Origin of language
The origin of language and its evolutionary emergence in the human species have been subjects of speculation for several centuries. The topic is difficult to study because of the lack of direct evidence. Consequently, scholars wishing to study the origins of language must draw inferences from other kinds of evidence such as the fossil record, archaeological evidence, contemporary language diversity, studies of language acquisition and comparisons between human language and systems of communication existing among animals (particularly other primates). Many argue that the origins of language probably relate closely to the origins of modern human behavior, but there is little agreement about the implications and directionality of this connection.
| Part of a series on |
| Linguistics |
|---|
|
|
This shortage of empirical evidence has caused many scholars to regard the entire topic as unsuitable for serious study. In 1866, the Linguistic Society of Paris banned any existing or future debates on the subject, a prohibition which remained influential across much of the Western world until late in the twentieth century.[1][2] Today, there are various hypotheses about how, why, when, and where language might have emerged.[3] Despite this, there is scarcely more agreement today than a hundred years ago, when Charles Darwin's theory of evolution by natural selection provoked a rash of armchair speculation on the topic.[4] Since the early 1990s, however, a number of linguists, archaeologists, psychologists, anthropologists, and others have attempted to address with new methods what some consider one of the hardest problems in science.[5]
Approaches
One can sub-divide approaches to the origin of language according to some underlying assumptions:[6]
- "Continuity theories" built on the idea that language exhibits so much complexity that one cannot imagine it simply appearing from nothing in its final form; therefore it must have evolved from earlier pre-linguistic systems among our primate ancestors.
- "Discontinuity theories" take the opposite approach—that language, as a unique trait which cannot be compared to anything found among non-humans, must have appeared fairly suddenly during the course of human evolution.
- Some theories consider language mostly as an innate faculty—largely genetically encoded.
- Other theories regard language as a mainly cultural system—learned through social interaction.
Noam Chomsky, a proponent of discontinuity theory, argues that a single chance mutation occurred in one individual in the order of 100,000 years ago, installing the language faculty (a component of the mid-brain) in "perfect" or "near-perfect" form.[7] A majority of linguistic scholars as of 2018 believe continuity-based theories, but they vary in how they hypothesize language development. Among those who consider language as mostly innate, some—notably Steven Pinker[8]—avoid speculating about specific precursors in nonhuman primates, stressing simply that the language faculty must have evolved in the usual gradual way.[9] Others in this intellectual camp—notably Ib Ulbæk[6]—hold that language evolved not from primate communication but from primate cognition, which is significantly more complex.
Those who consider language as learned socially, such as Michael Tomasello, consider it developing from the cognitively controlled aspects of primate communication, these being mostly gestural as opposed to vocal.[10][11] Where vocal precursors are concerned, many continuity theorists envisage language evolving from early human capacities for song.[12][13][14][15][16]
Transcending the continuity-versus-discontinuity divide, some scholars view the emergence of language as the consequence of some kind of social transformation[17] that, by generating unprecedented levels of public trust, liberated a genetic potential for linguistic creativity that had previously lain dormant.[18][19][20] "Ritual/speech coevolution theory" exemplifies this approach.[21][22] Scholars in this intellectual camp point to the fact that even chimpanzees and bonobos have latent symbolic capacities that they rarely—if ever—use in the wild.[23] Objecting to the sudden mutation idea, these authors argue that even if a chance mutation were to install a language organ in an evolving bipedal primate, it would be adaptively useless under all known primate social conditions. A very specific social structure—one capable of upholding unusually high levels of public accountability and trust—must have evolved before or concurrently with language to make reliance on "cheap signals" (words) an evolutionarily stable strategy.
Because the emergence of language lies so far back in human prehistory, the relevant developments have left no direct historical traces; neither can comparable processes be observed today. Despite this, the emergence of new sign languages in modern times—Nicaraguan Sign Language, for example—may potentially offer insights into the developmental stages and creative processes necessarily involved.[24] Another approach inspects early human fossils, looking for traces of physical adaptation to language use.[25][26] In some cases, when the DNA of extinct humans can be recovered, the presence or absence of genes considered to be language-relevant —FOXP2, for example—may prove informative.[27] Another approach, this time archaeological, involves invoking symbolic behavior (such as repeated ritual activity) that may leave an archaeological trace—such as mining and modifying ochre pigments for body-painting—while developing theoretical arguments to justify inferences from symbolism in general to language in particular.[28][29][30]
The time range for the evolution of language and/or its anatomical prerequisites extends, at least in principle, from the phylogenetic divergence of Homo (2.3 to 2.4 million years ago) from Pan (5 to 6 million years ago) to the emergence of full behavioral modernity some 50,000–150,000 years ago. Few dispute that Australopithecus probably lacked vocal communication significantly more sophisticated than that of great apes in general,[31] but scholarly opinions vary as to the developments since the appearance of Homo some 2.5 million years ago. Some scholars assume the development of primitive language-like systems (proto-language) as early as Homo habilis, while others place the development of symbolic communication only with Homo erectus (1.8 million years ago) or with Homo heidelbergensis (0.6 million years ago) and the development of language proper with Homo sapiens, currently estimated at less than 200,000 years ago.
Using statistical methods to estimate the time required to achieve the current spread and diversity in modern languages, Johanna Nichols—a linguist at the University of California, Berkeley—argued in 1998 that vocal languages must have begun diversifying in our species at least 100,000 years ago.[32] A further study by Q. D. Atkinson[13] suggests that successive population bottlenecks occurred as our African ancestors migrated to other areas, leading to a decrease in genetic and phenotypic diversity. Atkinson argues that these bottlenecks also affected culture and language, suggesting that the further away a particular language is from Africa, the fewer phonemes it contains. By way of evidence, Atkinson claims that today's African languages tend to have relatively large numbers of phonemes, whereas languages from areas in Oceania (the last place to which humans migrated), have relatively few. Relying heavily on Atkinson's work, a subsequent study has explored the rate at which phonemes develop naturally, comparing this rate to some of Africa's oldest languages. The results suggest that language first evolved around 50,000–150,000 years ago, which is around the time when modern Homo sapiens evolved.[33] Estimates of this kind are not universally accepted, but jointly considering genetic, archaeological, palaeontological and much other evidence indicates that language probably emerged somewhere in sub-Saharan Africa during the Middle Stone Age, roughly contemporaneous with the speciation of Homo sapiens.[34]
Language origin hypotheses
Early speculations
I cannot doubt that language owes its origin to the imitation and modification, aided by signs and gestures, of various natural sounds, the voices of other animals, and man's own instinctive cries.
— Charles Darwin, 1871. The Descent of Man, and Selection in Relation to Sex[35]
In 1861, historical linguist Max Müller published a list of speculative theories concerning the origins of spoken language:[36]
- Bow-wow. The bow-wow or cuckoo theory, which Müller attributed to the German philosopher Johann Gottfried Herder, saw early words as imitations of the cries of beasts and birds.
- Pooh-pooh. The pooh-pooh theory saw the first words as emotional interjections and exclamations triggered by pain, pleasure, surprise, etc.
- Ding-dong. Müller suggested what he called the ding-dong theory, which states that all things have a vibrating natural resonance, echoed somehow by man in his earliest words.
- Yo-he-ho. The yo-he-ho theory claims language emerged from collective rhythmic labor, the attempt to synchronize muscular effort resulting in sounds such as heave alternating with sounds such as ho.
- Ta-ta. This did not feature in Max Müller's list, having been proposed in 1930 by Sir Richard Paget.[37] According to the ta-ta theory, humans made the earliest words by tongue movements that mimicked manual gestures, rendering them audible.
Most scholars today consider all such theories not so much wrong—they occasionally offer peripheral insights—as naïve and irrelevant.[38][39] The problem with these theories is that they are so narrowly mechanistic. They assume that once our ancestors had stumbled upon the appropriate ingenious mechanism for linking sounds with meanings, language automatically evolved and changed.
Problems of reliability and deception
From the perspective of signalling theory, the main obstacle to the evolution of language-like communication in nature is not a mechanistic one. Rather, it is the fact that symbols—arbitrary associations of sounds or other perceptible forms with corresponding meanings—are unreliable and may well be false.[40] As the saying goes, "words are cheap".[41] The problem of reliability was not recognized at all by Darwin, Müller or the other early evolutionary theorists.
Animal vocal signals are, for the most part, intrinsically reliable. When a cat purrs, the signal constitutes direct evidence of the animal's contented state. We trust the signal, not because the cat is inclined to be honest, but because it just cannot fake that sound. Primate vocal calls may be slightly more manipulable, but they remain reliable for the same reason—because they are hard to fake.[42] Primate social intelligence is "Machiavellian"—self-serving and unconstrained by moral scruples. Monkeys and apes often attempt to deceive each other, while at the same time remaining constantly on guard against falling victim to deception themselves.[43][44] Paradoxically, it is theorized that primates' resistance to deception is what blocks the evolution of their signalling systems along language-like lines. Language is ruled out because the best way to guard against being deceived is to ignore all signals except those that are instantly verifiable. Words automatically fail this test.[21]
Words are easy to fake. Should they turn out to be lies, listeners will adapt by ignoring them in favor of hard-to-fake indices or cues. For language to work, then, listeners must be confident that those with whom they are on speaking terms are generally likely to be honest.[45] A peculiar feature of language is "displaced reference", which means reference to topics outside the currently perceptible situation. This property prevents utterances from being corroborated in the immediate "here" and "now". For this reason, language presupposes relatively high levels of mutual trust in order to become established over time as an evolutionarily stable strategy. This stability is born of a longstanding mutual trust and is what grants language its authority. A theory of the origins of language must therefore explain why humans could begin trusting cheap signals in ways that other animals apparently cannot (see signalling theory).
The 'mother tongues' hypothesis
The "mother tongues" hypothesis was proposed in 2004 as a possible solution to this problem.[46] W. Tecumseh Fitch suggested that the Darwinian principle of 'kin selection'[47]—the convergence of genetic interests between relatives—might be part of the answer. Fitch suggests that languages were originally 'mother tongues'. If language evolved initially for communication between mothers and their own biological offspring, extending later to include adult relatives as well, the interests of speakers and listeners would have tended to coincide. Fitch argues that shared genetic interests would have led to sufficient trust and cooperation for intrinsically unreliable signals—words—to become accepted as trustworthy and so begin evolving for the first time.[48]
Critics of this theory point out that kin selection is not unique to humans.[49] So even if one accepts Fitch's initial premises, the extension of the posited 'mother tongue' networks from close relatives to more distant relatives remains unexplained.[49] Fitch argues, however, that the extended period of physical immaturity of human infants and the postnatal growth of the human brain give the human-infant relationship a different and more extended period of intergenerational dependency than that found in any other species.[46]
The 'obligatory reciprocal altruism' hypothesis
Ib Ulbæk[6] invokes another standard Darwinian principle—'reciprocal altruism'[50]—to explain the unusually high levels of intentional honesty necessary for language to evolve. 'Reciprocal altruism' can be expressed as the principle that if you scratch my back, I'll scratch yours. In linguistic terms, it would mean that if you speak truthfully to me, I'll speak truthfully to you. Ordinary Darwinian reciprocal altruism, Ulbæk points out, is a relationship established between frequently interacting individuals. For language to prevail across an entire community, however, the necessary reciprocity would have needed to be enforced universally instead of being left to individual choice. Ulbæk concludes that for language to evolve, society as a whole must have been subject to moral regulation.
Critics point out that this theory fails to explain when, how, why or by whom 'obligatory reciprocal altruism' could possibly have been enforced.[22] Various proposals have been offered to remedy this defect.[22] A further criticism is that language does not work on the basis of reciprocal altruism anyway. Humans in conversational groups do not withhold information to all except listeners likely to offer valuable information in return. On the contrary, they seem to want to advertise to the world their access to socially relevant information, broadcasting that information without expectation of reciprocity to anyone who will listen.[51]
The gossip and grooming hypothesis
Gossip, according to Robin Dunbar in his book Grooming, Gossip and the Evolution of Language, does for group-living humans what manual grooming does for other primates—it allows individuals to service their relationships and so maintain their alliances on the basis of the principle: if you scratch my back, I'll scratch yours. Dunbar argues that as humans began living in increasingly larger social groups, the task of manually grooming all one's friends and acquaintances became so time-consuming as to be unaffordable.[52] In response to this problem, humans developed 'a cheap and ultra-efficient form of grooming'—vocal grooming. To keep allies happy, one now needs only to 'groom' them with low-cost vocal sounds, servicing multiple allies simultaneously while keeping both hands free for other tasks. Vocal grooming then evolved gradually into vocal language—initially in the form of 'gossip'.[52] Dunbar's hypothesis seems to be supported by the fact that the structure of language shows adaptations to the function of narration in general.[53]
Critics of this theory point out that the very efficiency of 'vocal grooming'—the fact that words are so cheap—would have undermined its capacity to signal commitment of the kind conveyed by time-consuming and costly manual grooming.[54] A further criticism is that the theory does nothing to explain the crucial transition from vocal grooming—the production of pleasing but meaningless sounds—to the cognitive complexities of syntactical speech.
Ritual/speech coevolution
The ritual/speech coevolution theory was originally proposed by social anthropologist Roy Rappaport[18] before being elaborated by anthropologists such as Chris Knight,[21] Jerome Lewis,[55] Nick Enfield,[56] Camilla Power[45] and Ian Watts.[30] Cognitive scientist and robotics engineer Luc Steels[57] is another prominent supporter of this general approach, as is biological anthropologist/neuroscientist Terrence Deacon.[58]
These scholars argue that there can be no such thing as a 'theory of the origins of language'. This is because language is not a separate adaptation but an internal aspect of something much wider—namely, human symbolic culture as a whole.[20] Attempts to explain language independently of this wider context have spectacularly failed, say these scientists, because they are addressing a problem with no solution. Can we imagine a historian attempting to explain the emergence of credit cards independently of the wider system of which they are a part? Using a credit card makes sense only if you have a bank account institutionally recognized within a certain kind of advanced capitalist society—one where electronic communications technology and digital computers have already been invented and fraud can be detected and prevented. In much the same way, language would not work outside a specific array of social mechanisms and institutions. For example, it would not work for a nonhuman ape communicating with others in the wild. Not even the cleverest nonhuman ape could make language work under such conditions.
Lie and alternative, inherent in language ... pose problems to any society whose structure is founded on language, which is to say all human societies. I have therefore argued that if there are to be words at all it is necessary to establish The Word, and that The Word is established by the invariance of liturgy.
— Roy Rappaport[59]
Advocates of this school of thought point out that words are cheap. As digital hallucinations, they are intrinsically unreliable.[60] Should an especially clever nonhuman ape, or even a group of articulate nonhuman apes, try to use words in the wild, they would carry no conviction. The primate vocalizations that do carry conviction—those they actually use—are unlike words, in that they are emotionally expressive, intrinsically meaningful and reliable because they are relatively costly and hard to fake.
Language consists of digital contrasts whose cost is essentially zero. As pure social conventions, signals of this kind cannot evolve in a Darwinian social world—they are a theoretical impossibility.[40] Being intrinsically unreliable, language works only if you can build up a reputation for trustworthiness within a certain kind of society—namely, one where symbolic cultural facts (sometimes called 'institutional facts') can be established and maintained through collective social endorsement.[61] In any hunter-gatherer society, the basic mechanism for establishing trust in symbolic cultural facts is collective ritual.[62] Therefore, the task facing researchers into the origins of language is more multidisciplinary than is usually supposed. It involves addressing the evolutionary emergence of human symbolic culture as a whole, with language an important but subsidiary component.
Critics of the theory include Noam Chomsky, who terms it the 'non-existence' hypothesis—a denial of the very existence of language as an object of study for natural science.[63] Chomsky's own theory is that language emerged in an instant and in perfect form,[64] prompting his critics in turn to retort that only something that does not exist—a theoretical construct or convenient scientific fiction—could possibly emerge in such a miraculous way.[19] The controversy remains unresolved.
Tool culture resilience and grammar in early Homo
While it is possible to imitate the making of tools like those made by early Homo under circumstances of demonstration, research on primate tool cultures show that non-verbal cultures are vulnerable to environmental change. In particular, if the environment in which a skill can be used disappears for a longer period of time than an individual ape's or early human's lifespan, the skill will be lost if the culture is imitative and non-verbal. Chimpanzees, macaques and capuchin monkeys are all known to lose tool techniques under such circumstances. Researchers on primate culture vulnerability therefore argue that since early Homo species as far back as Homo habilis retained their tool cultures despite many climate change cycles at the timescales of centuries to millennia each, these species had sufficiently developed language abilities to verbally describe complete procedures, and therefore grammar and not only two-word "proto-language".[65][66]
The theory that early Homo species had sufficiently developed brains for grammar is also supported by researchers who study brain development in children, noting that grammar is developed while connections across the brain are still significantly lower than adult level. These researchers argue that these lowered system requirements for grammatical language make it plausible that the genus Homo had grammar at connection levels in the brain that were significantly lower than those of Homo sapiens and that more recent steps in the evolution of the human brain were not about language.[67][68]
Humanistic theory
The humanistic tradition considers language as a human invention. Renaissance philosopher Antoine Arnauld gave a detailed description of his idea of the origin of language in Port-Royal Grammar. According to Arnauld, people are social and rational by nature, and this urged them to create language as a means to communicate their ideas to others. Language construction would have occurred through a slow and gradual process.[69] In later theory, especially in functional linguistics, the primacy of communication is emphasised over psychological needs.[70]
The exact way language evolved is however not considered as vital to the study of languages. Structural linguist Ferdinand de Saussure abandoned evolutionary linguistics after having come to the firm conclusion that it would not be able to provide any further revolutionary insight after the completion of the major works in historical linguistics by the end of the 19th century. Saussure was particularly sceptical of the attempts of August Schleicher and other Darwinian linguists to access prehistorical languages through series of reconstructions of proto-languages.[71]
Evolutionary research had many other critics, too. The Paris linguistic society famously banned the topic of language evolution in 1866 because it was considered as lacking scientific proof.[72] Around the same time, Max Müller ridiculed popular accounts to explain language origin. In his classifications, the 'bow-wow theory' is the type of explanation that considers languages as having evolved as an imitation of natural sounds. The 'pooh-pooh theory' holds that speech originated from spontaneous human cries and exclamations; the 'yo-he-ho theory' suggests that language developed from grunts and gasps evoked by physical exertion; while the 'sing-song theory' claims that speech arose from primitive ritual chants.[73]
Saussure's solution to the problem of language evolution involves dividing theoretical linguistics in two. Evolutionary and historical linguistics are renamed as diachronic linguistics. It is the study of language change, but it has only limited explanatory power due to the inadequacy of all of the reliable research material that could ever be made available. Synchronic linguistics, in contrast, aims to widen scientists' understanding of language through a study of a given contemporary or historical language stage as a system in its own right.[74]
Although Saussure paid much focus to diachronic linguistics, later structuralists who equated structuralism with the synchronic analysis were sometimes criticised of ahistoricism. According to structural anthropologist Claude Lévi-Strauss, language and meaning – in opposition to "knowledge, which develops slowly and progressively" – must have appeared in an instant.[75]
Structuralism, as first introduced to sociology by Émile Durkheim, is nonetheless a type of humanistic evolutionary theory which explains diversification as necessitated by growing complexity.[76] There was a shift of focus to functional explanation after Saussure's death. Functional structuralists including the Prague Circle linguists and André Martinet explained the growth and maintenance of structures as being necessitated by their functions.[70] For example, novel technologies make it necessary for people to invent new words, but these may lose their function and fall into oblivion as the technologies are eventually replaced by more modern ones.
Chomsky's single step theory
According to Chomsky's single mutation theory, the emergence of language resembled the formation of a crystal; with digital infinity as the seed crystal in a super-saturated primate brain, on the verge of blossoming into the human mind, by physical law, once evolution added a single small but crucial keystone.[77][64] Thus, in this theory, language appeared rather suddenly within the history of human evolution. Chomsky, writing with computational linguist and computer scientist Robert C. Berwick, suggests that this scenario is completely compatible with modern biology. They note "none of the recent accounts of human language evolution seem to have completely grasped the shift from conventional Darwinism to its fully stochastic modern version—specifically, that there are stochastic effects not only due to sampling like directionless drift, but also due to directed stochastic variation in fitness, migration, and heritability—indeed, all the "forces" that affect individual or gene frequencies.... All this can affect evolutionary outcomes—outcomes that as far as we can make out are not brought out in recent books on the evolution of language, yet would arise immediately in the case of any new genetic or individual innovation, precisely the kind of scenario likely to be in play when talking about language's emergence."
Citing evolutionary geneticist Svante Pääbo they concur that a substantial difference must have occurred to differentiate Homo sapiens from Neanderthals to "prompt the relentless spread of our species who had never crossed open water up and out of Africa and then on across the entire planet in just a few tens of thousands of years. ... What we do not see is any kind of "gradualism" in new tool technologies or innovations like fire, shelters, or figurative art." Berwick and Chomsky therefore suggest language emerged approximately between 200,000 years ago and 60,000 years ago (between the arrival of the first anatomically modern humans in southern Africa, and the last exodus from Africa, respectively). "That leaves us with about 130,000 years, or approximately 5,000–6,000 generations of time for evolutionary change. This is not 'overnight in one generation' as some have (incorrectly) inferred—but neither is it on the scale of geological eons. It's time enough—within the ballpark for what Nilsson and Pelger (1994) estimated as the time required for the full evolution of a vertebrate eye from a single cell, even without the invocation of any 'evo-devo' effects."[78]
The Romulus and Remus hypothesis
The Romulus and Remus hypothesis, proposed by neuroscientist Andrey Vyshedskiy, seeks to address the question as to why the modern speech apparatus originated over 500,000 years before the earliest signs of modern human imagination. This hypothesis proposes that there were two phases that led to modern recursive language. (In linguistics, recursion is the ability to repeat a rule indefinitely. For example, a simple sentence in English may have the form: Noun Verb, as in "I walked." Recursion means you can repeat that rule: "I walked. I ran. I played" or "I walked but I did not run because I had played too strenuously.")[79]
The first phase includes the slow development of non-recursive language with a large vocabulary along with the modern speech apparatus, which includes changes to the hyoid bone, increased voluntary control of the muscles of the diaphragm, the evolution of the FOXP2 gene, as well as other changes by 600,000 years ago.[80] Then, the second phase was a rapid Chomskian Single Step, consisting of three distinct events that happened in quick succession around 70,000 years ago and allowed for the shift from non-recursive to recursive language in early hominins.
- A genetic mutation that slowed down the Prefrontal Synthesis (PFS) critical period of at least two children that lived together;
- This allowed these children to create recursive elements of language such as spatial prepositions;
- Then this merged with their parent's non-recursive language to create recursive language.[81]
It is not enough for children to have a modern Prefrontal Cortex (PFC) to allow for the development of PFS, the children must also be mentally stimulated and have recursive elements already in their language to acquire PFS. Since, their parents would not have invented these elements yet, the children would have had to do it themselves, which is a common occurrence among young children that live together, in a process called cryptophasia.[82] This means that delayed PFC development would have allowed for more time to acquire PFS, and develop recursive elements.
Of course, delayed PFC development comes with downsides, such as a longer period of reliance on one's parents to survive, and lower survival rates. For modern language to have occurred, PFC delay had to have an immense survival benefit in later life, such as PFS ability. This suggests that the mutation that caused PFC delay and the development of recursive language and PFS occurred simultaneously, which lines up with evidence of a genetic bottleneck around 70,000 years ago.[83] This could have been the result of a few individuals who developed PFS and recursive language which gave them significant competitive advantage over all other humans at the time.[81]
Gestural theory
The gestural theory states that human language developed from gestures that were used for simple communication.
Two types of evidence support this theory.
- Gestural language and vocal language depend on similar neural systems. The regions on the cortex that are responsible for mouth and hand movements border each other.
- Nonhuman primates can use gestures or symbols for at least primitive communication, and some of their gestures resemble those of humans, such as the "begging posture", with the hands stretched out, which humans share with chimpanzees.[84]
Research has found strong support for the idea that verbal language and sign language depend on similar neural structures. Patients who used sign language, and who suffered from a left-hemisphere lesion, showed the same disorders with their sign language as vocal patients did with their oral language.[85] Other researchers found that the same left-hemisphere brain regions were active during sign language as during the use of vocal or written language.[86]
Primate gesture is at least partially genetic: different nonhuman apes will perform gestures characteristic of their species, even if they have never seen another ape perform that gesture. For example, gorillas beat their breasts. This shows that gestures are an intrinsic and important part of primate communication, which supports the idea that language evolved from gesture.[87]
Further evidence suggests that gesture and language are linked. In humans, manually gesturing has an effect on concurrent vocalizations, thus creating certain natural vocal associations of manual efforts. Chimpanzees move their mouths when performing fine motor tasks. These mechanisms may have played an evolutionary role in enabling the development of intentional vocal communication as a supplement to gestural communication. Voice modulation could have been prompted by preexisting manual actions.[87]
There is also the fact that, from infancy, gestures both supplement and predict speech.[88][89] This addresses the idea that gestures quickly change in humans from a sole means of communication (from a very young age) to a supplemental and predictive behavior that we use despite being able to communicate verbally. This too serves as a parallel to the idea that gestures developed first and language subsequently built upon it.
Two possible scenarios have been proposed for the development of language,[90] one of which supports the gestural theory:
- Language developed from the calls of our ancestors.
- Language was derived from gesture.
The first perspective that language evolved from the calls of our ancestors seems logical because both humans and animals make sounds or cries. One evolutionary reason to refute this is that, anatomically, the center that controls calls in monkeys and other animals is located in a completely different part of the brain than in humans. In monkeys, this center is located in the depths of the brain related to emotions. In the human system, it is located in an area unrelated to emotion. Humans can communicate simply to communicate—without emotions. So, anatomically, this scenario does not work.[90] Therefore, we resort to the idea that language was derived from gesture (we communicated by gesture first and sound was attached later).
The important question for gestural theories is why there was a shift to vocalization. Various explanations have been proposed:
- Our ancestors started to use more and more tools, meaning that their hands were occupied and could no longer be used for gesturing.[91]
- Manual gesturing requires that speakers and listeners be visible to one another. In many situations, they might need to communicate, even without visual contact—for example after nightfall or when foliage obstructs visibility.
- A composite hypothesis holds that early language took the form of part gestural and part vocal mimesis (imitative 'song-and-dance'), combining modalities because all signals (like those of nonhuman apes and monkeys) still needed to be costly in order to be intrinsically convincing. In that event, each multi-media display would have needed not just to disambiguate an intended meaning but also to inspire confidence in the signal's reliability. The suggestion is that only once community-wide contractual understandings had come into force[92] could trust in communicative intentions be automatically assumed, at last allowing Homo sapiens to shift to a more efficient default format. Since vocal distinctive features (sound contrasts) are ideal for this purpose, it was only at this point—when intrinsically persuasive body-language was no longer required to convey each message—that the decisive shift from manual gesture to our current primary reliance on spoken language occurred.[19][21][93]
A comparable hypothesis states that in 'articulate' language, gesture and vocalisation are intrinsically linked, as language evolved from equally intrinsically linked dance and song.[16] Humans still use manual and facial gestures when they speak, especially when people meet who have no language in common.[94] There are also, of course, a great number of sign languages still in existence, commonly associated with deaf communities. These sign languages are equal in complexity, sophistication, and expressive power, to any oral language. The cognitive functions are similar and the parts of the brain used are similar. The main difference is that the "phonemes" are produced on the outside of the body, articulated with hands, body, and facial expression, rather than inside the body articulated with tongue, teeth, lips, and breathing. (Compare the motor theory of speech perception.)
Critics of gestural theory note that it is difficult to name serious reasons why the initial pitch-based vocal communication (which is present in primates) would be abandoned in favor of the much less effective non-vocal, gestural communication. However, Michael Corballis has pointed out that it is supposed that primate vocal communication (such as alarm calls) cannot be controlled consciously, unlike hand movement, and thus is not credible as precursor to human language; primate vocalization is rather homologous to and continued in involuntary reflexes (connected with basic human emotions) such as screams or laughter (the fact that these can be faked does not disprove the fact that genuine involuntary responses to fear or surprise exist). Also, gesture is not generally less effective, and depending on the situation can even be advantageous, for example in a loud environment or where it is important to be silent, such as on a hunt. Other challenges to the "gesture-first" theory have been presented by researchers in psycholinguistics, including David McNeill.
Tool-use associated sound in the evolution of language
Proponents of the motor theory of language evolution have primarily focused on the visual domain and communication through observation of movements. The Tool-use sound hypothesis suggests that the production and perception of sound also contributed substantially, particularly incidental sound of locomotion (ISOL) and tool-use sound (TUS).[95] Human bipedalism resulted in rhythmic and more predictable ISOL. That may have stimulated the evolution of musical abilities, auditory working memory, and abilities to produce complex vocalizations, and to mimic natural sounds.[96] Since the human brain proficiently extracts information about objects and events from the sounds they produce, TUS, and mimicry of TUS, might have achieved an iconic function. The prevalence of sound symbolism in many extant languages supports this idea. Self-produced TUS activates multimodal brain processing (motor neurons, hearing, proprioception, touch, vision), and TUS stimulates primate audiovisual mirror neurons, which is likely to stimulate the development of association chains. Tool use and auditory gestures involve motor-processing of the forelimbs, which is associated with the evolution of vertebrate vocal communication. The production, perception, and mimicry of TUS may have resulted in a limited number of vocalizations or protowords that were associated with tool use.[95] A new way to communicate about tools, especially when out of sight, would have had selective advantage. A gradual change in acoustic properties and/or meaning could have resulted in arbitrariness and an expanded repertoire of words. Humans have been increasingly exposed to TUS over millions of years, coinciding with the period during which spoken language evolved.
Mirror neurons and language origins
In humans, functional MRI studies have reported finding areas homologous to the monkey mirror neuron system in the inferior frontal cortex, close to Broca's area, one of the language regions of the brain. This has led to suggestions that human language evolved from a gesture performance/understanding system implemented in mirror neurons. Mirror neurons have been said to have the potential to provide a mechanism for action-understanding, imitation-learning, and the simulation of other people's behavior.[97] This hypothesis is supported by some cytoarchitectonic homologies between monkey premotor area F5 and human Broca's area.[98]
Rates of vocabulary expansion link to the ability of children to vocally mirror non-words and so to acquire the new word pronunciations. Such speech repetition occurs automatically, quickly[99] and separately in the brain to speech perception.[100][101] Moreover, such vocal imitation can occur without comprehension such as in speech shadowing[102] and echolalia.[98][103] Further evidence for this link comes from a recent study in which the brain activity of two participants was measured using fMRI while they were gesturing words to each other using hand gestures with a game of charades—a modality that some have suggested might represent the evolutionary precursor of human language. Analysis of the data using Granger Causality revealed that the mirror-neuron system of the observer indeed reflects the pattern of activity of in the motor system of the sender, supporting the idea that the motor concept associated with the words is indeed transmitted from one brain to another using the mirror system.[104]
Not all linguists agree with the above arguments, however. In particular, supporters of Noam Chomsky argue against the possibility that the mirror neuron system can play any role in the hierarchical recursive structures essential to syntax.[105]
Putting the baby down theory
According to Dean Falk's 'putting the baby down' theory, vocal interactions between early hominid mothers and infants sparked a sequence of events that led, eventually, to our ancestors' earliest words.[106] The basic idea is that evolving human mothers, unlike their counterparts in other primates, could not move around and forage with their infants clinging onto their backs. Loss of fur in the human case left infants with no means of clinging on. Frequently, therefore, mothers had to put their babies down. As a result, these babies needed to be reassured that they were not being abandoned. Mothers responded by developing 'motherese'—an infant-directed communicative system embracing facial expressions, body language, touching, patting, caressing, laughter, tickling and emotionally expressive contact calls. The argument is that language somehow developed out of all this.[106]
In The Mental and Social Life of Babies, psychologist Kenneth Kaye noted that no usable adult language could have evolved without interactive communication between very young children and adults. "No symbolic system could have survived from one generation to the next if it could not have been easily acquired by young children under their normal conditions of social life."[107]
From where to what theory

The 'from where to what' model is a language evolution model that is derived primarily from the organization of language processing in the brain and two of its structures: the auditory dorsal stream and the auditory ventral stream.[108][109] It hypothesizes 7 stages of language evolution (see illustration). Speech originated for the purpose of exchanging contact calls between mothers and their offspring to find one another in the event they became separated (illustration part 1). The contact calls could be modified with intonations in order to express either a higher or lower level of distress (illustration part 2). The use of two types of contact calls enabled the first question-answer conversation. In this scenario, the child would emit a low-level distress call to express a desire to interact with an object, and the mother would respond with either another low-level distress call (to express approval of the interaction) or a high-level distress call (to express disapproval) (illustration part 3). Over time, the improved use of intonations and vocal control led to the invention of unique calls (phonemes) associated with distinct objects (illustration part 4). At first, children learned the calls (phonemes) from their parents by imitating their lip-movements (illustration part 5). Eventually, infants were able to encode into long-term memory all the calls (phonemes). Consequentially, mimicry via lip-reading was limited to infancy and older children learned new calls through mimicry without lip-reading (illustration part 6). Once individuals became capable of producing a sequence of calls, this allowed multi-syllabic words, which increased the size of their vocabulary (illustration part 7). The use of words, composed of sequences of syllables, provided the infra structure for communicating with sequences of words (i.e., sentences).
The theory's name is derived from the two auditory streams, which are both found in the brains of humans and other primates. The auditory ventral stream is responsible for sound recognition, and so it is referred to as the auditory what stream.[110][111][112] In primates, the auditory dorsal stream is responsible for sound localization. It's a so-called auditory where stream. Only in humans (in the left hemisphere), is it also responsible for other processes associated with language use and acquisition, such as speech repetition and production, integration of phonemes with their lip movements, perception and production of intonations, phonological long-term memory (long-term memory storage of the sounds of words), and phonological working memory (the temporary storage of the sounds of words).[113][114][115][116][117][118][119][120] Some evidence also indicates a role in recognizing others by their voices.[121][122] The emergence of each of these functions in the auditory dorsal stream represents an intermediate stage in the evolution of language.
A contact call origin for human language is consistent with animal studies, as like human language, contact call discrimination in monkeys is lateralized to the left hemisphere.[123][124] Mice with knock-out to language related genes (such as FOXP2 and SRPX2) also resulted with the pups no longer emitting contact calls when separated from their mothers.[125][126] Supporting this model is also its ability to explain unique human phenomena, such as the use of intonations when converting words into commands and questions, the tendency of infants to mimic vocalizations during the first year of life (and its disappearance later on) and the protruding and visible human lips, which are not found in other apes. This theory could be considered an elaboration of the 'putting the baby down' theory of language evolution.
Grammaticalisation theory
'Grammaticalisation' is a continuous historical process in which free-standing words develop into grammatical appendages, while these in turn become ever more specialized and grammatical. An initially 'incorrect' usage, in becoming accepted, leads to unforeseen consequences, triggering knock-on effects and extended sequences of change. Paradoxically, grammar evolves because, in the final analysis, humans care less about grammatical niceties than about making themselves understood.[127] If this is how grammar evolves today, according to this school of thought, we can legitimately infer similar principles at work among our distant ancestors, when grammar itself was first being established.[128][129][130]
In order to reconstruct the evolutionary transition from early language to languages with complex grammars, we need to know which hypothetical sequences are plausible and which are not. In order to convey abstract ideas, the first recourse of speakers is to fall back on immediately recognizable concrete imagery, very often deploying metaphors rooted in shared bodily experience.[131] A familiar example is the use of concrete terms such as 'belly' or 'back' to convey abstract meanings such as 'inside' or 'behind'. Equally metaphorical is the strategy of representing temporal patterns on the model of spatial ones. For example, English speakers might say 'It is going to rain,' modeled on 'I am going to London.' This can be abbreviated colloquially to 'It's gonna rain.' Even when in a hurry, English speakers do not say 'I'm gonna London'—the contraction is restricted to the job of specifying tense. From such examples we can see why grammaticalization is consistently unidirectional—from concrete to abstract meaning, not the other way around.[128]
Grammaticalization theorists picture early language as simple, perhaps consisting only of nouns.[130]p. 111 Even under that extreme theoretical assumption, however, it is difficult to imagine what would realistically have prevented people from using, say, 'spear' as if it were a verb ('Spear that pig!'). People might have used their nouns as verbs or their verbs as nouns as occasion demanded. In short, while a noun-only language might seem theoretically possible, grammaticalization theory indicates that it cannot have remained fixed in that state for any length of time.[128][132]
Creativity drives grammatical change.[132] This presupposes a certain attitude on the part of listeners. Instead of punishing deviations from accepted usage, listeners must prioritize imaginative mind-reading. Imaginative creativity—emitting a leopard alarm when no leopard was present, for example—is not the kind of behavior which, say, vervet monkeys would appreciate or reward.[133] Creativity and reliability are incompatible demands; for 'Machiavellian' primates as for animals generally, the overriding pressure is to demonstrate reliability.[134] If humans escape these constraints, it is because in our case, listeners are primarily interested in mental states.
To focus on mental states is to accept fictions—inhabitants of the imagination—as potentially informative and interesting. Take the use of metaphor. A metaphor is, literally, a false statement.[135] Think of Romeo's declaration, 'Juliet is the sun!' Juliet is a woman, not a ball of plasma in the sky, but human listeners are not (or not usually) pedants insistent on point-by-point factual accuracy. They want to know what the speaker has in mind. Grammaticalization is essentially based on metaphor. To outlaw its use would be to stop grammar from evolving and, by the same token, to exclude all possibility of expressing abstract thought.[131][136]
A criticism of all this is that while grammaticalization theory might explain language change today, it does not satisfactorily address the really difficult challenge—explaining the initial transition from primate-style communication to language as we know it. Rather, the theory assumes that language already exists. As Bernd Heine and Tania Kuteva acknowledge: "Grammaticalization requires a linguistic system that is used regularly and frequently within a community of speakers and is passed on from one group of speakers to another".[130] Outside modern humans, such conditions do not prevail.
Evolution-Progression Model
Human language is used for self-expression; however, expression displays different stages. The consciousness of self and feelings represents the stage immediately prior to the external, phonetic expression of feelings in the form of sound, i.e., language. Intelligent animals such as dolphins, Eurasian magpies, and chimpanzees live in communities, wherein they assign themselves roles for group survival and show emotions such as sympathy.[137] When such animals view their reflection (mirror test), they recognize themselves and exhibit self-consciousness.[138] Notably, humans evolved in a quite different environment than that of these animals. Human survival became easier with the development of tools, shelter, and fire, thus facilitating further advancement of interaction, self-expression, and tool-making.[139] The increasing brain size allowed advanced provisioning and tools and the technological advances during the Palaeolithic era that built upon the previous evolutionary innovations of bipedalism and hand versatility allowed the development of human language.
Self-domesticated ape theory
According to a study investigating the song differences between white-rumped munias and its domesticated counterpart (Bengalese finch), the wild munias use a highly stereotyped song sequence, whereas the domesticated ones sing a highly unconstrained song. In wild finches, song syntax is subject to female preference—sexual selection—and remains relatively fixed. However, in the Bengalese finch, natural selection is replaced by breeding, in this case for colorful plumage, and thus, decoupled from selective pressures, stereotyped song syntax is allowed to drift. It is replaced, supposedly within 1000 generations, by a variable and learned sequence. Wild finches, moreover, are thought incapable of learning song sequences from other finches.[140] In the field of bird vocalization, brains capable of producing only an innate song have very simple neural pathways: the primary forebrain motor center, called the robust nucleus of arcopallium, connects to midbrain vocal outputs, which in turn project to brainstem motor nuclei. By contrast, in brains capable of learning songs, the arcopallium receives input from numerous additional forebrain regions, including those involved in learning and social experience. Control over song generation has become less constrained, more distributed, and more flexible.[140]
One way to think about human evolution is that we are self-domesticated apes. Just as domestication relaxed selection for stereotypic songs in the finches—mate choice was supplanted by choices made by the aesthetic sensibilities of bird breeders and their customers—so might our cultural domestication have relaxed selection on many of our primate behavioral traits, allowing old pathways to degenerate and reconfigure. Given the highly indeterminate way that mammalian brains develop—they basically construct themselves "bottom up", with one set of neuronal interactions setting the stage for the next round of interactions—degraded pathways would tend to seek out and find new opportunities for synaptic hookups. Such inherited de-differentiations of brain pathways might have contributed to the functional complexity that characterizes human language. And, as exemplified by the finches, such de-differentiations can occur in very rapid time-frames.[141]
Speech and language for communication
A distinction can be drawn between speech and language. Language is not necessarily spoken: it might alternatively be written or signed. Speech is among a number of different methods of encoding and transmitting linguistic information, albeit arguably the most natural one.[142]
Some scholars view language as an initially cognitive development, its 'externalisation' to serve communicative purposes occurring later in human evolution. According to one such school of thought, the key feature distinguishing human language is recursion,[143] (in this context, the iterative embedding of phrases within phrases). Other scholars—notably Daniel Everett—deny that recursion is universal, citing certain languages (e.g. Pirahã) which allegedly lack this feature.[144]
The ability to ask questions is considered by some to distinguish language from non-human systems of communication.[145] Some captive primates (notably bonobos and chimpanzees), having learned to use rudimentary signing to communicate with their human trainers, proved able to respond correctly to complex questions and requests. Yet they failed to ask even the simplest questions themselves. Conversely, human children are able to ask their first questions (using only question intonation) at the babbling period of their development, long before they start using syntactic structures. Although babies from different cultures acquire native languages from their social environment, all languages of the world without exception—tonal, non-tonal, intonational and accented—use similar rising "question intonation" for yes–no questions.[146][147] This fact is a strong evidence of the universality of question intonation. In general, according to some authors, sentence intonation/pitch is pivotal in spoken grammar and is the basic information used by children to learn the grammar of whatever language.[16]
Cognitive development and language
One of the intriguing abilities that language users have is that of high-level reference (or deixis), the ability to refer to things or states of being that are not in the immediate realm of the speaker. This ability is often related to theory of mind, or an awareness of the other as a being like the self with individual wants and intentions. According to Chomsky, Hauser and Fitch (2002), there are six main aspects of this high-level reference system:
- Theory of mind
- Capacity to acquire non-linguistic conceptual representations, such as the object/kind distinction
- Referential vocal signals
- Imitation as a rational, intentional system
- Voluntary control over signal production as evidence of intentional communication
- Number representation[143]
Theory of mind
Simon Baron-Cohen (1999) argues that theory of mind must have preceded language use, based on evidence of use of the following characteristics as much as 40,000 years ago: intentional communication, repairing failed communication, teaching, intentional persuasion, intentional deception, building shared plans and goals, intentional sharing of focus or topic, and pretending. Moreover, Baron-Cohen argues that many primates show some, but not all, of these abilities. Call and Tomasello's research on chimpanzees supports this, in that individual chimps seem to understand that other chimps have awareness, knowledge, and intention, but do not seem to understand false beliefs. Many primates show some tendencies toward a theory of mind, but not a full one as humans have. Ultimately, there is some consensus within the field that a theory of mind is necessary for language use. Thus, the development of a full theory of mind in humans was a necessary precursor to full language use.
Number representation
In one particular study, rats and pigeons were required to press a button a certain number of times to get food. The animals showed very accurate distinction for numbers less than four, but as the numbers increased, the error rate increased.[143] Matsuzawa (1985) attempted to teach chimpanzees Arabic numerals. The difference between primates and humans in this regard was very large, as it took the chimps thousands of trials to learn 1–9 with each number requiring a similar amount of training time; yet, after learning the meaning of 1, 2 and 3 (and sometimes 4), children easily comprehend the value of greater integers by using a successor function (i.e. 2 is 1 greater than 1, 3 is 1 greater than 2, 4 is 1 greater than 3; once 4 is reached it seems most children have an "a-ha!" moment and understand that the value of any integer n is 1 greater than the previous integer). Put simply, other primates learn the meaning of numbers one by one, similar to their approach to other referential symbols, while children first learn an arbitrary list of symbols (1, 2, 3, 4...) and then later learn their precise meanings.[148] These results can be seen as evidence for the application of the "open-ended generative property" of language in human numeral cognition.[143]
Linguistic structures
Lexical-phonological principle
Hockett (1966) details a list of features regarded as essential to describing human language.[149] In the domain of the lexical-phonological principle, two features of this list are most important:
- Productivity: users can create and understand completely novel messages.
- New messages are freely coined by blending, analogizing from, or transforming old ones.
- Either new or old elements are freely assigned new semantic loads by circumstances and context. This says that in every language, new idioms constantly come into existence.
- Duality (of Patterning): a large number of meaningful elements are made up of a conveniently small number of independently meaningless yet message-differentiating elements.
The sound system of a language is composed of a finite set of simple phonological items. Under the specific phonotactic rules of a given language, these items can be recombined and concatenated, giving rise to morphology and the open-ended lexicon. A key feature of language is that a simple, finite set of phonological items gives rise to an infinite lexical system wherein rules determine the form of each item, and meaning is inextricably linked with form. Phonological syntax, then, is a simple combination of pre-existing phonological units. Related to this is another essential feature of human language: lexical syntax, wherein pre-existing units are combined, giving rise to semantically novel or distinct lexical items.
Certain elements of the lexical-phonological principle are known to exist outside of humans. While all (or nearly all) have been documented in some form in the natural world, very few coexist within the same species. Bird-song, singing nonhuman apes, and the songs of whales all display phonological syntax, combining units of sound into larger structures apparently devoid of enhanced or novel meaning. Certain other primate species do have simple phonological systems with units referring to entities in the world. However, in contrast to human systems, the units in these primates' systems normally occur in isolation, betraying a lack of lexical syntax. There is new evidence to suggest that Campbell's monkeys also display lexical syntax, combining two calls (a predator alarm call with a "boom", the combination of which denotes a lessened threat of danger), however it is still unclear whether this is a lexical or a morphological phenomenon.
Pidgins and creoles
Pidgins are significantly simplified languages with only rudimentary grammar and a restricted vocabulary. In their early stage, pidgins mainly consist of nouns, verbs, and adjectives with few or no articles, prepositions, conjunctions or auxiliary verbs. Often the grammar has no fixed word order and the words have no inflection.[150]
If contact is maintained between the groups speaking the pidgin for long periods of time, the pidgins may become more complex over many generations. If the children of one generation adopt the pidgin as their native language it develops into a creole language, which becomes fixed and acquires a more complex grammar, with fixed phonology, syntax, morphology, and syntactic embedding. The syntax and morphology of such languages may often have local innovations not obviously derived from any of the parent languages.
Studies of creole languages around the world have suggested that they display remarkable similarities in grammar and are developed uniformly from pidgins in a single generation. These similarities are apparent even when creoles do not share any common language origins. In addition, creoles share similarities despite being developed in isolation from each other. Syntactic similarities include subject–verb–object word order. Even when creoles are derived from languages with a different word order they often develop the SVO word order. Creoles tend to have similar usage patterns for definite and indefinite articles, and similar movement rules for phrase structures even when the parent languages do not.[150]
Evolutionary timeline
Primate communication
Field primatologists can give us useful insights into great ape communication in the wild.[31] An important finding is that nonhuman primates, including the other great apes, produce calls that are graded, as opposed to categorically differentiated, with listeners striving to evaluate subtle gradations in signalers' emotional and bodily states. Nonhuman apes seemingly find it extremely difficult to produce vocalizations in the absence of the corresponding emotional states.[42] In captivity, nonhuman apes have been taught rudimentary forms of sign language or have been persuaded to use lexigrams—symbols that do not graphically resemble the corresponding words—on computer keyboards. Some nonhuman apes, such as Kanzi, have been able to learn and use hundreds of lexigrams.[151][152]
The Broca's and Wernicke's areas in the primate brain are responsible for controlling the muscles of the face, tongue, mouth, and larynx, as well as recognizing sounds. Primates are known to make "vocal calls", and these calls are generated by circuits in the brainstem and limbic system.[153]
In the wild, the communication of vervet monkeys has been the most extensively studied.[150] They are known to make up to ten different vocalizations. Many of these are used to warn other members of the group about approaching predators. They include a "leopard call", a "snake call", and an "eagle call".[154] Each call triggers a different defensive strategy in the monkeys who hear the call and scientists were able to elicit predictable responses from the monkeys using loudspeakers and prerecorded sounds. Other vocalizations may be used for identification. If an infant monkey calls, its mother turns toward it, but other vervet mothers turn instead toward that infant's mother to see what she will do.[155][156]
Similarly, researchers have demonstrated that chimpanzees (in captivity) use different "words" in reference to different foods. They recorded vocalizations that chimps made in reference, for example, to grapes, and then other chimps pointed at pictures of grapes when they heard the recorded sound.[157][158]
Ardipithecus ramidus
A study published in HOMO: Journal of Comparative Human Biology in 2017 claims that Ardipithecus ramidus, a hominin dated at approximately 4.5Ma, shows the first evidence of an anatomical shift in the hominin lineage suggestive of increased vocal capability.[159] This study compared the skull of A. ramidus with twenty nine chimpanzee skulls of different ages and found that in numerous features A. ramidus clustered with the infant and juvenile measures as opposed to the adult measures. Significantly, such affinity with the shape dimensions of infant and juvenile chimpanzee skull architecture was argued may have resulted in greater vocal capability. This assertion was based on the notion that the chimpanzee vocal tract ratios that prevent speech are a result of growth factors associated with puberty—growth factors absent in A. ramidus ontogeny. A. ramidus was also found to have a degree of cervical lordosis more conducive to vocal modulation when compared with chimpanzees as well as cranial base architecture suggestive of increased vocal capability.
What was significant in this study was the observation that the changes in skull architecture that correlate with reduced aggression are the same changes necessary for the evolution of early hominin vocal ability. In integrating data on anatomical correlates of primate mating and social systems with studies of skull and vocal tract architecture that facilitate speech production, the authors argue that paleoanthropologists to date have failed to grasp the important relationship between early hominin social evolution and language capacity.
While the skull of A. ramidus, according to the authors, lacks the anatomical impediments to speech evident in chimpanzees, it is unclear what the vocal capabilities of this early hominin were. While they suggest A. ramidus—based on similar vocal tract ratios—may have had vocal capabilities equivalent to a modern human infant or very young child, they concede this is obviously a debatable and speculative hypothesis. However, they do claim that changes in skull architecture through processes of social selection were a necessary prerequisite for language evolution. As they write:
We propose that as a result of paedomorphic morphogenesis of the cranial base and craniofacial morphology Ar. ramidus would have not been limited in terms of the mechanical components of speech production as chimpanzees and bonobos are. It is possible that Ar. ramidus had vocal capability approximating that of chimpanzees and bonobos, with its idiosyncratic skull morphology not resulting in any significant advances in speech capability. In this sense the anatomical features analysed in this essay would have been exapted in later more voluble species of hominin. However, given the selective advantages of pro-social vocal synchrony, we suggest the species would have developed significantly more complex vocal abilities than chimpanzees and bonobos.[159]
Early Homo
Anatomically, some scholars believe that features of bipedalism developed in the australopithecines around 3.5 million years ago. Around this time, these structural developments within the skull led to a more prominently L-shaped vocal tract.[160] In order to generate the sounds modern homo sapiens are capable of making, such as vowels, it is vital that Early Homo populations must have a specifically shaped voice track and a lower sitting larynx.[161] Opposing research previously suggested that Neanderthals were physically incapable of creating the full range of vocals seen in modern humans due to the differences in larynx placement. Establishing distinct larynx positions through fossil remains of Homo sapiens and Neanderthals would support this theory, however, modern research has revealed that the hyoid bone was indistinguishable from the two populations. Though research has shown a lower sitting larynx is important to producing speech, another theory states it may not be as important as once thought.[162] Cataldo, Migliano, & Vinicius (2018) stated that speech may have emerged due to an increase in trade and communication between different groups. Another view by Cataldo states that speech was evolved to enable tool-making by the Neanderthals.[163]
Archaic Homo sapiens
Steven Mithen proposed the term Hmmmmm for the pre-linguistic system of communication posited to have been used by archaic Homo, beginning with Homo ergaster and reaching the highest sophistication in the Middle Pleistocene with Homo heidelbergensis and Homo neanderthalensis. Hmmmmm is an acronym for holistic (non-compositional), manipulative (utterances are commands or suggestions, not descriptive statements), multi-modal (acoustic as well as gestural and facial), musical, and mimetic.[164]
Homo heidelbergensis
Homo heidelbergensis was a close relative (most probably a migratory descendant) of Homo ergaster. Some researchers believe this species to be the first hominin to make controlled vocalizations, possibly mimicking animal vocalizations,[164] and that as Homo heidelbergensis developed more sophisticated culture, proceeded from this point and possibly developed an early form of symbolic language.
Homo neanderthalensis
The discovery in 1989 of the (Neanderthal) Kebara 2 hyoid bone suggests that Neanderthals may have been anatomically capable of producing sounds similar to modern humans.[165][166] The hypoglossal nerve, which passes through the hypoglossal canal, controls the movements of the tongue, which may have enabled voicing for size exaggeration (see size exaggeration hypothesis below) or may reflect speech abilities.[26][167][168][169][170][171]
However, although Neanderthals may have been anatomically able to speak, Richard G. Klein in 2004 doubted that they possessed a fully modern language. He largely bases his doubts on the fossil record of archaic humans and their stone tool kit. Bart de Boer in 2017 acknowledges this ambiguity of a universally accepted Neanderthal vocal tract, however he notes the similarities in the thoracic vertebral canal, potential air sacs, and hyoid bones between modern humans and Neanderthals to suggest the presence of complex speech.[172] For 2 million years following the emergence of Homo habilis, the stone tool technology of hominins changed very little. Klein, who has worked extensively on ancient stone tools, describes the crude stone tool kit of archaic humans as impossible to break down into categories based on their function, and reports that Neanderthals seem to have had little concern for the final aesthetic form of their tools. Klein argues that the Neanderthal brain may have not reached the level of complexity required for modern speech, even if the physical apparatus for speech production was well-developed.[173][174] The issue of the Neanderthal's level of cultural and technological sophistication remains a controversial one.
Based on computer simulations used to evaluate that evolution of language that resulted in showing three stages in the evolution of syntax, Neanderthals are thought to have been in stage 2, showing they had something more evolved than proto-language but not quite as complex as the language of modern humans.[175]
Homo sapiens
Anatomically modern humans begin to appear in the fossil record in Ethiopia some 200,000 years ago.[176] Although there is still much debate as to whether behavioural modernity emerged in Africa at around the same time, a growing number of archaeologists nowadays invoke the southern African Middle Stone Age use of red ochre pigments—for example at Blombos Cave—as evidence that modern anatomy and behaviour co-evolved.[177] These archaeologists argue strongly that if modern humans at this early stage were using red ochre pigments for ritual and symbolic purposes, they probably had symbolic language as well.[28]
According to the recent African origins hypothesis, from around 60,000 – 50,000 years ago[178] a group of humans left Africa and began migrating to occupy the rest of the world, carrying language and symbolic culture with them.[179]
The descended larynx
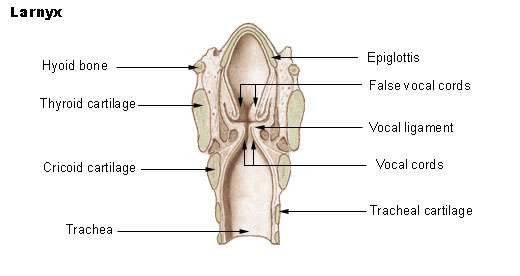
The larynx or voice box is an organ in the neck housing the vocal folds, which are responsible for phonation. In humans, the larynx is descended. Our species is not unique in this respect: goats, dogs, pigs and tamarins lower the larynx temporarily, to emit loud calls.[180] Several deer species have a permanently lowered larynx, which may be lowered still further by males during their roaring displays.[181] Lions, jaguars, cheetahs and domestic cats also do this.[182] However, laryngeal descent in nonhumans (according to Philip Lieberman) is not accompanied by descent of the hyoid; hence the tongue remains horizontal in the oral cavity, preventing it from acting as a pharyngeal articulator.[183]
| Larynx | |
|---|---|
 Anatomy of the larynx, anterolateral view | |
| Anatomical terminology |
Despite all this, scholars remain divided as to how "special" the human vocal tract really is. It has been shown that the larynx does descend to some extent during development in chimpanzees, followed by hyoidal descent.[184] As against this, Philip Lieberman points out that only humans have evolved permanent and substantial laryngeal descent in association with hyoidal descent, resulting in a curved tongue and two-tube vocal tract with 1:1 proportions. Uniquely in the human case, simple contact between the epiglottis and velum is no longer possible, disrupting the normal mammalian separation of the respiratory and digestive tracts during swallowing. Since this entails substantial costs—increasing the risk of choking while swallowing food—we are forced to ask what benefits might have outweighed those costs. The obvious benefit—so it is claimed—must have been speech. But this idea has been vigorously contested. One objection is that humans are in fact not seriously at risk of choking on food: medical statistics indicate that accidents of this kind are extremely rare.[185] Another objection is that in the view of most scholars, speech as we know it emerged relatively late in human evolution, roughly contemporaneously with the emergence of Homo sapiens.[33] A development as complex as the reconfiguration of the human vocal tract would have required much more time, implying an early date of origin. This discrepancy in timescales undermines the idea that human vocal flexibility was initially driven by selection pressures for speech, thus not excluding that it was selected for e.g. improved singing ability.
The size exaggeration hypothesis
To lower the larynx is to increase the length of the vocal tract, in turn lowering formant frequencies so that the voice sounds "deeper"—giving an impression of greater size. John Ohala argues that the function of the lowered larynx in humans, especially males, is probably to enhance threat displays rather than speech itself.[186] Ohala points out that if the lowered larynx were an adaptation for speech, we would expect adult human males to be better adapted in this respect than adult females, whose larynx is considerably less low. In fact, females invariably outperform males in verbal tests, falsifying this whole line of reasoning. W. Tecumseh Fitch likewise argues that this was the original selective advantage of laryngeal lowering in our species. Although (according to Fitch) the initial lowering of the larynx in humans had nothing to do with speech, the increased range of possible formant patterns was subsequently co-opted for speech. Size exaggeration remains the sole function of the extreme laryngeal descent observed in male deer. Consistent with the size exaggeration hypothesis, a second descent of the larynx occurs at puberty in humans, although only in males. In response to the objection that the larynx is descended in human females, Fitch suggests that mothers vocalising to protect their infants would also have benefited from this ability.[187]
Phonemic diversity
In 2011, Quentin Atkinson published a survey of phonemes from 500 different languages as well as language families and compared their phonemic diversity by region, number of speakers and distance from Africa. The survey revealed that African languages had the largest number of phonemes, and Oceania and South America had the smallest number. After allowing for the number of speakers, the phonemic diversity was compared to over 2000 possible origin locations. Atkinson's "best fit" model is that language originated in central and southern Africa between 80,000 and 160,000 years ago. This predates the hypothesized southern coastal peopling of Arabia, India, southeast Asia, and Australia. It would also mean that the origin of language occurred at the same time as the emergence of symbolic culture.[13]
History
In religion and mythology
The search for the origin of language has a long history rooted in mythology. Most mythologies do not credit humans with the invention of language but speak of a divine language predating human language. Mystical languages used to communicate with animals or spirits, such as the language of the birds, are also common, and were of particular interest during the Renaissance.
Vāc is the Hindu goddess of speech, or "speech personified". As Brahman's "sacred utterance", she has a cosmological role as the "Mother of the Vedas". The Aztecs' story maintains that only a man, Coxcox, and a woman, Xochiquetzal, survived a flood, having floated on a piece of bark. They found themselves on land and begat many children who were at first born unable to speak, but subsequently, upon the arrival of a dove, were endowed with language, although each one was given a different speech such that they could not understand one another.[188]
In the Old Testament, the Book of Genesis (11) says that God prevented the Tower of Babel from being completed through a miracle that made its construction workers start speaking different languages. After this, they migrated to other regions, grouped together according to which of the newly created languages they spoke, explaining the origins of languages and nations outside of the fertile crescent.[189]
Historical experiments
History contains a number of anecdotes about people who attempted to discover the origin of language by experiment. The first such tale was told by Herodotus (Histories 2.2). He relates that Pharaoh Psammetichus (probably Psammetichus I, 7th century BC) had two children raised by a shepherd, with the instructions that no one should speak to them, but that the shepherd should feed and care for them while listening to determine their first words. When one of the children cried "bekos" with outstretched arms the shepherd concluded that the word was Phrygian, because that was the sound of the Phrygian word for "bread". From this, Psammetichus concluded that the first language was Phrygian. King James V of Scotland is said to have tried a similar experiment; his children were supposed to have spoken Hebrew.[190]
Both the medieval monarch Frederick II and Akbar are said to have tried similar experiments; the children involved in these experiments did not speak. The current situation of deaf people also points into this direction.
History of research
Modern linguistics does not begin until the late 18th century, and the Romantic or animist theses of Johann Gottfried Herder and Johann Christoph Adelung remained influential well into the 19th century. The question of language origin seemed inaccessible to methodical approaches, and in 1866 the Linguistic Society of Paris famously banned all discussion of the origin of language, deeming it to be an unanswerable problem. An increasingly systematic approach to historical linguistics developed in the course of the 19th century, reaching its culmination in the Neogrammarian school of Karl Brugmann and others.
However, scholarly interest in the question of the origin of language has only gradually been rekindled from the 1950s on (and then controversially) with ideas such as universal grammar, mass comparison and glottochronology.
The "origin of language" as a subject in its own right emerged from studies in neurolinguistics, psycholinguistics and human evolution. The Linguistic Bibliography introduced "Origin of language" as a separate heading in 1988, as a sub-topic of psycholinguistics. Dedicated research institutes of evolutionary linguistics are a recent phenomenon, emerging only in the 1990s.
See also
- Abiogenesis
- Biolinguistics
- Bow-wow theory
- Digital infinity
- Essay on the Origin of Languages
- Evolutionary psychology of language
- FOXP2 and human evolution
- Generative anthropology
- Historical linguistics
- Man's First Word (children's book)
- Neurobiological origins of language
- Origins of society
- Origin of speech
- Proto-language
- Theory of language
References
- Hauser, M. D.; Yang, C.; Berwick, R. C.; Tattersall, I.; Ryan, M. J.; Watumull, J.; Chomsky, N.; Lewontin, R. C. (2014). "The mystery of language evolution". Frontiers in Psychology. 5: 401. doi:10.3389/fpsyg.2014.00401. PMC 4019876. PMID 24847300.
- Stam, J. H. 1976. Inquiries into the origins of language. New York: Harper and Row, p. 255.
- Tallerman, Maggie; Gibson, Kathleen Rita (2012). The Oxford handbook of language evolution. Oxford; New York: Oxford University Press. ISBN 978-0-19-954111-9. OCLC 724665645.
- Müller, F. M. 1996 [1861]. The theoretical stage, and the origin of language. Lecture 9 from Lectures on the Science of Language. Reprinted in R. Harris (ed.), The Origin of Language. Bristol: Thoemmes Press, pp. 7–41.
- Christiansen, Morten H; Kirby, Simon (2003). Morten H. Christiansen; Simon Kirby (eds.). Language evolution : the hardest problem in science?. Language evolution. Oxford; New York: Oxford University Press. pp. 77–93. ISBN 978-0-19-924484-3. OCLC 51235137.
- Ulbæk, Ib (1998). James R Hurford; Michael Studdert-Kennedy; Chris Knight (eds.). The origin of language and cognition. Approaches to the evolution of language : social and cognitive base. Cambridge, UK; New York: Cambridge University Press. pp. 30–43. ISBN 978-0-521-63964-4. OCLC 37742390.
- Chomsky, N, 1996. Powers and Prospects. Reflections on human nature and the social order. London: Pluto Press, p 30.
- Pinker, S.; Bloom, P. (1990). "Natural language and natural selection". Behavioral and Brain Sciences. 13 (4): 707–784. doi:10.1017/S0140525X00081061.
- Pinker, Steven (1994). The language instinct. New York: W. Morrow and Co. ISBN 978-0-688-12141-9. OCLC 28723210.
- Tomasello, Michael (1996). B M Velichkovskiĭ; Duane M Rumbaugh; Universität Bielefeld Zentrum für Interdisziplinäre Forschung (eds.). The cultural roots of language. Communicating meaning : the evolution and development of language. Mahwah, N.J.: L. Erlbaum. ISBN 978-0-8058-2118-5. OCLC 34078362.
- Pika, Simone; Mitani, John (2006). "Referential gestural communication in wild chimpanzees (Pan troglodytes)". Current Biology. 16 (6): R191–R192. doi:10.1016/j.cub.2006.02.037. ISSN 0960-9822. PMID 16546066.
- Dunn, M.; Greenhill, SJ.; Levinson, SC.; Gray, RD. (May 2011). "Evolved structure of language shows lineage-specific trends in word-order universals". Nature. 473 (7345): 79–82. Bibcode:2011Natur.473...79D. doi:10.1038/nature09923. hdl:11858/00-001M-0000-0013-3B19-B. PMID 21490599.
- Atkinson, Quentin (2011). "Phonemic Diversity Supports a Serial Founder Effect Model of Language Expansion from Africa" (PDF). Science Magazine. 332 (6027): 346–349. Bibcode:2011Sci...332..346A. doi:10.1126/science.1199295. PMID 21493858. Retrieved 9 July 2017.
- The Economist, "The evolution of language: Babel or babble?", 16 April 2011, pp. 85–86.
- Cross, Ian; Woodruff, Ghofur Eliot (23 April 2009). "Music as a Communicative medium". In Rudolf Botha; Chris Knight (eds.). The Prehistory of Language. OUP Oxford. pp. 77–98. doi:10.1093/acprof:oso/9780199545872.003.0005. ISBN 978-0-19-156287-7.
- Vaneechoutte, Mario (2014). "The Origin of Articulate Language Revisited: The Potential of a Semi-Aquatic Past of Human Ancestors to Explain the Origin of Human Musicality and Articulate Language" (PDF). Human Evolution. 29: 1–33.
- Knight, Chris; Power, Camilla (2012). Maggie Tallerman; Kathleen R. Gibson (eds.). Social conditions for the evolutionary emergence of language (PDF). The Oxford handbook of language evolution. Oxford; New York: Oxford University Press. pp. 346–49. ISBN 978-0-19-954111-9. OCLC 724665645.
- Rappaport, Roy (1999). Ritual and religion in the making of humanity. Cambridge, U.K. New York: Cambridge University Press. ISBN 9780521296908. OCLC 848728046.
- Knight, C. (2008). "'Honest fakes' and language origins" (PDF). Journal of Consciousness Studies. 15 (10–11): 236–48.
- Knight, Chris (2010). Ulrich J Frey; Charlotte Störmer; Kai P Willführ (eds.). The origins of symbolic culture (PDF). Homo novus : a human without illusion. Berlin; New York: Springer. pp. 193–211. ISBN 978-3-642-12141-8. OCLC 639461749.
- Knight, Chris (1998). James R Hurford; Michael Studdert-Kennedy; Chris Knight (eds.). Ritual/speech coevolution: a solution to the problem of deception (PDF). Approaches to the evolution of language : social and cognitive base. Cambridge, UK; New York: Cambridge University Press. pp. 68–91. ISBN 978-0-521-63964-4. OCLC 37742390.
- Knight, Chris (2006). Angelo Cangelosi; Andrew D M Smith; Kenny Smith (eds.). Language co-evolved with the rule of law (PDF). The evolution of language : proceedings of the 6th international conference (EVOLANG6), Rome, Italy, 12–15 April 200. New Jersey: World Scientific Publishing. pp. 168–175. ISBN 978-981-256-656-0. OCLC 70797781.
- Savage-Rumbaugh, Sue; McDonald, Kelly (1988). Richard W Byrne; Andrew Whiten (eds.). Deception and social manipulation in symbol-using apes. Machiavellian intelligence : social expertise and the evolution of intellect in monkeys, apes, and human. Oxford: Clarendon Press. pp. 224–237. ISBN 978-0-19-852175-4. OCLC 17260831.
- Kegl, J., A. Senghas and M. Coppola (1998). Creation through Contact: Sign language emergence and sign language change in Nicaragua. In M. DeGraff (ed.), Language Creation and Change: Creolization, Diachrony and Development. Cambridge, Massachusetts: MIT Press.
- Lieberman, P.; Crelin, E. S. (1971). "On the speech of Neandertal Man". Linguistic Inquiry. 2: 203–22.
- Arensburg, B.; Tillier, A. M.; Vandermeersch, B.; Duday, H.; Schepartz, L. A.; Rak, Y. (1989). "A Middle Palaeolithic human hyoid bone". Nature. 338 (6218): 758–760. Bibcode:1989Natur.338..758A. doi:10.1038/338758a0. PMID 2716823.
- Diller, Karl C.; Cann, Rebecca L. (2009). Rudolf P Botha; Chris Knight (eds.). Evidence Against a Genetic-Based Revolution in Language 50,000 Years Ago. The cradle of language. Oxford; New York: Oxford University Press. pp. 135–149. ISBN 978-0-19-954586-5. OCLC 804498749.
- Henshilwood, Christopher Stuart; Dubreuil, Benoît (2009). Rudolf P Botha; Chris Knight (eds.). Reading the Artefacts: Gleaning Language Skills From the Middle Stone Age in Southern Africa. The cradle of language. Oxford; New York: Oxford University Press. pp. 41–61. ISBN 978-0-19-954586-5. OCLC 804498749.
- Knight, Chris (2009). Rudolf P Botha; Chris Knight (eds.). Language, Ochre, and the Rule of Law. The cradle of language. Oxford; New York: Oxford University Press. pp. 281–303. ISBN 978-0-19-954586-5. OCLC 804498749.
- Watts, Ian (2009). Rudolf P Botha; Chris Knight (eds.). Red Ochre, Body Painting, and Language: Interpreting the Blombos Ochre. The cradle of language. Oxford; New York: Oxford University Press. pp. 62–92. ISBN 978-0-19-954586-5. OCLC 804498749.
- Arcadi, AC. (August 2000). "Vocal responsiveness in male wild chimpanzees: implications for the evolution of language". Journal of Human Evolution. 39 (2): 205–23. doi:10.1006/jhev.2000.0415. PMID 10968929.
- Johanna Nichols, 1998. The origin and dispersal of languages: Linguistic evidence. In Nina Jablonski and Leslie C. Aiello, eds., The Origin and Diversification of Language, pp. 127–70. (Memoirs of the California Academy of Sciences, 24.) San Francisco: California Academy of Sciences.
- Perreault, C.; Mathew, S. (2012). "Dating the origin of language using phonemic diversity". PLOS ONE. 7 (4): e35289. Bibcode:2012PLoSO...735289P. doi:10.1371/journal.pone.0035289. PMC 3338724. PMID 22558135.
- Botha, Rudolf P.; Knight, Chris (2009). The cradle of language. Oxford; New York: Oxford University Press. ISBN 978-0-19-954586-5. OCLC 804498749.
- Darwin, C. (1871). "The Descent of Man, and Selection in Relation to Sex, 2 vols. London: Murray, p. 56.
- Müller, F. M. 1996 [1861]. The theoretical stage, and the origin of language. Lecture 9 from Lectures on the Science of Language. Reprinted in R. Harris (ed.), The Origin of Language. Bristol: Thoemmes Press, pp. 7–41.
- Paget, R. 1930. Human speech: some observations, experiments, and conclusions as to the nature, origin, purpose and possible improvement of human speech. London: Routledge & Kegan Paul.
- Firth, J. R. 1964. The Tongues of Men and Speech. London: Oxford University Press, pp. 25–6.
- Stam, J. H. 1976. Inquiries into the origins of language. New York: Harper and Row, p. 243-44.
- Zahavi, A. (May 1993). "The fallacy of conventional signalling". Philosophical Transactions of the Royal Society B: Biological Sciences. 340 (1292): 227–230. Bibcode:1993RSPTB.340..227Z. doi:10.1098/rstb.1993.0061. PMID 8101657.
- Smith, J.Maynard (1994). "Must reliable signals always be costly?". Animal Behaviour. 47 (5): 1115–1120. doi:10.1006/anbe.1994.1149. ISSN 0003-3472.
- Goodall, Jane (1986). The chimpanzees of Gombe : patterns of behavior. Cambridge, Massachusetts: Belknap Press of Harvard University Press. ISBN 978-0-674-11649-8. OCLC 12550961.
- Byrne, Richard W.; Whiten, Andrew. (1988). Machiavellian intelligence : social expertise and the evolution of intellect in monkeys, apes, and humans. Oxford: Clarendon Press. ISBN 978-0-19-852175-4. OCLC 17260831.
- de Waal, Frans B. M. (2005). "Intentional Deception in Primates". Evolutionary Anthropology. 1 (3): 86–92. doi:10.1002/evan.1360010306.
- Power, Camilla (1998). James R Hurford; Michael Studdert-Kennedy; Chris Knight (eds.). Old wives' tales: the gossip hypothesis and the reliability of cheap signals. Approaches to the evolution of language : social and cognitive base. Cambridge, UK; New York: Cambridge University Press. pp. 111–129. ISBN 978-0-521-63964-4. OCLC 37742390.
- Fitch, W. T. (2004). "Kin selection and "mother tongues": a neglected component in language evolution" (PDF). In Ulrike Griebel; D Kimbrough Oller (eds.). Evolution of communication systems : a comparative approach. Cambridge, Massachusetts: MIT Press. pp. 275–296. ISBN 978-0-262-15111-5. OCLC 845673575.
- Hamilton, W. D. (1964). "The genetical evolution of social behaviour. I, II". Journal of Theoretical Biology. 7 (1): 1–52. doi:10.1016/0022-5193(64)90038-4. PMID 5875341.
- Knight, Chris (13 November 2000). "Play as Precursor of Phonology and Syntax". The Evolutionary Emergence of Language. Cambridge University Press. pp. 99–120. doi:10.1017/cbo9780511606441.007. ISBN 978-0-521-78157-2.
- Tallerman, Maggie (2013). Rudolf P Botha; Martin Everaert (eds.). Kin selection, pedagogy and linguistic complexity: whence protolanguage?. The evolutionary emergence of language : evidence and inference. Oxford, UK: Oxford University Press. pp. 77–96. ISBN 978-0-19-965485-7. OCLC 856795812.
- Trivers, R. L. (1971). "The evolution of reciprocal altruism". Quarterly Review of Biology. 46: 35–57. doi:10.1086/406755.
- Dessalles, Jean L. (1998). James R Hurford; Michael Studdert-Kennedy; Chris Knight (eds.). Altruism, status and the origin of relevance. Approaches to the evolution of language : social and cognitive base. Cambridge, UK; New York: Cambridge University Press. pp. 130–147. ISBN 978-0-521-63964-4. OCLC 37742390.
- Dunbar, R. I. M. (1996). Grooming, gossip and the evolution of language. London: Faber and Faber. ISBN 9780571173969. OCLC 34546743.
- von Heiseler; Till Nikolaus (2014). "Language evolved for storytelling in a super-fast evolution". In R. L. C. Cartmill (ed.). Evolution of Language. London: World Scientific. pp. 114–121.
- Power, C. (1998). "Old wives' tales: the gossip hypothesis and the reliability of cheap signals". In J. R. Hurford; M. Studdert Kennedy; C. Knight (eds.). Approaches to the Evolution of Language: Social and Cognitive Bases. Cambridge: Cambridge University Press. pp. 111 29.
- Lewis, Jerome (2009). Rudolf P Botha; Chris Knight (eds.). As Well as Words: Congo Pygmy Hunting, Mimicry, and Play. The cradle of language. Oxford; New York: Oxford University Press. pp. 236–256. ISBN 978-0-19-954586-5. OCLC 804498749.
- Enfield, N. J. (2010). "Without social context?" (PDF). Science. 329 (5999): 1600–1601. Bibcode:2010Sci...329.1600E. doi:10.1126/science.1194229.
- Steels, L. (2009). Rudolf P. Botha; Chris Knight (eds.). Is sociality a crucial prerequisite for the emergence of language?. The prehistory of language. Oxford; New York: Oxford University Press. ISBN 978-0-19-954587-2. OCLC 819189595.
- Deacon, Terrence William (1997). The symbolic species : the co-evolution of language and the brain. New York: W.W. Norton. ISBN 978-0-393-03838-5. OCLC 807018262.
- Rappaport, Roy A. (1979). Ecology, Meaning, and Religion. Richmond, Calif.: North Atlantic Books. pp. 201–211. ISBN 978-0-913028-54-4. OCLC 397893672.
- Thomas, Neil; Bless, Josef J.; Alderson-Day, Ben; Bell, Imogen H.; Cella, Matteo; Craig, Tom; Delespaul, Philippe; Hugdahl, Kenneth; Laloyaux, Julien; Larøi, Frank; Lincoln, Tania M. (1 February 2019). "Potential Applications of Digital Technology in Assessment, Treatment, and Self-help for Hallucinations". Schizophrenia Bulletin. 45 (Supplement_1): S32–S42. doi:10.1093/schbul/sby103. ISSN 0586-7614. PMC 6357981. PMID 30715539.
- Searle, J. R. 1996. The Construction of Social Reality. London: Penguin.
- Durkheim, E. 1947 [1915]. "Origins of these beliefs". Chapter VII. In É. Durkheim, The Elementary Forms of the Religious Life: A study in religious sociology. Trans. J. W. Swain. Glencoe, Illinois: The Free Press, pp. 205–239.
- Chomsky, Noam (2011). "Language and Other Cognitive Systems. What is Special About Language?". Language Learning and Development. 7 (4): 263–78. doi:10.1080/15475441.2011.584041.
- Chomsky, N. (2005). "=Three factors in language design". Linguistic Inquiry. 36 (1): 1–22. doi:10.1162/0024389052993655.
- The multiple roles of cultural transmission experiments in understanding human cultural evolution, Alex Mesoudi, Andrew Whiten
- Chimpanzee Material Culture: Implications for Human Evolution, William Clement McGrew
- New Frontiers in Language Evolution and Development: Introduction to the topics Volume, D. Kimbrough Oller, Rick Dale, Ulrike Griebel
- Birdsong, speech, and language: exploring the evolution of mind and brain, JJ Bolhuis, M Everaert
- Arnauld, Antoine; Lancelot, Claude (1975) [First published 1660]. General and Rational Grammar : The Port-Royal Grammar. The Hague: Mouton. ISBN 902793004X.
- Daneš, František (1987). "On Prague school functionalism in linguistics". In Dirven, R.; Fried, V. (eds.). Functionalism in Linguistics. John Benjamins. pp. 3–38. ISBN 9789027215246.
- Aronoff, Mark (2017). "Darwinism tested by the science of language". In Bowern; Horn; Zanuttini (eds.). On Looking into Words (and Beyond): Structures, Relations, Analyses. SUNY Press. pp. 443–456. ISBN 978-3-946234-92-0. Retrieved 3 March 2020.
- Hauser, Mark D.; Yang, Charles; Berwick, Robert C.; Tattersall, Ian; Ryan, Michael J.; Watumull, Jeffrey; Chomsky, Noam; Lewontin, Richard C. (2014). "The mystery of language evolution". Frontiers in Psychology. 5: 401. doi:10.3389/fpsyg.2014.00401x (inactive 23 May 2020). PMC 4019876. PMID 24847300.
- Anderson, Earl R. (1998). A Grammar of Iconism. Fairleigh Dickinson University Press. ISBN 9780838637647.
- de Saussure, Ferdinand (1959) [First published 1916]. Course in general linguistics (PDF). New York: Philosophy Library. ISBN 9780231157278.
- Lévi-Strauss, Claude (1987). Introduction to the work of Marcel Mauss. Routledge & Kegan Paul Ltd. pp. 59–60. ISBN 0-7100-9066-8.
- Hejl, P. M. (2013). "The importance of the concepts of "organism" and "evolution" in Emile Durkheim's division of social labor and the influence of Herbert Spencer". In Maasen, Sabine; Mendelsohn, E.; Weingart, P. (eds.). Biology as Society, Society as Biology: Metaphors. Springer. pp. 155–191. ISBN 9789401106733.
- Chomsky, N. (2004). Language and Mind: Current thoughts on ancient problems. Part I & Part II. In Lyle Jenkins (ed.), Variation and Universals in Biolinguistics. Amsterdam: Elsevier, pp. 379–405.
- Berwick, Robert; Chomsky, Noam (2016). Why Only Us: Language and Evolution. Cambridge, Massachusetts: MIT Press. ISBN 9780262034241. OCLC 934433881.
- Collins, Brian (22 September 2019). "What is recursion/recursiveness?".
- Dediu, Dan; Levinson, Stephen C. (2013). "On the antiquity of language: the reinterpretation of Neandertal linguistic capacities and its consequences". Frontiers in Psychology. 4: 397. doi:10.3389/fpsyg.2013.00397. ISSN 1664-1078. PMC 3701805. PMID 23847571.
- Vyshedskiy, Andrey (29 July 2019). "Language evolution to revolution: the leap from rich-vocabulary non-recursive communication system to recursive language 70,000 years ago was associated with acquisition of a novel component of imagination, called Prefrontal Synthesis, enabled by a mutation that slowed down the prefrontal cortex maturation simultaneously in two or more children – the Romulus and Remus hypothesis". Research Ideas and Outcomes. 5. doi:10.3897/rio.5.e38546. ISSN 2367-7163.
- Bakker, Peter (July 1987). "Autonomous Languages of Twins". Acta Geneticae Medicae et Gemellologiae: Twin Research. 36 (2): 233–238. doi:10.1017/S0001566000004463. ISSN 0001-5660.
- Amos W.; Hoffman J. I. (7 January 2010). "Evidence that two main bottleneck events shaped modern human genetic diversity". Proceedings of the Royal Society B: Biological Sciences. 277 (1678): 131–137. doi:10.1098/rspb.2009.1473. PMC 2842629. PMID 19812086.
- Premack, David & Premack, Ann James. The Mind of an Ape, ISBN 0-393-01581-5.
- Kimura, Doreen (1993). Neuromotor mechanisms in human communication. New York: Oxford University Press. ISBN 978-0-19-505492-7. OCLC 26396505.
- Newman, A. J.; et al. (2002). "A Critical Period for Right Hemisphere Recruitment in American Sign Language Processing". Nature Neuroscience. 5 (1): 76–80. doi:10.1038/nn775. PMID 11753419.
- Arbib MA; Liebal, K; Pika, S (December 2008). "Primate vocalization, gesture, and the evolution of human language". Current Anthropology. 49 (6): 1053–63, discussion 1063–76. doi:10.1086/593015. PMID 19391445.
- Capone, Nina C.; McGregor, Karla K. (2004). "Gesture Development". Journal of Speech, Language, and Hearing Research. 47 (1): 173–86. doi:10.1044/1092-4388(2004/015). PMID 15072537.
- Ozçalişkan S; Goldin-Meadow S (July 2005). "Gesture is at the cutting edge of early language development". Cognition. 96 (3): B101–13. doi:10.1016/j.cognition.2005.01.001. PMID 15996556.
- Rizzolatti, G. (2008). Giacomo Rizzolatti on the Evolution of Language. Retrieved from http://gocognitive.net/interviews/evolution-language-gestures%5B%5D
- Corballis, Michael C. (2002). Alison Wray (ed.). The transition to language. Oxford: Oxford University Press. pp. 161–179. ISBN 978-0-19-925066-0. OCLC 48532303.
- Knight, Chris (2006). Angelo Cangelosi; Andrew D M Smith; Kenny Smith (eds.). Language co-evolved with the rule of law (PDF). The evolution of language : proceedings of the 6th international conference (EVOLANG6), Rome, Italy, 12–15 April 200. 7. New Jersey: World Scientific. pp. 109–128. doi:10.1007/s11299-007-0039-1. ISBN 9789812566560. OCLC 70797781.
- Knight, Chris (2000). Chris Knight; Michael Studdert-Kennedy; James R Hurford (eds.). Play as precursor of phonology and syntax. The Evolutionary emergence of language : social function and the origins of linguistic for. Cambridge; New York: Cambridge University Press. pp. 99–1119. ISBN 978-0-521-78157-2. OCLC 807262339.
- Kolb, Bryan & Ian Q. Whishaw (2003). Fundamentals of Human Neuropsychology (5th ed.). Worth Publishers. ISBN 978-0-7167-5300-1.
- Larsson, M (2015). "Tool-use-associated sound in the evolution of language". Animal Cognition. 18 (5): 993–1005. doi:10.1007/s10071-015-0885-x. PMID 26118672.
- Larsson, M (2014). "Self-generated sounds of locomotion and ventilation and the evolution of human rhythmic abilities". Animal Cognition. 17 (1): 1–14. doi:10.1007/s10071-013-0678-z. PMC 3889703. PMID 23990063.
- Skoyles, John R., Gesture, Language Origins, and Right Handedness, Psycholoqy: 11,#24, 2000
- Petrides, M.; Cadoret, G.; Mackey, S. (June 2005). "Orofacial somatomotor responses in the macaque monkey homologue of Broca's area". Nature. 435 (7046): 1235–8. Bibcode:2005Natur.435.1235P. doi:10.1038/nature03628. PMID 15988526. S2CID 4397762.
- Porter, RJ.; Lubker, JF. (September 1980). "Rapid reproduction of vowel-vowel sequences: evidence for a fast and direct acoustic-motoric linkage in speech". Journal of Speech and Hearing Research. 23 (3): 593–602. doi:10.1044/jshr.2303.593. PMID 7421161.
- McCarthy, R.; Warrington, EK. (June 1984). "A two-route model of speech production. Evidence from aphasia". Brain. 107 (2): 463–85. doi:10.1093/brain/107.2.463. PMID 6722512.
- McCarthy, RA.; Warrington, EK. (2001). "Repeating without semantics: surface dysphasia?". Neurocase. 7 (1): 77–87. doi:10.1093/neucas/7.1.77. PMID 11239078. S2CID 12988855.
- Marslen-Wilson, W. (1973). "Linguistic structure and speech shadowing at very short latencies". Nature. 244 (5417): 522–523. Bibcode:1973Natur.244..522M. doi:10.1038/244522a0. PMID 4621131. S2CID 4220775.
- Fay, WH.; Coleman, RO. (July 1977). "A human sound transducer/reproducer: temporal capabilities of a profoundly echolalic child". Brain and Language. 4 (3): 396–402. doi:10.1016/0093-934x(77)90034-7. PMID 907878.
- Schippers, MB; Roebroeck, A; Renken, R; Nanetti, L; Keysers, C (2010). "Mapping the Information flow from one brain to another during gestural communication". Proceedings of the National Academy of Sciences of the United States of America. 107 (20): 9388–93. Bibcode:2010PNAS..107.9388S. doi:10.1073/pnas.1001791107. PMC 2889063. PMID 20439736.
- Moro, Andrea (2008). The boundaries of Babel : the brain and the enigma of impossible language. Cambridge, Massachusetts: MIT Press. ISBN 978-0-262-13498-9. OCLC 804408004.
- Falk, D. (August 2004). "Prelinguistic evolution in early hominins: whence motherese?" (PDF). Behavioral and Brain Sciences. 27 (4): 491–503, discussion 503–83. doi:10.1017/s0140525x04000111. PMID 15773427. Archived from the original (PDF) on 4 January 2014. Retrieved 4 January 2014.
- Kaye, K. (1982). The Mental and Social Life of Babies. Univ. Chicago Press. pp. 186. ISBN 0226428486.
- Poliva, Oren (20 September 2017). "From where to what: a neuroanatomically based evolutionary model of the emergence of speech in humans". F1000Research. 4: 67. doi:10.12688/f1000research.6175.3. ISSN 2046-1402. PMC 5600004. PMID 28928931.
- Poliva, Oren (30 June 2016). "From Mimicry to Language: A Neuroanatomically Based Evolutionary Model of the Emergence of Vocal Language". Frontiers in Neuroscience. 10: 307. doi:10.3389/fnins.2016.00307. ISSN 1662-453X. PMC 4928493. PMID 27445676.
- Scott, S. K. (1 December 2000). "Identification of a pathway for intelligible speech in the left temporal lobe". Brain. 123 (12): 2400–2406. doi:10.1093/brain/123.12.2400. ISSN 1460-2156. PMC 5630088. PMID 11099443.
- Davis, Matthew H.; Johnsrude, Ingrid S. (15 April 2003). "Hierarchical Processing in Spoken Language Comprehension". The Journal of Neuroscience. 23 (8): 3423–3431. doi:10.1523/jneurosci.23-08-03423.2003. ISSN 0270-6474. PMC 6742313. PMID 12716950.
- Petkov, Christopher I; Kayser, Christoph; Steudel, Thomas; Whittingstall, Kevin; Augath, Mark; Logothetis, Nikos K (10 February 2008). "A voice region in the monkey brain". Nature Neuroscience. 11 (3): 367–374. doi:10.1038/nn2043. ISSN 1097-6256. PMID 18264095.
- Buchsbaum, Bradley R.; Baldo, Juliana; Okada, Kayoko; Berman, Karen F.; Dronkers, Nina; D'Esposito, Mark; Hickok, Gregory (December 2011). "Conduction aphasia, sensory-motor integration, and phonological short-term memory – An aggregate analysis of lesion and fMRI data". Brain and Language. 119 (3): 119–128. doi:10.1016/j.bandl.2010.12.001. ISSN 0093-934X. PMC 3090694. PMID 21256582.
- Warren, Jane E.; Wise, Richard J.S.; Warren, Jason D. (December 2005). "Sounds do-able: auditory–motor transformations and the posterior temporal plane". Trends in Neurosciences. 28 (12): 636–643. doi:10.1016/j.tins.2005.09.010. ISSN 0166-2236. PMID 16216346.
- Campbell, Ruth (12 March 2008). "The processing of audio-visual speech: empirical and neural bases". Philosophical Transactions of the Royal Society of London B: Biological Sciences. 363 (1493): 1001–1010. doi:10.1098/rstb.2007.2155. ISSN 0962-8436. PMC 2606792. PMID 17827105.
- Kayser, Christoph; Petkov, Christopher I.; Logothetis, Nikos K. (December 2009). "Multisensory interactions in primate auditory cortex: fMRI and electrophysiology". Hearing Research. 258 (1–2): 80–88. doi:10.1016/j.heares.2009.02.011. ISSN 0378-5955. PMID 19269312.
- Hickok, Gregory; Buchsbaum, Bradley; Humphries, Colin; Muftuler, Tugan (1 July 2003). "Auditory–Motor Interaction Revealed by fMRI: Speech, Music, and Working Memory in Area Spt". Journal of Cognitive Neuroscience. 15 (5): 673–682. doi:10.1162/089892903322307393. ISSN 1530-8898. PMID 12965041.
- Schwartz, M. F.; Faseyitan, O.; Kim, J.; Coslett, H. B. (20 November 2012). "The dorsal stream contribution to phonological retrieval in object naming". Brain. 135 (12): 3799–3814. doi:10.1093/brain/aws300. ISSN 0006-8950. PMC 3525060. PMID 23171662.
- Gow, David W. (June 2012). "The cortical organization of lexical knowledge: A dual lexicon model of spoken language processing". Brain and Language. 121 (3): 273–288. doi:10.1016/j.bandl.2012.03.005. ISSN 0093-934X. PMC 3348354. PMID 22498237.
- Buchsbaum, Bradley R.; D'Esposito, Mark (May 2008). "The Search for the Phonological Store: From Loop to Convolution". Journal of Cognitive Neuroscience. 20 (5): 762–778. doi:10.1162/jocn.2008.20501. ISSN 0898-929X. PMID 18201133.
- Lachaux, Jean-Philippe; Jerbi, Karim; Bertrand, Olivier; Minotti, Lorella; Hoffmann, Dominique; Schoendorff, Benjamin; Kahane, Philippe (31 October 2007). "A Blueprint for Real-Time Functional Mapping via Human Intracranial Recordings". PLOS ONE. 2 (10): e1094. Bibcode:2007PLoSO...2.1094L. doi:10.1371/journal.pone.0001094. ISSN 1932-6203. PMC 2040217. PMID 17971857.
- Jardri, Renaud; Houfflin-Debarge, Véronique; Delion, Pierre; Pruvo, Jean-Pierre; Thomas, Pierre; Pins, Delphine (April 2012). "Assessing fetal response to maternal speech using a noninvasive functional brain imaging technique". International Journal of Developmental Neuroscience. 30 (2): 159–161. doi:10.1016/j.ijdevneu.2011.11.002. ISSN 0736-5748. PMID 22123457.
- Petersen, M.; Beecher, M.; Zoloth; Moody, D.; Stebbins, W. (20 October 1978). "Neural lateralization of species-specific vocalizations by Japanese macaques (Macaca fuscata)". Science. 202 (4365): 324–327. Bibcode:1978Sci...202..324P. doi:10.1126/science.99817. ISSN 0036-8075. PMID 99817.
- Heffner, H.; Heffner, R. (5 October 1984). "Temporal lobe lesions and perception of species-specific vocalizations by macaques". Science. 226 (4670): 75–76. Bibcode:1984Sci...226...75H. doi:10.1126/science.6474192. ISSN 0036-8075. PMID 6474192.
- Shu, W.; Cho, J. Y.; Jiang, Y.; Zhang, M.; Weisz, D.; Elder, G. A.; Schmeidler, J.; De Gasperi, R.; Sosa, M. A. G. (27 June 2005). "Altered ultrasonic vocalization in mice with a disruption in the Foxp2 gene". Proceedings of the National Academy of Sciences. 102 (27): 9643–9648. Bibcode:2005PNAS..102.9643S. doi:10.1073/pnas.0503739102. ISSN 0027-8424. PMC 1160518. PMID 15983371.
- Sia, G. M.; Clem, R. L.; Huganir, R. L. (31 October 2013). "The Human Language-Associated Gene SRPX2 Regulates Synapse Formation and Vocalization in Mice". Science. 342 (6161): 987–991. Bibcode:2013Sci...342..987S. doi:10.1126/science.1245079. ISSN 0036-8075. PMC 3903157. PMID 24179158.
- Sperber, D. and D. Wilson 1986. Relevance. Communication and cognition. Oxford: Blackwell.
- Deutscher, Guy (2005). The unfolding of language : an evolutionary tour of mankind's greatest invention. New York: Metropolitan Books. ISBN 978-0-8050-7907-4. OCLC 57311730.
- Hopper, P. J. 1998. Emergent grammar. In M. Tomasello (ed.), The New Psychology of Language. Mahwah, NJ: Lawrence Erlbaum, 155–175.
- Heine, Bernd; Kuteva, Tania (2007). The genesis of grammar : a reconstructio. Oxford; New York: Oxford University Press. ISBN 978-0-19-922777-8. OCLC 849464326.
- Lakoff, G. and M. Johnson 1980. Metaphors We Live By. Chicago: University of Chicago Press.
- Heine, Bernd; Kuteva, Tania (2012). Maggie Tallerman; Kathleen R. Gibson (eds.). Grammaticalization theory as a tool for reconstructing language evolution. The Oxford handbook of language evolution. Oxford; New York: Oxford University Press. pp. 512–527. ISBN 978-0-19-954111-9. OCLC 724665645.
- Cheney, Dorothy L.; Seyfarth, Robert M. (2005). "Constraints and preadaptations in the earliest stages of language evolution" (PDF). The Linguistic Review. 22 (2–4): 135–59. doi:10.1515/tlir.2005.22.2-4.135.
- Maynard Smith, John; Harper, David (2003). Animal signals. New York: Oxford University Press. ISBN 978-0-19-852684-1. OCLC 54460090.
- Davidson, R. D. 1979. What metaphors mean. In S. Sacks (ed.), On Metaphor. Chicago: University of Chicago Press, pp. 29–45.
- Lakoff, G. and R. Núñez 2000. Where mathematics comes from. New York: Basic Books.
- Gallup, GG Jr. (1970). "Chimpanzees: Self recognition". Science. 167 (3914): 86–87. Bibcode:1970Sci...167...86G. doi:10.1126/science.167.3914.86. PMID 4982211.
- Mitchell, R.W. (1995). "Evidence of dolphin self-recognition and the difficulties of interpretation". Consciousness and Cognition. 4 (2): 229–234. doi:10.1006/ccog.1995.1029. PMID 8521261.
- Ko, Kwang Hyun (2016). "Origins of human intelligence: The chain of tool-making and brain evolution" (PDF). Anthropological Notebooks. 22 (1): 5–22.
- Soma, M; Hiraiwa-Hasegawa, M; Okanoya, K (2009). "Early ontogenetic effects on song quality in the Bengalese finch (Lonchura striata var. domestica): laying order, sibling competition and song syntax". Behavioral Ecology and Sociobiology. 63 (3): 363–370. doi:10.1007/s00265-008-0670-9.
- Graham Ritchie & Simon Kirby (2005). "Selection, domestication, and the emergence of learned communication systems" (PDF). Second International Symposium on the Emergence and Evolution of Linguistic Communication. Archived from the original (PDF) on 21 January 2012.
- MacNeilage, P. 1998. Evolution of the mechanism of language output: comparative neurobiology of vocal and manual communication. In J. R. Hurford, M. Studdert Kennedy and C. Knight (eds), Approaches to the Evolution of Language. Cambridge: Cambridge University Press, pp. 222 41.
- Hauser, MD.; Chomsky, N.; Fitch, WT. (November 2002). "The faculty of language: what is it, who has it, and how did it evolve?" (PDF). Science. 298 (5598): 1569–79. doi:10.1126/science.298.5598.1569. PMID 12446899. Archived from the original (PDF) on 28 December 2013.
- Everett, Daniel L. (2005). "Cultural Constraints on Grammar and Cognition in Piraha Another Look at the Design Features of Human Language" (PDF). Current Anthropology. 46 (4): 621–646. doi:10.1086/431525. hdl:2066/41103.
- Zhordania, I. M. (2006). Who asked the first question : the origins of human choral singing, intelligence, language and speech. Tbilisi, Georgia: Logos Tbilisi Ivane Javakhishvili State University. ISBN 9789994031818. OCLC 224993377.
- Bolinger, Dwight L. (Editor) 1972. Intonation. Selected Readings. Harmondsworth: Penguin, pg.314
- Cruttenden, Alan (1986). Intonation. Cambridge [Cambridgeshire]; New York: Cambridge University Press. pp. 169–174. ISBN 978-0-521-26028-2. OCLC 12103838.
- Carey, Susan (2001). "Cognitive Foundations of Arithmetic: Evolution and Ontogenisis" (PDF). Mind and Language. 16 (1): 37–55. doi:10.1111/1468-0017.00155. Archived from the original (PDF) on 25 July 2013. Retrieved 13 January 2014.
- Hockett, Charles F. (1960). "The Origin of Speech" (PDF). Scientific American. 203 (3): 88–96. Bibcode:1960SciAm.203c..88H. doi:10.1038/scientificamerican0960-88. Archived from the original (PDF) on 6 January 2014. Retrieved 6 January 2014.
- Diamond, Jared M. (1992). "Bridges to human language". The third chimpanzee : the evolution and future of the human animal. New York: HarperCollins. pp. 141–167. ISBN 978-0-06-018307-3. OCLC 24246928.
- Savage-Rumbaugh, E. Sue; Lewin, Roger. (1994). Kanzi : the ape at the brink of the human mind. New York: Wiley. ISBN 978-0-471-58591-6. OCLC 30112573.
- Savage-Rumbaugh, E. Sue; Shanker, Stuart.; Taylor, Talbot J. (1998). Apes, language, and the human mind. New York: Oxford University Press. ISBN 978-0-19-510986-3. OCLC 38566026.
- Freeman, Scott; Jon C. Herron., Evolutionary Analysis (4th ed.), Pearson Education, Inc. (2007), ISBN 0-13-227584-8 pages 789–90
- Seyfarth, Robert M.; Cheney, Dorothy L.; Marler, Peter (1980). "Vervet monkey alarm calls: Semantic communication in a free-ranging primate". Animal Behaviour. 28 (4): 1070–1094. doi:10.1016/S0003-3472(80)80097-2.
- Arnold, Kate; Zuberbühler, Klaus (2006). "Language evolution: Semantic combinations in primate calls". Nature. 441 (7091): 303. Bibcode:2006Natur.441..303A. doi:10.1038/441303a. PMID 16710411.
- Wade, Nicholas (23 May 2006). "Nigerian Monkeys Drop Hints on Language Origin". The New York Times. Retrieved 9 September 2007.
- Gibbons, Christopher M. (2007). The referentiality of chimpanzee vocal signaling: behavioral and acoustic analysis of food barks (Thesis). Ohio State University.
- Slocombe, Katie E.; Zuberbühler, Klaus (2005). "Functionally Referential Communication in a Chimpanzee". Current Biology. 15 (19): 1779–1784. doi:10.1016/j.cub.2005.08.068. PMID 16213827.
- Clark, Gary; Henneberg, Maciej (2017). "Ardipithecus ramidus and the evolution of language and singing: An early origin for hominin vocal capability". HOMO. 68 (2): 101–121. doi:10.1016/j.jchb.2017.03.001. PMID 28363458.
- Aronoff, Mark; Rees-Miller, Janie. (2001). The handbook of linguistics. Malden, Mass.: Blackwell Publishers. ISBN 0631204970. OCLC 43115110.
- Fitch, W.Tecumseh (2000). "The evolution of speech: a comparative review". Trends in Cognitive Sciences. 4 (7): 258–267. doi:10.1016/S1364-6613(00)01494-7. PMID 10859570.
- Ohala, John J. (10 September 1987). "Experimental Phonology". Annual Meeting of the Berkeley Linguistics Society. 13: 207. doi:10.3765/bls.v13i0.1803. ISSN 2377-1666.
- "APA Upgrades APA PsycNET Content Delivery Platform". American Psychological Association. 2017. doi:10.1037/e500792018-001. Cite journal requires
|journal=(help) - Mithen, Steven J. (2006). The singing neanderthals : the origins of music, language, mind, and body. Cambridge, Massachusetts: Harvard University Press. ISBN 978-0-674-02192-1. OCLC 62090869.
- Arensburg, B.; Schepartz, LA.; Tillier, AM.; Vandermeersch, B.; Rak, Y. (October 1990). "A reappraisal of the anatomical basis for speech in Middle Palaeolithic hominids". American Journal of Physical Anthropology. 83 (2): 137–46. doi:10.1002/ajpa.1330830202. PMID 2248373.
- D'Anastasio, R.; Wroe, S.; Tuniz, C.; Mancini, L.; Cesana, DT.; Dreossi, D.; Ravichandiran, M.; Attard, M.; et al. (2013). "Micro-biomechanics of the kebara 2 hyoid and its implications for speech in neanderthals". PLOS ONE. 8 (12): e82261. Bibcode:2013PLoSO...882261D. doi:10.1371/journal.pone.0082261. PMC 3867335. PMID 24367509.
- Jungers, WL.; Pokempner, AA.; Kay, RF.; Cartmill, M. (August 2003). "Hypoglossal canal size in living hominoids and the evolution of human speech" (PDF). Human Biology. 75 (4): 473–84. doi:10.1353/hub.2003.0057. PMID 14655872. Archived from the original (PDF) on 12 June 2007.
- DeGusta, D.; Gilbert, WH.; Turner, SP. (February 1999). "Hypoglossal canal size and hominid speech". Proceedings of the National Academy of Sciences of the United States of America. 96 (4): 1800–4. Bibcode:1999PNAS...96.1800D. doi:10.1073/pnas.96.4.1800. PMC 15600. PMID 9990105.
- Johansson, Sverker (April 2006). "Constraining the Time When Language Evolved" (PDF). Evolution of Language: Sixth International Conference, Rome: 152–159. doi:10.1142/9789812774262_0020. ISBN 9789812566560. Archived from the original (PDF) on 15 October 2006. Retrieved 10 September 2007.
- Houghton, P. (February 1993). "Neandertal supralaryngeal vocal tract". American Journal of Physical Anthropology. 90 (2): 139–46. doi:10.1002/ajpa.1330900202. PMID 8430750.
- Boë, Louis-Jean; Maeda, Shinji; Heim, Jean-Louis (1999). "Neandertal man was not morphologically handicapped for speech". Evolution of Communication. 3 (1): 49–77. doi:10.1075/eoc.3.1.05boe.
- de Boer, Bart (2017). "Evolution of speech and evolution of language". Psychonomic Bulletin & Review. 24 (1): 158–162. doi:10.3758/s13423-016-1130-6. ISSN 1069-9384.
- Klarreich, E. (2004). "Biography of Richard G. Klein". Proceedings of the National Academy of Sciences. 101 (16): 5705–7. Bibcode:2004PNAS..101.5705K. doi:10.1073/pnas.0402190101. PMC 395972. PMID 15079069.
- Klein, Richard G. "Three Distinct Human Populations". Biological and Behavioral Origins of Modern Humans. Access Excellence @ The National Health Museum. Retrieved 10 September 2007.
- Marwick, Ben (2003). "Pleistocene Exchange Networks as Evidence for the Evolution of Language". Cambridge Archaeological Journal. 13: 67–81. doi:10.1017/S0959774303000040. hdl:1885/42089.
- Fleagle, John G.; Assefa, Zelalem; Brown, Francis H.; Shea, John J. (2008). "Paleoanthropology of the Kibish Formation, southern Ethiopia: Introduction". Journal of Human Evolution. 55 (3): 360–5. doi:10.1016/j.jhevol.2008.05.007. PMID 18617219.
- Henshilwood, C. S.; d'Errico, F.; Yates, R.; Jacobs, Z.; Tribolo, C.; Duller, G. A. T.; Mercier, N.; Sealy, J. C.; Valladas, H.; Watts, I.; Wintle, A. G. (2002). "Emergence of modern human behavior: Middle Stone Age engravings from South Africa". Science. 295 (5558): 1278–1280. Bibcode:2002Sci...295.1278H. doi:10.1126/science.1067575. PMID 11786608.
- Minkel, J. R. (18 July 2007). "Skulls Add to "Out of Africa" Theory of Human Origins: Pattern of skull variation bolsters the case that humans took over from earlier species". Scientific American.com. Retrieved 9 September 2007.
- Chris Stringer, 2011. The Origin of Our Species. London: Penguin.
- Fitch, WT. (2000). "The phonetic potential of nonhuman vocal tracts: comparative cineradiographic observations of vocalizing animals". Phonetica. 57 (2–4): 205–18. doi:10.1159/000028474. PMID 10992141.
- Fitch, WT.; Reby, D. (August 2001). "The descended larynx is not uniquely human". Proceedings of the Royal Society B. 268 (1477): 1669–75. doi:10.1098/rspb.2001.1704. PMC 1088793. PMID 11506679.
- Weissengruber, GE.; Forstenpointner, G.; Peters, G.; Kübber-Heiss, A.; Fitch, WT. (September 2002). "Hyoid apparatus and pharynx in the lion (Panthera leo), jaguar (Panthera onca), tiger (Panthera tigris), cheetah (Acinonyxjubatus) and domestic cat (Felis silvestris f. catus)". Journal of Anatomy. 201 (3): 195–209. doi:10.1046/j.1469-7580.2002.00088.x. PMC 1570911. PMID 12363272.
- Lieberman, Philip (2007). "The Evolution of Human Speech: Its Anatomical and Neural Bases" (PDF). Current Anthropology. 48 (1): 39–66. doi:10.1086/509092.
- Nishimura, T.; Mikami, A.; Suzuki, J.; Matsuzawa, T. (September 2006). "Descent of the hyoid in chimpanzees: evolution of face flattening and speech". Journal of Human Evolution. 51 (3): 244–54. doi:10.1016/j.jhevol.2006.03.005. PMID 16730049.
- M. Clegg 2001. The Comparative Anatomy and Evolution of the Human Vocal Tract Unpublished thesis, University of London.
- John J. Ohala, 2000. The irrelevance of the lowered larynx in modern Man for the development of speech. Paris, ENST: The Evolution of Language, pp. 171–172.
- Fitch, W. T. (2002). Comparative vocal production and the evolution of speech: Reinterpreting the descent of the larynx. In A. Wray (ed.), The Transition to Language. Oxford: Oxford University Press, pp. 21–45.
- Turner, P. and Russell-Coulter, C. (2001) Dictionary of Ancient Deities (Oxford: OUP)
- Pennock, Robert T. (2000). Tower of Babel: The Evidence against the New Creationism. Bradford Books. ISBN 9780262661652.
- Lindsay, Robert (1728). The history of Scotland: from 21 February 1436. to March, 1565. In which are contained accounts of many remarkable passages altogether differing from our other historians; and many facts are related, either concealed by some, or omitted by others. Baskett and company. p. 104.
Further reading
- Allott, Robin. (1989). The motor theory of language origin. Sussex, England: Book Guild. ISBN 978-0-86332-359-1. OCLC 21874255.
- Armstrong, David F.; Stokoe, William C.; Wilcox, Sherman E. (1995). Gesture and the nature of language. Cambridge: Cambridge University Press. ISBN 978-0-52-146772-8.
- Botha, Rudolf P.; Knight, Chris (2009). The prehistory of language. Oxford; New York: Oxford University Press. ISBN 978-0-19-954587-2. OCLC 819189595.
- Burling, Robbins (2005). The talking ape : how language evolved. Oxford; New York: Oxford University Press. ISBN 978-0-19-927940-1. OCLC 750809912.
- Cangelosi, Angelo; Greco, Alberto; Harnad, Stevan (2002). Angelo Cangelosi; Domenico Parisi (eds.). Symbol grounding and the symbolic theft hypothesis. Simulating the evolution of language. London; New York: Springer. ISBN 978-1-85233-428-4. OCLC 47824669.
- Christiansen, Morten H. (2013). Rudolf P Botha; Martin Everaert (eds.). Language has evolved to depend on multiple-cue integration. The evolutionary emergence of language : evidence and inferenc. Oxford, UK: Oxford University Press. ISBN 978-0-19-965484-0. OCLC 828055639.
- Corballis, Michael C. (2002). From hand to mouth : the origins of language. Princeton: Princeton University Press. ISBN 978-0-691-08803-7. OCLC 469431753.
- Crystal, David (1997). The Cambridge encyclopedia of language. Cambridge; New York: Cambridge University Press. ISBN 978-0-521-55967-6. OCLC 34704876.
- de Grolier, E. (ed.), 1983. The Origin and Evolution of Language. Paris: Harwood Academic Publishers.
- Dessalles, J-L., 2007. Why We Talk. The evolutionary origins of language. Oxford: Oxford University Press.
- Dor, Dan; Knight, Chris; Lewis, Jerome (2015). The Social Origins of Language. Oxford: Oxford University Press. ISBN 978-0-19-966533-4.
- Dunbar, R. I. M. (Robin Ian MacDonald); Knight, Chris; Power, Camilla (1999). The evolution of culture : an interdisciplinary view. Edinburgh: Edinburgh University Press. ISBN 978-0-7486-1076-1. OCLC 807340111.
- Fitch, W. Tecumseh (2010). The Evolution of Language. Cambridge: Cambridge. ISBN 978-0-521-67736-3. OCLC 428024376.
- Givón, Talmy; Malle, Bertram F (2002). The evolution of language out of pre-language. Amsterdam; Philadelphia: J. Benjamins Pub. ISBN 978-1-58811-237-8. OCLC 223393453.
- Harnad, Stevan R. (1976). Steklis, Horst D.; Lancaster, Jane (eds.). Origins and Evolution of Language and Speech. Annals of the New York Academy of Sciences, v. 280. New York: New York Academy of Sciences. ISBN 0-89072-026-6. OCLC 2493424.
- Hillert, Dieter (2014). The Nature of Language. Evolution, Paradigms and Circuits. New York: Springer Nature. ISBN 978-1-4939-0609-3.
- Hurford, James R (1990). I. M. Roca (ed.). Nativist and functional explanations in language acquisition (PDF). Logical issues in language acquisition. Dordrecht, Holland Providence, R.I: Foris Publications. ISBN 9789067655064. OCLC 832515162.
- Hurford, James R. (2007). The origins of meaning – 1 The origins of meaning. Oxford; New York: Oxford University Press. ISBN 978-0-19-920785-5. OCLC 263645256.
- Hurford, James R.; Studdert-Kennedy, Michael.; Knight, Chris (1998). Approaches to the evolution of language : social and cognitive bases. Cambridge, UK; New York: Cambridge University Press. ISBN 978-0-521-63964-4. OCLC 37742390.
- Kenneally, Christine. (2007). The first word : the search for the origins of language. New York: Viking. ISBN 978-0-670-03490-1. OCLC 80460757.
- Knight, Chris (2016). "Puzzles and mysteries in the origin of language" (PDF). Language and Communication. 50: 12–21. doi:10.1016/j.langcom.2016.09.002.
- Knight, Chris (2016). Decoding Chomsky: Science and revolutionary politics. New Haven & London: Yale University Press. ISBN 978-0-300-22146-6.
- Knight, Chris; Studdert-Kennedy, Michael.; Hurford, James R. (2000). The Evolutionary emergence of language : social function and the origins of linguistic form. Cambridge; New York: Cambridge University Press. ISBN 978-0-521-78157-2. OCLC 807262339.
- Komarova, Natalia L. (2006). L E Grinin; Victor C De Munck; A V Korotaev; Rossiĭskiĭ gosudarstvennyĭ gumanitarnyĭ universitet. (eds.). Language and Mathematics: An evolutionary model of grammatical communication. History and mathematics. Analyzing and modeling global development. [Moskva]: URSS. pp. 164–179. ISBN 978-5-484-01001-1. OCLC 182730511.
- Laitman, Jeffrey T.; Reidenberg. Joy S (2009). Marvin P Fried; Alfio Ferlito (eds.). Evolution of the Human Larynx: Nature's Great Experiment. The larynx. San Diego: Plural Pub. pp. 19–38. ISBN 9781597560627. OCLC 183609898.
- Lenneberg, E. H. 1967. Biological Foundations of Language. New York: Wiley.
- Leroi-Gourhan, A. 1993. Gesture and Speech. Trans. A. Bostock Berger. Cambridge, Massachusetts: MIT Press.
- Lieberman, Philip. (1991). Uniquely human : the evolution of speech, thought, and selfless behavior. Cambridge, Massachusetts: Harvard University Press. ISBN 978-0-674-92182-5. OCLC 21764294.
- Lieberman, P. (2007). "The evolution of human speech: Its anatomical and neural bases" (PDF). Current Anthropology. 48 (1): 39–66. doi:10.1086/509092.
- Lieberman, Philip. (2006). Toward an evolutionary biology of language. Cambridge, Massachusetts: Belknap Press of Harvard University Press. ISBN 978-0-674-02184-6. OCLC 62766735.
- Lieberman, Philip; McCarthy, Robert C.; Strait, David (2006). "The recent origin of human speech". The Journal of the Acoustical Society of America. 119 (5): 3441. Bibcode:2006ASAJ..119.3441L. doi:10.1121/1.4786937.
- Logan, Robert K. 2007. "The Extended Mind: The Emergence of Language, the Human Mind and Culture. Toronto: University of Toronto Press.
- MacNeilage, P. 2008. The Origin of Speech. Oxford: Oxford University Press.
- Mazlumyan, Victoria 2008. Origins of Language and Thought. ISBN 0977391515.
- Mithen, Stephen 2006. The Singing Neanderthals: The Origins of Music, Language, Mind and Body. ISBN 9780753820513
- Ostler, Nicholas. (2005). Empires of the word : a language history of the world. New York: HarperCollins Publishers. ISBN 978-0-06-621086-5. OCLC 58563178.
- Pinker, Steven (2007). The language instinct : how the mind creates language. New York: HarperPerennial ModernClassics. ISBN 9780061336461. OCLC 672454779.
- Pollick, AS.; de Waal, FB. (May 2007). "Ape gestures and language evolution". Proceedings of the National Academy of Sciences of the United States of America. 104 (19): 8184–9. Bibcode:2007PNAS..104.8184P. doi:10.1073/pnas.0702624104. PMC 1876592. PMID 17470779.
- Tomasello, M. 2008. Origins of Human Communication. Cambridge, Massachusetts: MIT Press.
- Weiss, B. (1974). "Medieval Muslim discussions of the origin of language". Zeitschrift der Deutschen Morgenländischen Gesellschaft. 124 (1): 33–41.
- Weiss, B. (1987). "'Ilm al-wad': an introductory account of a later Muslim philological science". Arabica. 34 (1): 339–356. doi:10.1163/157005887X00054.
- Zahavi, A. and A. Zahavi 1997. The Handicap Principle. A missing piece in Darwin's puzzle. New York and Oxford: Oxford University Press.
External links
| Look up glottogony in Wiktionary, the free dictionary. |
- Origin of Language – Givens, David B.
- Behavioral and Biological Origins of Modern Humans – Klein, Richard G.
- The Origin of Language – Vajda, Edward
- First Language Acquisition – Vajda, Edward
- Speaking in Tongues: The History of Language
- Decoding Chomsky: Science and revolutionary politics – Chris Knight
| Taxonomy (Hominins) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ancestors |
| ||||||||||||||||||||||||||
| Models |
| ||||||||||||||||||||||||||
| Timelines | |||||||||||||||||||||||||||
| Others |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||
Long-range comparative linguistics | |
|---|---|
| Concepts | |
| Language families | |
| Linguists | |
| Publications | |
| Authority control |
|
|---|