Symbiogenesis
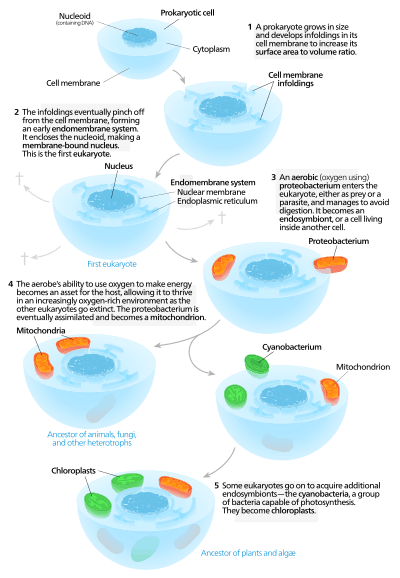
Symbiogenesis, or endosymbiotic theory, is the leading evolutionary theory of the origin of eukaryotic cells from prokaryotic organisms.[1] The theory holds that mitochondria, plastids such as chloroplasts, and possibly other organelles of eukaryotic cells are descended from formerly free-living prokaryotes (bacteria and archaea) taken one inside the other in endosymbiosis. Mitochondria appear to be phylogenetically related to Rickettsiales proteobacteria, and chloroplasts to nitrogen-fixing filamentous cyanobacteria. The theory was first articulated in 1905 and 1910 by the Russian botanist Konstantin Mereschkowski, and advanced and substantiated with microbiological evidence by Lynn Margulis in 1967. Among the many lines of evidence supporting symbiogenesis are that new mitochondria and plastids are formed only through binary fission, and that cells cannot create new ones otherwise; that the transport proteins called porins are found in the outer membranes of mitochondria, chloroplasts and bacterial cell membranes; that cardiolipin is found only in the inner mitochondrial membrane and bacterial cell membranes; and that some mitochondria and plastids contain single circular DNA molecules similar to the chromosomes of bacteria.
History
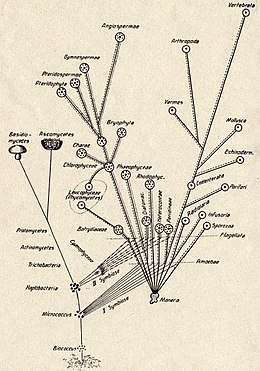
The Russian botanist Konstantin Mereschkowski first outlined the theory of symbiogenesis (from Greek: σύν syn "together", βίος bios "life", and γένεσις genesis "origin, birth") in his 1905 work, The nature and origins of chromatophores in the plant kingdom, and then elaborated it in his 1910 The Theory of Two Plasms as the Basis of Symbiogenesis, a New Study of the Origins of Organisms.[3][4][5] Mereschkowski knew of the work of botanist Andreas Schimper, who had observed in 1883 that the division of chloroplasts in green plants closely resembled that of free-living cyanobacteria, and who had himself tentatively proposed (in a footnote) that green plants had arisen from a symbiotic union of two organisms.[6] In 1918 the French scientist Paul Jules Portier published Les Symbiotes, in which he claimed that the mitochondria originated from a symbiosis process.[7][8] Ivan Wallin advocated the idea of an endosymbiotic origin of mitochondria in the 1920s.[9][10] The Russian botanist Boris Kozo-Polyansky became the first to explain the theory in terms of Darwinian evolution.[11] In his 1924 book A New Principle of Biology. Essay on the Theory of Symbiogenesis,[12] he wrote, "The theory of symbiogenesis is a theory of selection relying on the phenomenon of symbiosis."[13]
These theories did not gain traction until more detailed electron-microscopic comparisons between cyanobacteria and chloroplasts (for example studies by Hans Ris published in 1961 and 1962[14][15]), combined with the discovery that plastids and mitochondria contain their own DNA[16] (which by that stage was recognized as the hereditary material of organisms) led to a resurrection of the idea of symbiogenesis in the 1960s. Lynn Margulis advanced and substantiated the theory with microbiological evidence in a 1967 paper, On the origin of mitosing cells.[17] In her 1981 work Symbiosis in Cell Evolution she argued that eukaryotic cells originated as communities of interacting entities, including endosymbiotic spirochaetes that developed into eukaryotic flagella and cilia. This last idea has not received much acceptance, because flagella lack DNA and do not show ultrastructural similarities to bacteria or to archaea (see also: Evolution of flagella and Prokaryotic cytoskeleton). According to Margulis and Dorion Sagan,[18] "Life did not take over the globe by combat, but by networking" (i.e., by cooperation). Christian de Duve proposed that the peroxisomes may have been the first endosymbionts, allowing cells to withstand growing amounts of free molecular oxygen in the Earth's atmosphere. However, it now appears that peroxisomes may be formed de novo, contradicting the idea that they have a symbiotic origin.[19]
The fundamental theory of symbiogenesis as the origin of mitochondria and chloroplasts is now widely accepted.[1]
From endosymbionts to organelles
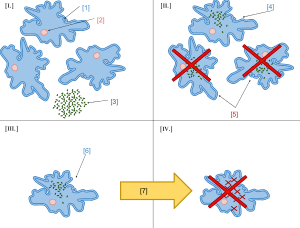
According to Keeling and Archibald,[20] the usual way to distinguish organelles from endosymbionts is by their reduced genome sizes. As an endosymbiont evolves into an organelle, most of their genes are transferred to the host cell genome.[21] The host cell and organelle need to develop a transport mechanism that enables the return of the protein products needed by the organelle but now manufactured by the cell. Cyanobacteria and α-proteobacteria are the most closely related free-living organisms to plastids and mitochondria respectively.[22] Both cyanobacteria and α-proteobacteria maintain a large (>6Mb) genome encoding thousands of proteins.[22] Plastids and mitochondria exhibit a dramatic reduction in genome size when compared with their bacterial relatives.[22] Chloroplast genomes in photosynthetic organisms are normally 120-200kb[23] encoding 20-200 proteins[22] and mitochondrial genomes in humans are approximately 16kb and encode 37 genes, 13 of which are proteins.[24] Using the example of the freshwater amoeboid, however, Paulinella chromatophora, which contains chromatophores found to be evolved from cyanobacteria, Keeling and Archibald argue that this is not the only possible criterion; another is that the host cell has assumed control of the regulation of the former endosymbiont's division, thereby synchronizing it with the cell's own division.[20] Nowack and her colleagues[25] performed gene sequencing on the chromatophore (1.02 Mb) and found that only 867 proteins were encoded by these photosynthetic cells. Comparisons with their closest free living cyanobacteria of the genus Synechococcus (having a genome size 3 Mb, with 3300 genes) revealed that chromatophores underwent a drastic genome shrinkage. Chromatophores contained genes that were accountable for photosynthesis but were deficient in genes that could carry out other biosynthetic functions; this observation suggests that these endosymbiotic cells are highly dependent on their hosts for their survival and growth mechanisms. Thus, these chromatophores were found to be non-functional for organelle-specific purposes when compared with mitochondria and plastids. This distinction could have promoted the early evolution of photosynthetic organelles. The loss of genetic autonomy, that is, the loss of many genes from endosymbionts, occurred very early in evolutionary time.[26] Taking into account the entire original endosymbiont genome, there are three main possible fates for genes over evolutionary time. The first fate involves the loss of functionally redundant genes,[26] in which genes that are already represented in the nucleus are eventually lost. The second fate involves the transfer of genes to the nucleus.[22][26][27][28][29] The loss of autonomy and integration of the endosymbiont with its host can be primarily attributed to nuclear gene transfer.[29] As organelle genomes have been greatly reduced over evolutionary time, nuclear genes have expanded and become more complex.[22] As a result, many plastid and mitochondrial processes are driven by nuclear encoded gene products.[22] In addition, many nuclear genes originating from endosymbionts have acquired novel functions unrelated to their organelles.[22][29] The mechanisms of gene transfer are not fully known; however, multiple hypotheses exist to explain this phenomenon. The cDNA hypothesis involves the use of messenger RNA (mRNAs) to transport genes from organelles to the nucleus where they are converted to cDNA and incorporated into the genome.[22][27] The cDNA hypothesis is based on studies of the genomes of flowering plants. Protein coding RNAs in mitochondria are spliced and edited using organelle-specific splice and editing sites. Nuclear copies of some mitochondrial genes, however, do not contain organelle-specific splice sites, suggesting a processed mRNA intermediate. The cDNA hypothesis has since been revised as edited mitochondrial cDNAs are unlikely to recombine with the nuclear genome and are more likely to recombine with their native mitochondrial genome. If the edited mitochondrial sequence recombines with the mitochondrial genome, mitochondrial splice sites would no longer exist in the mitochondrial genome. Any subsequent nuclear gene transfer would therefore also lack mitochondrial splice sites.[22] The bulk flow hypothesis is the alternative to the cDNA hypothesis, stating that escaped DNA, rather than mRNA, is the mechanism of gene transfer.[22][27] According to this hypothesis, disturbances to organelles, including autophagy (normal cell destruction), gametogenesis (the formation of gametes), and cell stress, release DNA which is imported into the nucleus and incorporated into the nuclear DNA using non-homologous end joining (repair of double stranded breaks).[27] For example, in the initial stages of endosymbiosis, due to a lack of major gene transfer, the host cell had little to no control over the endosymbiont. The endosymbiont underwent cell division independently of the host cell, resulting in many "copies" of the endosymbiont within the host cell. Some of the endosymbionts lysed (burst), and high levels of DNA were incorporated into the nucleus. A similar mechanism is thought to occur in tobacco plants, which show a high rate of gene transfer and whose cells contain multiple chloroplasts.[26] In addition, the bulk flow hypothesis is also supported by the presence of non-random clusters of organelle genes, suggesting the simultaneous movement of multiple genes.[27] Molecular and biochemical evidence suggests that mitochondria are related to Rickettsiales proteobacteria (in particular, the SAR11 clade,[30][31] or close relatives), and that chloroplasts are related to nitrogen-fixing filamentous cyanobacteria.[32][33]
Endosymbiosis of Protomitochondria
Endosymbiotic theory for the origin of mitochondria suggests that the proto-eukaryote engulfed a protomitochondria, and this endosymbiont became an organelle.[34]
Mitochondria

Mitochondria are organelles that synthesize ATP for the cell by metabolizing carbon-based macromolecules.[35] The presence of deoxyribonucleic acid (DNA) in mitochondria and proteins, derived from mtDNA, suggest that this organelle may have been a prokaryote prior to its integration into the proto-eukaryote.[36] Mitochondria are regarded as organelles rather than endosymbionts because mitochondria and the host cells share some parts of their genome, undergo mitosis simultaneously, and provide each other means to produce energy.[36] Endomembrane system and nuclear membrane were hypothesized to have derived from the protomitochondria.[37][38][39]
Nuclear Membrane
The presence of a nucleus is one major difference between eukaryotes and prokaryotes.[40] Some conserved nuclear proteins between eukaryotes and prokaryotes suggest that these two types had a common ancestor.[41] Another theory behind nucleation is that early nuclear membrane proteins caused the cell membrane to fold inwardly and form a sphere with pores like the nuclear envelope.[42] Strictly regarding energy expenditure, endosymbiosis would save the cell more energy to develop a nuclear membrane than if the cell was to fold its cell membrane to develop this structure since the interactions between proteins are usually enabled by ATP.[38] Digesting engulfed cells without a complex metabolic system that produces massive amounts of energy like mitochondria would have been challenging for the host cell.[37] This theory suggests that the vesicles leaving the protomitochondria may have formed the nuclear envelope.[37]
The process of symbiogenesis by which the early eukaryotic cell integrated the proto-mitochondrion likely included protection of the archaeal host genome from the release of reactive oxygen species (ROS). ROS would have been formed during oxidative phosphorylation and ATP production by the proto-mitochondrion. The nuclear membrane may have evolved as an adaptive innovation for protecting against nuclear genome DNA damage caused by such ROS.[43] Substantial transfer of genes from the ancestral proto-mitochondrial genome to the nuclear genome likely occurred during early eukaryotic evolution.[44] The greater protection of the nuclear genome against ROS afforded by the nuclear membrane may explain the adaptive benefit of this gene transfer.
Endomembrane system
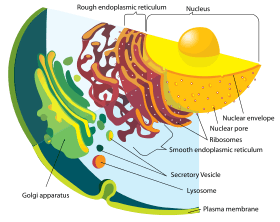
Modern eukaryotic cells use the endomembrane system to transport products and wastes in, within, and out of cells.[45] The membrane of nuclear envelope and endomembrane vesicles are composed of similar membrane proteins.[46] These vesicles also share similar membrane proteins with the organelle they originated from or are traveling towards.[47] This suggests that what formed the nuclear membrane also formed the endomembrane system. Prokaryotes do not have a complex internal membrane network like the modern eukaryotes, but the prokaryotes could produce extracellular vesicles from their outer membrane.[37] After the early prokaryote was consumed by a proto-eukaryote, the prokaryote would have continued to produce vesicles that accumulated within the cell.[37] Interaction of internal components of vesicles may have led to formation of the endoplasmic reticulum and contributed to the formation of Golgi apparatus.[37]
Organellar genomes
Plastomes and mitogenomes

The third and final possible fate of endosymbiont genes is that they remain in the organelles. Plastids and mitochondria, although they have lost much of their genomes, retain genes encoding rRNAs, tRNAs, proteins involved in redox reactions, and proteins required for transcription, translation, and replication.[22][23][26] There are many hypotheses to explain why organelles retain a small portion of their genome; however no one hypothesis will apply to all organisms[26] and the topic is still quite controversial.[22] The hydrophobicity hypothesis states that highly hydrophobic (water hating) proteins (such as the membrane bound proteins involved in redox reactions) are not easily transported through the cytosol and therefore these proteins must be encoded in their respective organelles.[22][26] The code disparity hypothesis states that the limit on transfer is due to differing genetic codes and RNA editing between the organelle and the nucleus.[26] The redox control hypothesis states that genes encoding redox reaction proteins are retained in order to effectively couple the need for repair and the synthesis of these proteins.[22][23][26] For example, if one of the photosystems is lost from the plastid, the intermediate electron carriers may lose or gain too many electrons, signalling the need for repair of a photosystem.[23] The time delay involved in signalling the nucleus and transporting a cytosolic protein to the organelle results in the production of damaging reactive oxygen species.[22][23][26] The final hypothesis states that the assembly of membrane proteins, particularly those involved in redox reactions, requires coordinated synthesis and assembly of subunits; however, translation and protein transport coordination is more difficult to control in the cytoplasm.[26]
Non-photosynthetic plastid genomes
The majority of the genes in the mitochondria and plastids are related to the expression (transcription, translation and replication) of genes encoding proteins involved in either photosynthesis (in plastids) or cellular respiration (in mitochondria).[22][23][26] One might predict that the loss of photosynthesis or cellular respiration would allow for the complete loss of the plastid genome or the mitochondrial genome respectively.[26] While there are numerous examples of mitochondrial descendants (mitosomes and hydrogenosomes) that have lost their entire organellar genome,[47] non-photosynthetic plastids tend to retain a small genome.[26] There are two main hypotheses to explain this occurrence: The essential tRNA hypothesis notes that there have been no documented functional plastid-to-nucleus gene transfers of genes encoding RNA products (tRNAs and rRNAs). As a result, plastids must make their own functional RNAs or import nuclear counterparts. The genes encoding tRNA-Glu and tRNA-fmet, however, appear to be indispensable. The plastid is responsible for haem biosynthesis, which requires plastid encoded tRNA-Glu (from the gene trnE) as a precursor molecule. Like other genes encoding RNAs, trnE cannot be transferred to the nucleus. In addition, it is unlikely trnE could be replaced by a cytosolic tRNA-Glu as trnE is highly conserved; single base changes in trnE have resulted in the loss of haem synthesis. The gene for tRNA-formylmethionine (tRNA-fmet) is also encoded in the plastid genome and is required for translation initiation in both plastids and mitochondria. A plastid is required to continue expressing the gene for tRNA-fmet so long as the mitochondrion is translating proteins.[26] The limited window hypothesis offers a more general explanation for the retention of genes in non-photosynthetic plastids.[48] According to the bulk flow hypothesis, genes are transferred to the nucleus following the disturbance of organelles.[27] Disturbance was common in the early stages of endosymbiosis, however, once the host cell gained control of organelle division, eukaryotes could evolve to have only one plastid per cell. Having only one plastid severely limits gene transfer[26] as the lysis of the single plastid would likely result in cell death.[26][48] Consistent with this hypothesis, organisms with multiple plastids show an 80-fold increase in plastid-to-nucleus gene transfer compared with organisms with single plastids.[48]
Evidence
There are many lines of evidence that mitochondria and plastids including chloroplasts arose from bacteria.[49][50][51][52][53]
- New mitochondria and plastids are formed only through binary fission, the form of cell division used by bacteria and archaea.[54]
- If a cell's mitochondria or chloroplasts are removed, the cell does not have the means to create new ones.[55] For example, in some algae, such as Euglena, the plastids can be destroyed by certain chemicals or prolonged absence of light without otherwise affecting the cell. In such a case, the plastids will not regenerate.
- Transport proteins called porins are found in the outer membranes of mitochondria and chloroplasts and are also found in bacterial cell membranes.[56][57][58]
- A membrane lipid cardiolipin is exclusively found in the inner mitochondrial membrane and bacterial cell membranes.[59]
- Some mitochondria and some plastids contain single circular DNA molecules that are similar to the DNA of bacteria both in size and structure.[60]
- Genome comparisons suggest a close relationship between mitochondria and Rickettsial bacteria.[61]
- Genome comparisons suggest a close relationship between plastids and cyanobacteria.[62]
- Many genes in the genomes of mitochondria and chloroplasts have been lost or transferred to the nucleus of the host cell. Consequently, the chromosomes of many eukaryotes contain genes that originated from the genomes of mitochondria and plastids.[60]
- Mitochondrial and plastid ribosomes are more similar to those of bacteria (70S) than those of eukaryotes.[63]
- Proteins created by mitochondria and chloroplasts use N-formylmethionine as the initiating amino acid, as do proteins created by bacteria but not proteins created by eukaryotic nuclear genes or archaea.[64][65]
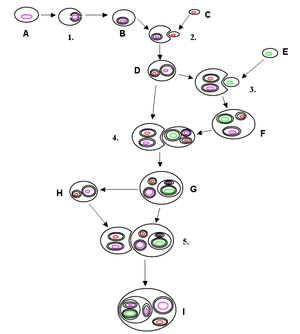
Secondary endosymbiosis
Primary endosymbiosis involves the engulfment of a cell by another free living organism. Secondary endosymbiosis occurs when the product of primary endosymbiosis is itself engulfed and retained by another free living eukaryote. Secondary endosymbiosis has occurred several times and has given rise to extremely diverse groups of algae and other eukaryotes. Some organisms can take opportunistic advantage of a similar process, where they engulf an alga and use the products of its photosynthesis, but once the prey item dies (or is lost) the host returns to a free living state. Obligate secondary endosymbionts become dependent on their organelles and are unable to survive in their absence.[66] RedToL, the Red Algal Tree of Life Initiative funded by the National Science Foundation highlights the role red algae or Rhodophyta played in the evolution of our planet through secondary endosymbiosis. One possible secondary endosymbiosis in process has been observed by Okamoto & Inouye (2005). The heterotrophic protist Hatena behaves like a predator until it ingests a green alga, which loses its flagella and cytoskeleton, while Hatena, now a host, switches to photosynthetic nutrition, gains the ability to move towards light and loses its feeding apparatus.[67] The process of secondary endosymbiosis left its evolutionary signature within the unique topography of plastid membranes. Secondary plastids are surrounded by three (in euglenophytes and some dinoflagellates) or four membranes (in haptophytes, heterokonts, cryptophytes, and chlorarachniophytes). The two additional membranes are thought to correspond to the plasma membrane of the engulfed alga and the phagosomal membrane of the host cell. The endosymbiotic acquisition of a eukaryote cell is represented in the cryptophytes; where the remnant nucleus of the red algal symbiont (the nucleomorph) is present between the two inner and two outer plastid membranes. Despite the diversity of organisms containing plastids, the morphology, biochemistry, genomic organisation, and molecular phylogeny of plastid RNAs and proteins suggest a single origin of all extant plastids – although this theory is still debated.[68][69] Some species including Pediculus humanus (lice) have multiple chromosomes in the mitochondrion. This and the phylogenetics of the genes encoded within the mitochondrion suggest that mitochondria have multiple ancestors, that these were acquired by endosymbiosis on several occasions rather than just once, and that there have been extensive mergers and rearrangements of genes on the several original mitochondrial chromosomes.[70]
Date
The question of when the transition from prokaryotic to eukaryotic form occurred and when the first crown group eukaryotes appeared on earth is still unresolved. The oldest known body fossils that can be positively assigned to the Eukaryota are acanthomorphic acritarchs from the 1631±1 Ma Deonar Formation (lower Vindhyan Supergroup) of India.[71] These fossils can still be identified as derived post-nuclear eukaryotes with a sophisticated, morphology-generating cytoskeleton sustained by mitochondria.[72] This fossil evidence indicates that endosymbiotic acquisition of alphaproteobacteria must have occurred before 1.6 Ga. Molecular clocks have also been used to estimate the last eukaryotic common ancestor (LECA), however these methods have large inherent uncertainty and give a wide range of dates. Reasonable results for LECA include the estimate of c. 1800 Mya.[73] A 2300 Mya estimate[74] also seems reasonable and has the added attraction of coinciding with one of the most pronounced biogeochemical perturbations in Earth history (the Great Oxygenation Event). The marked increase in atmospheric oxygen concentrations during the early Palaeoproterozoic Great Oxidation Event has been invoked as a contributing cause of eukaryogenesis – by inducing the evolution of oxygen-detoxifying mitochondria.[75] Alternatively, the Great Oxidation Event might be a consequence of eukaryogenesis and its impact on the export and burial of organic carbon.[76]
See also
- Angomonas deanei, a protozoan that harbours an obligate bacterial symbiont
- Hatena arenicola, a species that appears to be in the process of acquiring an endosymbiont
- Hydrogen hypothesis
- James A. Lake
- Kleptoplasty
- Mixotricha paradoxa, which itself is a symbiont, contains endosymbiotic bacteria
- Numt, abbreviation of "nuclear mitochondrial DNA"
- Parasite Eve, fiction about endosymbiosis
- Protocell
- Viral eukaryogenesis, hypothesis that the cell nucleus originated from endosymbiosis
References
- Edited by Athel Cornish-Bowden (December 2017). "The origin of mitosing cells: 50th anniversary of a classic paper by Lynn Sagan (Margulis)". Journal of Theoretical Biology. 343: 1–114.CS1 maint: extra text: authors list (link)
- "Mereschkowsky's Tree of Life". Scientific American. Retrieved 1 May 2017.
- Mereschkowski K (15 September 1905). "Über Natur und Ursprung der Chromatophoren im Pflanzenreiche" [On the nature and origin of chromatophores in the plant kingdom]. Biologisches Centralblatt (in German). 25 (18): 593–604.
- See:
- Mereschkowski, Konstantin (15 April 1910). "Theorie der zwei Plasmaarten als Grundlage der Symbiogenesis, einer neuen Lehre von der Entstehung der Organismen" [Theory of two types of plasms as the basis of symbiogenesis, a new study of the origin of organisms [part 1 of 4]]. Biologisches Centralblatt (in German). 30 (8): 278–288.
- Mereschkowsky, Konstantin (1 May 1910). "Theorie der zwei Plasmaarten als Grundlage der Symbiogenesis, einer neuen Lehre von der Entstehung der Organismen" [Theory of two types of plasms as the basis of symbiogenesis, a new study of the origin of organisms [part 2 of 4]]. Biologisches Centralblatt (in German). 30 (9): 289–303.
- Mereschkowski, Konstantin (15 May 1910). "Theorie der zwei Plasmaarten als Grundlage der Symbiogenesis, einer neuen Lehre von der Entstehung der Organismen" [Theory of two types of plasms as the basis of symbiogenesis, a new study of the origin of organisms [part 3 of 4]]. Biologisches Centralblatt (in German). 30 (10): 321–347.
- Mereschkowsky, Konstantin (1 June 1910). "Theorie der zwei Plasmaarten als Grundlage der Symbiogenesis, einer neuen Lehre von der Entstehung der Organismen" [Theory of two types of plasms as the basis of symbiogenesis, a new study of the origin of organisms [part 4 of 4]]. Biologisches Centralblatt (in German). 30 (11): 353–367.
- Martin W, Roettger M, Kloesges T, Thiergart T, Woehle C, Gould S, Dagan T. "Modern endosymbiotic theory: Getting lateral gene transfer into the equation" (PDF). Journal of Endocytobiosis and Cell Research. 23: 1–5.(journal URL: )
- See:
- Schimper, A. F. W. (16 February 1883). "Ueber die Entwicklung der Chlorophyllkörner und Farbkörper" [On the development of chlorophyll granules and colored bodies [part 1 of 4]]. Botanische Zeitung (in German). 41 (7): 105–114. From p. 105: "Inzwischen theilte mir Herr Professor Schmitz mit, dass … die höheren Pflanzen sich ebenso verhalten würden." (Meanwhile, Prof. Schmitz reported to me that among algae, the creation of chlorophyll granules from the cell plasm doesn't occur, but that they arise exclusively from one another by division. The spores receive from the mother plant chlorophyll granules, which create, by division, all of the chlorophyll granules of the plants that arises from them [i.e., the spores]. This finding in algae made it seem likely to Prof. Schmitz that the higher plants would behave likewise.) From p. 106: "Meine Untersuchungen haben ergeben, … aus dem Scheitelmeristem sich entwickelnden Gewebe erzeugen." (My investigations have revealed that the vegetation points [i.e., points of vegetative growth] always contain differentiated chlorophyll bodies or their colorless rudiments; that they arise not by creation from the cell plasm, but from one another by division, and that they create all chlorophyll bodies and starch-forming [bodies] of the tissues developing from the apical meristem.) From p. 112, footnote 2: "Sollte es sich definitiv bestätigen, … an eine Symbiose erinnern." (If it should definitely be confirmed that the plastids in egg cells are not formed anew, then their relation to the organism containing them would somewhat suggest a symbiosis.)
- Schimper, A. F. W. (23 February 1883). "Ueber die Entwicklung der Chlorophyllkörner und Farbkörper" [On the development of chlorophyll granules and colored bodies [part 2 of 4]]. Botanische Zeitung (in German). 41 (8): 121–131.
- Schimper, A. F. W. (2 March 1883). "Ueber die Entwicklung der Chlorophyllkörner und Farbkörper" [On the development of chlorophyll granules and colored bodies [part 3 of 4]]. Botanische Zeitung (in German). 41 (9): 137–146.
- Schimper, A. F. W. (9 March 1883). "Ueber die Entwicklung der Chlorophyllkörner und Farbkörper" [On the development of chlorophyll granules and colored bodies [part 4 of 4]]. Botanische Zeitung (in German). 41 (10): 153–162.
- Portier, Paul (1918). Les Symbiotes (in French). Paris, France: Masson et Cie. p. 293. From p. 293: "Cette modification dans les rapports des appareils nucléaire et mitochondrial peut être le résultat de deux mécanismes. … Cette la parthénogénèse." (This modification in the relations of the nuclear and mitochondrial systems could be the result of two mechanisms: (a) There is a combination of two factors: contribution of new symbionts by the spermatozoid and reduction division. That is fertilization. (b) A single factor exists: reduction division: in this case, the egg contains sufficiently active symbionts. That is parthenogenesis.)
- Lane, Nick (2005). Power, Sex, Suicide. Mitochondria and the Meaning of Life. New York: Oxford University Press. p. 14. ISBN 9780199205646.
- Wallin IE (1923). "The Mitochondria Problem". The American Naturalist. 57 (650): 255–61. doi:10.1086/279919.
- Wallin, I. E. (1927). Symbionticism and the origin of species. Baltimore: Williams & Wilkins Company. p. 117.
- Margulis, Lynn (2011). "Symbiogenesis. A new principle of evolution rediscovery of Boris Mikhaylovich Kozo-Polyansky (1890–1957)". Paleontological Journal. 44 (12): 1525–1539. doi:10.1134/S0031030110120087.
- Козо-полянский (Kozo-Polyansky), Б. (B.) (1924). Новый принцип биологии. Очерк теории симбиогенеза [A New Principle of Biology. Essay on the Theory of Symbiogenesis] (in Russian). Moscow and Leningrad (St. Petersburg), Russia: Пучина (Puchina).
- English translation: Kozo-Polyansky, Boris Mikhaylovich; Fet, Victor (trans.); Margulis, Lynn (ed.) (2010). Symbiogenesis: A New Principle of Evolution. Cambridge, Massachusetts, USA: Harvard University Press.CS1 maint: extra text: authors list (link)
- Reviewed in: Niklas, Karl J. (2010). "Boris M. Kozo-Polyansky, Symbiogenesis: A New Principle of Evolution". Symbiosis. 52 (1): 49–50. doi:10.1007/s13199-010-0098-7.
- Corning, Peter A. (2010). Holistic Darwinism: Synergy, Cybernetics, and the Bioeconomics of Evolution. Chicago: University of Chicago Press. p. 81. ISBN 978-0-22611-633-4.
- Ris H, Plaut W (June 1962). "Ultrastructure of DNA-containing areas in the chloroplast of Chlamydomonas". The Journal of Cell Biology. 13 (3): 383–91. doi:10.1083/jcb.13.3.383. PMC 2106071. PMID 14492436.
- Ris H, Singh RN (January 1961). "Electron microscope studies on blue-green algae". The Journal of Biophysical and Biochemical Cytology. 9 (1): 63–80. doi:10.1083/jcb.9.1.63. PMC 2224983. PMID 13741827.
- Stocking CR, Gifford Jr EM (1959). "Incorporation of thymidine into chloroplasts of Spirogyra". Biochem. Biophys. Res. Commun. 1 (3): 159–64. doi:10.1016/0006-291X(59)90010-5.
- Sagan L (March 1967). "On the origin of mitosing cells". Journal of Theoretical Biology. 14 (3): 255–74. doi:10.1016/0022-5193(67)90079-3. PMID 11541392.
- Margulis L, Sagan D (2001). "Marvellous microbes". Resurgence. 206: 10–12.
- Gabaldón T, Snel B, van Zimmeren F, Hemrika W, Tabak H, Huynen MA (March 2006). "Origin and evolution of the peroxisomal proteome". Biology Direct. 1 (1): 8. doi:10.1186/1745-6150-1-8. PMC 1472686. PMID 16556314. (Provides evidence that contradicts an endosymbiotic origin of peroxisomes, and suggests instead that they originate evolutionarily from the endoplasmic reticulum)
- Keeling PJ, Archibald JM (April 2008). "Organelle evolution: what's in a name?". Current Biology. 18 (8): R345-7. doi:10.1016/j.cub.2008.02.065. PMID 18430636.
- Michael Syvanen, Clarence I. Kado Horizontal Gene Transfer Academic Press, p. 405 ISBN 978-0126801262
- Timmis JN, Ayliffe MA, Huang CY, Martin W (February 2004). "Endosymbiotic gene transfer: organelle genomes forge eukaryotic chromosomes". Nature Reviews. Genetics. 5 (2): 123–35. doi:10.1038/nrg1271. PMID 14735123.
- Koumandou VL, Nisbet RE, Barbrook AC, Howe CJ (May 2004). "Dinoflagellate chloroplasts--where have all the genes gone?". Trends in Genetics. 20 (5): 261–7. doi:10.1016/j.tig.2004.03.008. PMID 15109781.
- Taanman JW (February 1999). "The mitochondrial genome: structure, transcription, translation and replication". Biochimica et Biophysica Acta (BBA) - Bioenergetics. 1410 (2): 103–23. doi:10.1016/S0005-2728(98)00161-3. PMID 10076021.
- Nowack EC, Melkonian M, Glöckner G (March 2008). "Chromatophore genome sequence of Paulinella sheds light on acquisition of photosynthesis by eukaryotes". Current Biology. 18 (6): 410–8. doi:10.1016/j.cub.2008.02.051. PMID 18356055.
- Barbrook AC, Howe CJ, Purton S (February 2006). "Why are plastid genomes retained in non-photosynthetic organisms?". Trends in Plant Science. 11 (2): 101–8. doi:10.1016/j.tplants.2005.12.004. PMID 16406301.
- Leister D (December 2005). "Origin, evolution and genetic effects of nuclear insertions of organelle DNA". Trends in Genetics. 21 (12): 655–63. doi:10.1016/j.tig.2005.09.004. hdl:11858/00-001M-0000-0012-3B56-7. PMID 16216380.
- Keeling PJ (October 2004). "Diversity and evolutionary history of plastids and their hosts". American Journal of Botany. 91 (10): 1481–93. doi:10.3732/ajb.91.10.1481. PMID 21652304.
- Archibald JM (January 2009). "The puzzle of plastid evolution". Current Biology. 19 (2): R81-8. doi:10.1016/j.cub.2008.11.067. PMID 19174147.
- "Mitochondria Share an Ancestor With SAR11, a Globally Significant Marine Microbe". ScienceDaily. July 25, 2011. Retrieved 2011-07-26.
- Thrash JC, Boyd A, Huggett MJ, Grote J, Carini P, Yoder RJ, et al. (2011). "Phylogenomic evidence for a common ancestor of mitochondria and the SAR11 clade". Scientific Reports. 1: 13. Bibcode:2011NatSR...1E..13T. doi:10.1038/srep00013. PMC 3216501. PMID 22355532.
- Deusch O, Landan G, Roettger M, Gruenheit N, Kowallik KV, Allen JF, et al. (April 2008). "Genes of cyanobacterial origin in plant nuclear genomes point to a heterocyst-forming plastid ancestor". Molecular Biology and Evolution. 25 (4): 748–61. doi:10.1093/molbev/msn022. PMID 18222943.
- Ochoa de Alda JA, Esteban R, Diago ML, Houmard J (September 2014). "The plastid ancestor originated among one of the major cyanobacterial lineages". Nature Communications. 5: 4937. Bibcode:2014NatCo...5.4937O. doi:10.1038/ncomms5937. PMID 25222494.
- Zimorski V, Ku C, Martin WF, Gould SB (December 2014). "Endosymbiotic theory for organelle origins". Current Opinion in Microbiology. 22: 38–48. doi:10.1016/j.mib.2014.09.008. PMID 25306530.
- "Mitochondria, Cell Energy, ATP Synthase | Learn Science at Scitable". www.nature.com. Retrieved 2019-03-24.
- Gruber A (January 2019). "What's in a name? How organelles of endosymbiotic origin can be distinguished from endosymbionts". Microbial Cell. 6 (2): 123–133. doi:10.15698/mic2019.02.668. PMC 6364258. PMID 30740457.
- Gould SB, Garg SG, Martin WF (July 2016). "Bacterial Vesicle Secretion and the Evolutionary Origin of the Eukaryotic Endomembrane System". Trends in Microbiology. 24 (7): 525–534. doi:10.1016/j.tim.2016.03.005. PMID 27040918.
- Martin WF, Garg S, Zimorski V (September 2015). "Endosymbiotic theories for eukaryote origin". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 370 (1678): 20140330. doi:10.1098/rstb.2014.0330. PMC 4571569. PMID 26323761.
- Garavís M, González C, Villasante A (June 2013). "On the origin of the eukaryotic chromosome: the role of noncanonical DNA structures in telomere evolution". Genome Biology and Evolution. 5 (6): 1142–50. doi:10.1093/gbe/evt079. PMC 3698924. PMID 23699225.
- "Typical prokaryotic (left) and eukaryotic (right) cells | Learn Science at Scitable". www.nature.com. Retrieved 2019-03-24.
- Devos DP, Gräf R, Field MC (June 2014). "Evolution of the nucleus". Current Opinion in Cell Biology. 28: 8–15. doi:10.1016/j.ceb.2014.01.004. PMC 4071446. PMID 24508984.
- Wilson KL, Dawson SC (October 2011). "Evolution: functional evolution of nuclear structure". The Journal of Cell Biology. 195 (2): 171–81. doi:10.1083/jcb.201103171. PMC 3198171. PMID 22006947.
- Bernstein H, Bernstein C (2017). "Sexual communication in archaea, the precursor to meiosis.". In Witzany G (ed.). Biocommunication of Archaea. Springer International Publishing. pp. 103–117. doi:10.1007/978-3-319-65536-9. ISBN 978-3-319-65535-2.
- Gabaldón T, Huynen MA (August 2003). "Reconstruction of the proto-mitochondrial metabolism". Science. 301 (5633): 609. doi:10.1126/science.1085463. PMID 12893934.
- "Endomembrane System - an overview | ScienceDirect Topics". www.sciencedirect.com. Retrieved 2019-03-24.
- Liashkovich I, Shahin V (August 2017). "Functional implication of the common evolutionary origin of nuclear pore complex and endomembrane management systems". Seminars in Cell & Developmental Biology. 68: 10–17. doi:10.1016/j.semcdb.2017.04.006. PMID 28473267.
- Howe CJ (May 2008). "Cellular evolution: what's in a mitochondrion?". Current Biology. 18 (10): R429–R431. doi:10.1016/j.cub.2008.04.007. PMID 18492476.
- Lane N (2011). "Plastids, genomes, and the probability of gene transfer". Genome Biology and Evolution. 3: 372–4. doi:10.1093/gbe/evr003. PMC 3101016. PMID 21292628.
- Kimball, J. 2010. Kimball's Biology Pages. Accessed October 13, 2010. An online open source biology text by Harvard professor, and author of a general biology text, John W. Kimball.
- Reece, J., Lisa A. Urry, Michael L. Cain, Steven A. Wasserman, Peter V. Minorsky, Robert B. Jackson, 2010. Campbell Biology. 9th Edition Benjamin Cummings; 9th Ed. (October 7, 2010)
- Raven P, Johnson G, Mason K, Losos J, Singer S (January 14, 2010). Biology (9th ed.). McGraw-Hill.
- Gray, MW (1992). The endosymbiont hypothesis revisited. International Review of Cytology. 141. pp. 233–357. doi:10.1016/S0074-7696(08)62068-9. ISBN 9780123645449. PMID 1452433.
- Zimorski V, Ku C, Martin WF, Gould SB (December 2014). "Endosymbiotic theory for organelle origins". Current Opinion in Microbiology. 22: 38–48. doi:10.1016/j.mib.2014.09.008. PMID 25306530.
- Margolin W (November 2005). "FtsZ and the division of prokaryotic cells and organelles". Nature Reviews. Molecular Cell Biology. 6 (11): 862–71. doi:10.1038/nrm1745. PMC 4757588. PMID 16227976.
- Wise RR, Hoober JK (2007). Structure and function of plastids. Berlin: Springer. p. 104. ISBN 9781402065705.
- Fischer K, Weber A, Brink S, Arbinger B, Schünemann D, Borchert S, et al. (October 1994). "Porins from plants. Molecular cloning and functional characterization of two new members of the porin family". The Journal of Biological Chemistry. 269 (41): 25754–60. PMID 7523392.
- Zeth K, Thein M (October 2010). "Porins in prokaryotes and eukaryotes: common themes and variations". The Biochemical Journal. 431 (1): 13–22. doi:10.1042/BJ20100371. PMID 20836765.
- Fairman JW, Noinaj N, Buchanan SK (August 2011). "The structural biology of β-barrel membrane proteins: a summary of recent reports". Current Opinion in Structural Biology. 21 (4): 523–31. doi:10.1016/j.sbi.2011.05.005. PMC 3164749. PMID 21719274.
- Mileykovskaya E, Dowhan W (October 2009). "Cardiolipin membrane domains in prokaryotes and eukaryotes". Biochimica et Biophysica Acta (BBA) - Biomembranes. 1788 (10): 2084–91. doi:10.1016/j.bbamem.2009.04.003. PMC 2757463. PMID 19371718.
- Timmis JN, Ayliffe MA, Huang CY, Martin W (February 2004). "Endosymbiotic gene transfer: organelle genomes forge eukaryotic chromosomes". Nature Reviews. Genetics. 5 (2): 123–35. doi:10.1038/nrg1271. PMID 14735123.
- Andersson SG, Zomorodipour A, Andersson JO, Sicheritz-Pontén T, Alsmark UC, Podowski RM, et al. (November 1998). "The genome sequence of Rickettsia prowazekii and the origin of mitochondria". Nature. 396 (6707): 133–40. Bibcode:1998Natur.396..133A. doi:10.1038/24094. PMID 9823893.
- Dagan T, Roettger M, Stucken K, Landan G, Koch R, Major P, et al. (2013). "Genomes of Stigonematalean cyanobacteria (subsection V) and the evolution of oxygenic photosynthesis from prokaryotes to plastids". Genome Biology and Evolution. 5 (1): 31–44. doi:10.1093/gbe/evs117. PMC 3595030. PMID 23221676.
- Manuell AL, Quispe J, Mayfield SP (August 2007). "Structure of the chloroplast ribosome: novel domains for translation regulation". PLOS Biology. 5 (8): e209. doi:10.1371/journal.pbio.0050209. PMC 1939882. PMID 17683199.
- Schwartz JH, Meyer R, Eisenstadt JM, Brawerman G (May 1967). "Involvement of N-formylmethionine in initiation of protein synthesis in cell-free extracts of Euglena gracilis". Journal of Molecular Biology. 25 (3): 571–4. doi:10.1016/0022-2836(67)90210-0. PMID 5340700.
- Smith AE, Marcker KA (December 1968). "N-formylmethionyl transfer RNA in mitochondria from yeast and rat liver". Journal of Molecular Biology. 38 (2): 241–3. doi:10.1016/0022-2836(68)90409-9. PMID 5760639.
- McFadden GI (2001). "Primary and secondary endosymbiosis and the origin of plastids". Journal of Phycology. 37 (6): 951–959. doi:10.1046/j.1529-8817.2001.01126.x.
- Okamoto N, Inouye I (October 2005). "A secondary symbiosis in progress?". Science. 310 (5746): 287. doi:10.1126/science.1116125. PMID 16224014.
- McFadden GI, van Dooren GG (July 2004). "Evolution: red algal genome affirms a common origin of all plastids". Current Biology. 14 (13): R514-6. doi:10.1016/j.cub.2004.06.041. PMID 15242632.
- Gould SB, Waller RF, McFadden GI (2008). "Plastid evolution". Annual Review of Plant Biology. 59 (1): 491–517. doi:10.1146/annurev.arplant.59.032607.092915. PMID 18315522.
- Georgiades K, Raoult D (October 2011). "The rhizome of Reclinomonas americana, Homo sapiens, Pediculus humanus and Saccharomyces cerevisiae mitochondria". Biology Direct. 6: 55. doi:10.1186/1745-6150-6-55. PMC 3214132. PMID 22014084.
- Prasad, Pijai (August 2005). "Organic-walled microfossils from the Proterozoic Vindhyan Supergroup of Son Valley, Madhya Pradesh, India" (PDF). Paleobotanist. 54.
- Butterfield, Nicholas J. (2014-11-26). "Early evolution of the Eukaryota". Palaeontology. 58 (1): 5–17. doi:10.1111/pala.12139.
- Parfrey LW, Lahr DJ, Knoll AH, Katz LA (August 2011). "Estimating the timing of early eukaryotic diversification with multigene molecular clocks". Proceedings of the National Academy of Sciences of the United States of America. 108 (33): 13624–9. Bibcode:2011PNAS..10813624P. doi:10.1073/pnas.1110633108. PMC 3158185. PMID 21810989.
- Hedges SB, Blair JE, Venturi ML, Shoe JL (January 2004). "A molecular timescale of eukaryote evolution and the rise of complex multicellular life". BMC Evolutionary Biology. 4: 2. doi:10.1186/1471-2148-4-2. PMC 341452. PMID 15005799.
- Gross J, Bhattacharya D (August 2010). "Uniting sex and eukaryote origins in an emerging oxygenic world". Biology Direct. 5: 53. doi:10.1186/1745-6150-5-53. PMC 2933680. PMID 20731852.
- Butterfield, Nicholas J. (1997). "Plankton ecology and the Proterozoic-Phanerozoic transition". Paleobiology. 23 (2): 247–262. doi:10.1017/S009483730001681X.
Further reading
- Alberts B (2002). Molecular Biology of the Cell. New York: Garland Science. ISBN 978-0-8153-3218-3. (General textbook)
- Brinkman FS, Blanchard JL, Cherkasov A, Av-Gay Y, Brunham RC, Fernandez RC, et al. (August 2002). "Evidence that plant-like genes in Chlamydia species reflect an ancestral relationship between Chlamydiaceae, cyanobacteria, and the chloroplast". Genome Research. 12 (8): 1159–67. doi:10.1101/gr.341802. PMC 186644. PMID 12176923.
- Cohen WE, Gardner RS (1959). "Viral Theory and Endosymbiosis" (PDF). Archived from the original (PDF) on 2011-07-15. Retrieved 2009-08-26. (Discusses theory of origin of eukaryotic cells by incorporating mitochondria and chloroplasts into anaerobic cells with emphasis on 'phage bacterial and putative viral mitochondrial/chloroplast interactions.)
- Jarvis P (April 2001). "Intracellular signalling: the chloroplast talks!". Current Biology. 11 (8): R307-10. doi:10.1016/S0960-9822(01)00171-3. PMID 11369220. (Recounts evidence that chloroplast-encoded proteins affect transcription of nuclear genes, as opposed to the more well-documented cases of nuclear-encoded proteins that affect mitochondria or chloroplasts.)
- Blanchard JL, Lynch M (July 2000). "Organellar genes: why do they end up in the nucleus?". Trends in Genetics. 16 (7): 315–20. doi:10.1016/S0168-9525(00)02053-9. PMID 10858662. (Discusses theories on how mitochondria and chloroplast genes are transferred into the nucleus, and also what steps a gene needs to go through in order to complete this process.)
- Okamoto N, Inouye I (October 2005). "A secondary symbiosis in progress?". Science. 310 (5746): 287. doi:10.1126/science.1116125. PMID 16224014.
- Understanding Science Team. "Cells within cells: An extraordinary claim with extraordinary evidence" (PDF). University of California, Berkeley. Retrieved 16 February 2014.
