Cat gap
The cat gap is a period in the fossil record of approximately 25 to 18.5 million years ago in which there are few fossils of cats or cat-like species found in North America. The cause of the "cat gap" is disputed, but may have been caused by changes in the climate (global cooling), changes in the habitat and environmental ecosystem, the increasingly hypercarnivorous trend of the cats (especially the nimravids), volcanic activity, evolutionary changes in dental morphology of the Canidae species present in North America, or a periodicity of extinctions called van der Hammen cycles.[1]
Cat evolution
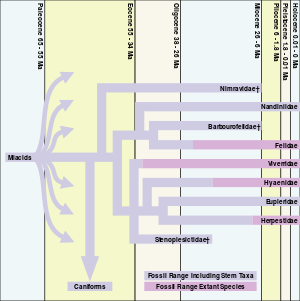
All modern carnivorans, including cats, evolved from miacoids, which existed from approximately 66 to 33 million years ago. There were other earlier cat-like species but Proailurus (meaning "before the cat"; also called "Leman's Dawn Cat"), which appeared about 30 million years ago, is generally considered the first "true cat".[2]
Following the appearance of the dawn cat, there is little in the fossil record for 10 million years to suggest that cats would prosper. In fact, although Proailurus persisted for at least 14 million years, there are so few felid fossils towards the end of the dawn cat's reign that paleontologists refer to this as the "cat gap". The turning point for cats came about with the appearance of a new genus of felids, Pseudaelurus[2]
The increase in disparity through the early Miocene occurs during a time when few feliform fossils have been found in North America. The hypercarnivorous nimravid feliforms were extinct in North America after 26 Ma and felids did not arrive in North America until the Middle Miocene with the appearance of Pseudaelurus. Pseudaelurus crossed over to North America by way of the Bering land bridge from surviving populations in Asia 18.5 million years ago. All modern-day cats are descended from Pseudaelurus.
Nimravids and barbourofelids were saber-toothed cat-like animals of the families Nimravidae and Barbourofelidae, respectively. Although not "true cats" of the family Felidae, they are closely related to felids. The Nimravidae are either basal feliforms or a sister group to both feliforms and caniforms, while the Barbourofelidae are a sister group to the Felidae. Physically, some Nimravidae and Barbourofelidae resembled the saber-toothed cat Smilodon, which would not appear until many millions of years later. Nimravidae also became extinct in North America during the "cat gap".[3]
Possible causes
Hypercarnivorous tendency
The history of carnivorous mammals is characterized by a series of rise-and-fall patterns of diversification, in which declining clades are replaced by phylogenetically distinct but functionally similar clades. Over the past 50 million years, successive clades of small and large carnivorous mammals diversified and then declined to extinction. In most instances, the cause of the decline was energetic constraints and pervasive selection for larger size (Cope's rule) that lead to hypercarnivory dietary specialization. Hypercarnivory leads to increased vulnerability to extinction.
The nimravids were large cat-like animals that occupied this ecomorphic niche in the ecosystem until 26 Ma. It is highly likely that their hypercarnivory led to their extinction in North America. After the extinction of the nimravids there were no other feliform or felid-like species until other felids arrived from Eurasia after crossing the Bering land bridge 18.5 million years ago. During this time there was great diversity among the other carnivorous mammals in North America – both hypocarnivorous and hypercarnivorous species – and other hypercarnivorous species existed before, during, and after the cat gap.
Changes in climate and habitat

Another possible explanation for the extinction of feliforms in North America is changes in the ecology of the continent. Evidence from the geologic temperature record shows that the earth was experiencing a period of global cooling, causing forests to give way to savannas.[2] Climatic changes to arid conditions that muted variation at about 25.8 Ma coincides with the first appearance of hoglike creodonts and of pocket gophers, and this also is the beginning of the "cat gap" and the "entelodont gap", a period of some 7 million years when there were no nimravids, felids, or entelodonts in North America. Faunal overturn at 25.8 Ma is the basis for division of the Arikareean time period (30.5–19 Ma), and the Arikareen NALMA (North American Land-Mammal Ages), into the Monroecreekian period (29.5–25.8 Ma), and then the Harrisonian period (25.8–23.5 Ma).[4]
Why did these cat-like creatures die out in North America (while surviving in Eurasia) with no replacement by the true cats? Their fate may be owed to the same factors that created the diversity of herbivorous mammals, for most cats need forest or cover from which to hunt. In an increasingly open America the nimravids may have found themselves without an ecological perch to hunt from, particularly if the competition with dogs prevented them from colonising the savannas.[5]
Other
Volcanic activity has also been promoted as a possible cause of the cat gap as well as other extinctions during this time period. The La Garita Caldera is a large volcanic caldera located in the San Juan Mountains in southwestern Colorado, United States, and is one of a number of calderas that formed during a massive ignimbrite flare-up in Colorado, Utah, and Nevada during the Oligocene Epoch. The La Garita Caldera was the site of the Fish Canyon eruption, an enormous eruption about 27 million years ago. The scale of the Fish Canyon eruption was far beyond anything known in human history (erupting more than 10,000 km3 or 2,400 cu mi for a VEI 8+ magnitude), and was possibly the most energetic event on Earth since the Chicxulub impact, which is thought by many paleontologists to have caused the extinction of the dinosaurs in the Cretaceous–Paleogene extinction event. The resulting explosive volcanism probably ejected large amounts of dust and debris into the stratosphere causing major cooling (see volcanic winter). Climatic effects could also have been caused by sulphur ejected into the stratosphere, which rapidly converts to sulphuric acid, an aerosol which cools the troposphere by blocking incoming solar radiation.
Another possible cause of the cat gap could have been the Late Cenozoic Ice Age that began 33.9 million years ago. This ice age caused glaciation in Antarctica that eventually spread to Arctic regions of southern Alaska, Greenland, and Iceland. Glaciers on the North American continent, as well as the cooling trend, could have made the ecosystem uninhabitable for feliformia cat-like species, although habitable for cold-weather caniformia species such as canids (dog-like species), mustelids (weasel-like species), and ursids (bear-like species).
There is also evidence that during the Miocene a sill surrounding the Arctic Ocean, known as the Greenland–Scotland Ridge, subsided, allowing more cold polar water to escape into the North Atlantic. As the salinity of the North Atlantic grew and as outflow of cold polar water increased, so the thermohaline circulation increased in vigour, providing the mild winter temperatures and large amounts of moisture to the North Atlantic, which are prerequisites to the build-up of the large continental ice caps on the adjacent cold continents.[6]
Evolution of caniforms during the gap

It has been suggested by some that as a result of the cat gap caniforms (dog-like species including canids, bears, weasels, and other related taxa) evolved to fill more carnivorous and hypercarnivorous ecological niches that would otherwise have been filled by cats.[7] This conclusion, however, is disputed.[8]
During or just prior to this "cat gap," numerous caniform species evolve catlike features indicative of hypercarnivory, such as reduced snouts, somewhat enlarged canines, and fairly extreme reduction of their crushing molars. In North America the first caniform group of moderate body size to move in the direction of hypercarnivory were the endemic hesperocyonine canids, with three genera (Parenhydrocyon, Enhydrocyon, and Mesocyon), ranging in size from jackals to small coyotes, appearing in the early Arikareean (circa 28 MYA). Notably, these three evolved alongside the last hyaenodont and the remaining three nimravids, two of which were puma-sized. The small hypercarnivorous canids were soon joined by and ultimately replaced by numerous species from other families which also had evolved more specialized meat-eating teeth and skulls. These included at least three larger genera of similarly adapted amphicyonids, one endemic (Daphoenodon) and two from the Old World (Temnocyon and Mammocyon), a leopard-sized mustelid (Megalictis) as well as two hypercarnivorous bears, the hemicyonines Cephalogale and Phoberocyon.[7]
However, other paleontologists take issue with this conclusion:
It has been suggested that canids evolved hypercarnivorous morphologies because feliforms were absent during this period (the ‘cat-gap’’, 26–16 Ma). The data presented here do not support this hypothesis. In the calculated morphospace... Canids never occupy the area of morphospace in which felids, nimravids, and hypercarnivorous creodonts are found. More pertinent to the issue at hand, however, is that most of these hypercarnivorous canids were present before the disappearance of the nimravids, and all became extinct before the appearance of felids....There was a progressive and marked decrease in hypercarnivorous forms during the ‘‘cat-gap.’’ 28–20 Ma are characterized by above average extinction intensities and below average origination intensities. 20 Ma was marked by an increase in origination intensity, and 18 Ma showed a decrease in extinction intensity and a large increase in origination intensity. Nonetheless, despite increased origination intensities and decreased extinction intensities near the end of the ‘‘cat-gap’’ (20–16 Ma), there was still no substantial invasion of hypercarnivorous morphospace until the immigration of felids into North America."[8]
References
- T. J. Meehan; L. D. Martin (2003). "Extinction and re-evolution of similar adaptive types (ecomorphs) in Cenozoic North American ungulates and carnivores reflect van der Hammens cycles" (PDF). Naturwissenschaften. 90 (3): 131–135. Bibcode:2003NW.....90..131M. doi:10.1007/s00114-002-0392-1. PMID 12649755. Retrieved 2008-11-28.
- Hunter, Luke; Hinde, Gerald (2006). Cats of Africa: Behavior, Ecology, and Conservation. Cape Town: Johns Hopkins University Press. pp. 40–42. ISBN 978-0-8018-8482-5.
- R. M. Joeckel; Stephane Peigneb; Robert M. Hunt; Robert I. Skolnick (2002). "The Auditory Region and Nasal Cavity of Oligocene Nimravidae". Journal of Vertebrate Paleontology. 22 (4): 131–135. doi:10.1671/0272-4634(2002)022[0830:TARANC]2.0.CO;2. Retrieved 2008-11-28.
- Gregory J. Retallack (2004). "Late Oligocene bunch grassland and early Miocene sod grassland paleosols from central Oregon, USA" (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 207 (3–4): 203–237. Bibcode:2004PPP...207..203R. doi:10.1016/j.palaeo.2003.09.027. Archived from the original (PDF) on 2008-08-30. Retrieved 2008-11-28.
- Flannery, Tim (2002). The Eternal Frontier: An Ecological History of North America and Its Peoples. New York: Grove Press. pp. 113–114. ISBN 0-8021-3888-8.
- Haggart, B. A. (2000). "Ice-age Theories". The Oxford Companion to the Earth. New York: Oxford University Press.
- Van Valkenburgh, Blaire (1999). "Major Patterns in the History of Carnivorous Mammals". Annual Review of Earth and Planetary Sciences. 27 (1): 463–493. Bibcode:1999AREPS..27..463V. doi:10.1146/annurev.earth.27.1.463.
- Gina D. Wesley-Hunt (2005). "The morphological diversification of carnivores in North America". Paleobiology. 31 (1): 35–55. doi:10.1666/0094-8373(2005)031<0035:TMDOCI>2.0.CO;2. Retrieved 2008-11-28.
