R.L. (1971) Evolution of the Earth, 4th ed. McGraw Hill, NY.
- Florin, R (1963). "The distribution of Conifer and Taxad genera in time and space". Acta Horti Bergiani. 20: 121–312.
- Melville, R (1966). "Continental drift, Mesozoic continents, and the migrations of the angiosperms". Nature. 211 (5045): 116. Bibcode:1966Natur.211..116M. doi:10.1038/211116a0.
- Darlington PJ, (1965) Biogeography of the southern end of the world. Harvard University Press, Cambridge Mass., on p 168.
- Retallack, GJ (1995). "Permian -Triassic life crises on land". Science. 267 (5194): 77–79. Bibcode:1995Sci...267...77R. doi:10.1126/science.267.5194.77. PMID 17840061.
- Eshet, Y; Rampino, Michael R.; Visscher, Henk (1995). "Fungal event and palynological record of ecological crises and recovery across Permian-Triassic boundary". Geology. 23 (11): 967–970 [969]. Bibcode:1995Geo....23..967E. doi:10.1130/0091-7613(1995)023<0967:FEAPRO>2.3.CO;2.
- Retallack, GJ; Veevers, John J.; Morante, Ric (1996). "Global coal gap between Permian-Triassic extinctions and middle Triassic recovery of peat forming plants (review)". Geological Society of America Bulletin. 108 (2): 195–207. Bibcode:1996GSAB..108..195R. doi:10.1130/0016-7606(1996)108<0195:GCGBPT>2.3.CO;2.
- Nomade et al.,2007 Palaeogeography, Palaeoclimatology, Palaeoecology 244, 326-344.
- Marzoli et al., 1999, Science 284. Extensive 200-million-year-old continental flood basalts of the Central Atlantic Magmatic Province, pp. 618-620.
- Hodych & Dunning, 1992.
- "NO SIGNIFICANT NONMARINE CARNIAN-NORIAN (LATE TRIASSIC) EXTINCTION EVENT: EVIDENCE FROM PETRIFIED FOREST NATIONAL PARK". gsa.confex.com. Archived from the original on 2003-11-06. Retrieved 2003-12-12.
References
- Emiliani, Cesare. (1992). Planet Earth: Cosmology, Geology, & the Evolution of Life & the Environment. Cambridge University Press. (Paperback Edition ISBN 0-521-40949-7)
- Ogg, Jim; June, 2004, Overview of Global Boundary Stratotype Sections and Points (GSSP's) Stratigraphy.org, Accessed April 30, 2006
- Stanley, Steven M. Earth System History. New York: W.H. Freeman and Company, 1999. ISBN 0-7167-2882-6
- Sues, Hans-Dieter & Fraser, Nicholas C. Triassic Life on Land: The Great Transition New York: Columbia University Press, 2010. Series: Critical Moments and Perspectives in Earth History and Paleobiology. ISBN 978-0-231-13522-1
- van Andel, Tjeerd, (1985) 1994, New Views on an Old Planet: A History of Global Change, Cambridge University Press
External links
| Wikimedia Commons has media related to Triassic. |
| Look up triassic in Wiktionary, the free dictionary. |
- A Visit To Ammonite Canyon, Nevada Virtual field trip to one of the most famous places on Earth to study the Late Triassic extinction in a marine geologic section.
- Middle Triassic Ammonoids From Nevada
- Ammonoids At Union Wash, California Virtual field trip to a classic Early Triassic ammonoid locality in Inyo County, California.
- Late Triassic Ichthyosaur And Invertebrate Fossils In Nevada
- Early Triassic Ammonoid Fossils In Nevada
- Overall introduction
- 'The Triassic world'
- Douglas Henderson's illustrations of Triassic animals
- Paleofiles page on the Triassic extinctions
- Examples of Triassic Fossils
- Triassic (chronostratigraphy scale)