Paranthropus robustus
Paranthropus robustus is a species of australopithecine from the Early and possibly Middle Pleistocene of the Cradle of Humankind, South Africa, about 2 to 1 or 0.6 million years ago. Discovered in 1938, it was among the first early hominins described and the first discovered robust australopithecine, and became the type species for the genus Paranthropus. However, it is argued that Paranthropus is an invalid grouping and synonymous with Australopithecus, so the species is also often classified as Australopithecus robustus.
| Paranthropus robustus | |
|---|---|
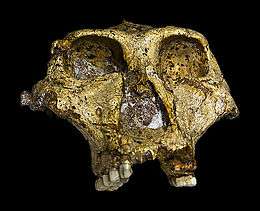 | |
| Presumed-male SK 48 at the Transvaal Museum | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Primates |
| Suborder: | Haplorhini |
| Infraorder: | Simiiformes |
| Family: | Hominidae |
| Subfamily: | Homininae |
| Tribe: | Hominini |
| Genus: | †Paranthropus |
| Species: | †P. robustus |
| Binomial name | |
| †Paranthropus robustus Broom, 1938 | |
| Synonyms | |
| |
Robust australopithecines are characterised by heavily built skulls capable of producing high stresses and bite forces, and some of the largest molars with the thickest enamel of any known ape. P. robustus may have had a genetic susceptibility for pitting enamel hypoplasia on the teeth, and seems to have had a dental cavity rate similar to non-agricultural modern humans. The species is thought to have exhibited marked sexual dimorphism, with males substantially larger and more robust than females. The brain volume of the specimen SK 1585 is estimated to have been 476 cc. Based on 3 specimens, males may have been 132 cm (4 ft 4 in) tall and females 110 cm (3 ft 7 in). Based on 4 specimens, males averaged 40 kg (88 lb) in weight and females 30 kg (66 lb). P. robustus limb anatomy is similar to that of other australopithecines, which may indicate a less efficient walking ability than modern humans, and perhaps some degree of arboreality in the trees.
In regard to plant foods, P. robustus likely had a similar diet to the contemporary Homo ergaster/Homo erectus. P. robustus is thought to have also consumed fruits, underground storage organs, and perhaps honey and termites. P. robustus may have used bones as tools to extract and process food. It is unclear if P. robustus lived in a harem society like gorillas or a multi-male society like baboons. P. robustus society may have been patrilocal, with adult females more likely to leave the group than males, but males may have been more likely to be evicted indicated by higher male mortality rates and assumed increased risk of predation to solitary individuals. P. robust contended with sabertooth cats, leopards, and hyenas on the mixed, open-to-closed landscape, and P. robustus bones probably accumulated in caves due to big cat predation.
Research history
The first remains, a partial skull including a part of the jawbone (TM 1517), were discovered in June 1938 at Kromdraai, South Africa, by local schoolboy Gert Terblanche. He gave the remains to South African conservationist Charles Sydney Barlow, who then relayed it to South African palaeontologist Robert Broom. Broom noted the Kromdraai remains were especially robust compared to other hominins. At this point in time, Australian anthropologist Raymond Dart had made the very first claim (quite controversially at the time) of an early ape-like human ancestor in 1924 from South Africa, Australopithecus africanus, based on the Taung child. In 1936, Broom had described "Plesianthropus transvaalensis" (now synonymised with A. africanus) from the Sterkfontein Caves only 2 km (1.2 mi) west from Kromdraii. Broom considered all these species, which dated to the Pleistocene and were found in the same general vicinity (now called the "Cradle of Humankind"), evidence of a greater diversity of hominins in the Pliocene out of which they and modern humans descended from, and that several hominin taxa existed alongside human ancestors. In August 1938, Broom classified the robust Kromdraai remains into a new genus as Paranthropus robustus.[1] "Paranthropus" derives from Ancient Greek παρα para beside or alongside; and άνθρωπος ánthropos man.[2] In 1939, Broom hypothesised that Paranthropus was closely related to the similarly large-toothed ape Gigantopithecus from Asia (extinct apes were primarily known from Asia at the time) believing Gigantopithecus to have been a hominin.[3]

In 1948, at the nearby Swartkrans Cave, Broom and South African palaeontologist John Talbot Robinson described "P. crassidens" based on a subadult jaw, SK 6, which was younger than the fossils from Kromdraai. They believed that the Swartkrans Paranthropus were reproductively isolated from Kromdraai Paranthropus and they eventually speciated.[4] By the 21st century, "P. crassidens" had more or less fallen out of use in favour of P. robustus.[5] In 1951, anthropologists Sherwood Washburn and Bruce D. Patterson recommended limiting hominin genera to only Australopithecus and Homo.[6] It has since been debated whether or not Paranthropus is a junior synonym of Australopithecus.[5]
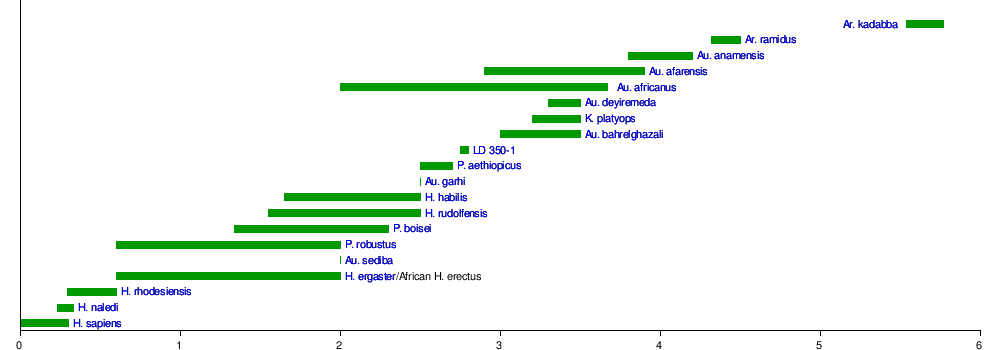
The genus Paranthropus (otherwise known as "robust australopithecines") now also includes the East African P. boisei and P. aethiopicus. It is unclear if this is a valid natural grouping (monophyletic) or an invalid grouping of similar-looking hominins (paraphyletic). Because skeletal elements are so limited in these species, their affinities with each other and to other australopithecines is difficult to gauge with accuracy. The jaws are the main argument for monophyly, but such anatomy is strongly influenced by diet and environment, and could in all likelihood have evolved independently in P. robustus and P. boisei. Proponents of monophyly consider P. aethiopicus to be ancestral to the other two species, or closely related to the ancestor. Proponents of paraphyly allocate these three species to the genus Australopithecus as A. boisei, A. aethiopicus, and A. robustus; and propose that P. robustus is closely related to or descended from A. africanus.[7]
| |||||||||||||||||||||
| |||||||||||||||||||||
| Family tree showing Paranthropus as monophyletic (top) and paraphyletic (bottom)[7]:119 |
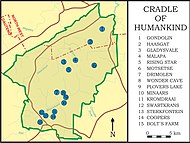
P. robustus was only definitively identified at Kromdraai and Swartkrans until around the turn of the century when the species was reported elsewhere in the Cradle of Humankind in Sterkfontein, Gondolin, Coopers, and Drimolen Caves.[5] In 2000, the most complete P. robustus skull, DNH-7, was discovered, preserving an articulated jawbone with almost all the teeth.[8] In 2020, the oldest P. robustus specimen, the braincase DNH 152, was dated to 2.04–1.95 million years ago.[9] P. robustus is recorded in Swartkrans until Member 3 dated to 1–0.6 mya (the Middle Pleistocene), though more likely the younger side of the estimate, which could mean this species outlived East African Paranthropus.[10]
 |
Anatomy
Skull

.JPG)
The P. robustus brain volume was first measured in 1972 by American physical anthropologist Ralph Holloway using the skullcap SK 1585, which is missing part of the frontal bone, and he reported a volume of about 530 cc.[11] In 2000, frontal bone anatomy was filled in using the P. boisei specimens KNM-ER 407, OH 5, and KNM-ER 732, and the brain volume of SK 1585 was recalculated at about 476 cc. Overall brain anatomy of P. robustus appears to have been more like that of non-human apes.[12] The posterior semicircular canals in the inner ear of SK 46 and SK 47 are unlike those of Australopithecus or Homo, suggesting different locomotory patterns since inner ear anatomy affects the vestibular system (sense of balance). The posterior semicircular canals of modern humans are thought to aid in stabilisation while running.[13]
Typical of Paranthropus, P. robustus exhibits post-canine megadontia with enormous cheek teeth but human-sized incisors and canines. The premolars are shaped like molars. P. robustus had a tall face with slight prognathism (the face was not completely flat). The skull had a well-defined sagittal crest on the midline and inflated cheek bones, which likely supported massive temporal muscles important in biting. The cheeks project so far from the face that, when in top-view, the nose appears to sit at the bottom of a concavity (a dished face). This displaced the eye sockets forward somewhat, causing a weak brow ridge and receding forehead. The inflated cheeks also would have pushed the masseter muscle (important in biting down) forward and pushed the tooth rows back, which would have created a higher bite force on the premolars. The ramus of the jawbone, which connects the lower jaw to the upper jaw, is tall, which would have increased lever arm (and thereby, torque) of the masseter and medial pterygoid muscles (both important in biting down), further increasing bite force.[14] However, the well-defined sagittal crest and inflated cheeks are absent in the presumed-female skull DNH-7, so male P. robustus may have been much larger and heavily built than females (P. robustus was sexually dimorphic).[8]
Size
Broom had noted that the ankle bone and humerus of the presumed male TM 1517 were about the same dimensions of that of a modern San woman, and so assumed humanlike proportions in P. robustus. The first attempt at estimating Paranthropus body size was made by Robinson in 1972. He calculated the humerus-to-femur ratio of P. robustus by using the presumed female humerus of STS 7 and comparing it with the presumed male femur of STS 14. He also had to estimate the length of the humerus using the femur assuming a similar degree of sexual dimorphism between P. robustus and humans. Comparing the ratio to humans, he concluded that P. robustus was a heavily-built species with a height of 140–150 cm (4 ft 7 in–4 ft 11 in) and a weight of 68–91 kg (150–201 lb).[15]
In 1991, American anthropologist Henry McHenry reported much lighter weights as well as notable sexual dimorphism. He estimated the living size of Swartkrans specimens by scaling down the dimensions of an average modern human to meet a preserved leg or foot element (he considered the arm measurements too variable among hominins to give accurate estimates). At Members 1 and 2, about 35% of the P. robustus individuals were the same size as those in a 28 kg (62 lb) human, 22% in a 43 kg (95 lb) human, and the remaining 43% bigger than the former but less than a 54 kg (119 lb) human except for KNM‐ER 1464 (an ankle bone). At Member 3, all individuals were consistent with a 45 kg (99 lb) human.[16] P. robustus sites are oddly dominated by small adults, which could be explained as heightened predation or mortality of the larger males of a group.[17] McHenry also estimated the living height of 3 P. robustus specimens (male SK 82, male SK 97, and female or subadult SK 3155), by scaling down an average human to meet the estimated size of the preserved femur, as 126 cm (4 ft 2 in), 137 cm (4 ft 6 in), and 110 cm (3 ft 7 in) respectively. Based on just these three, he reported an average height of 132 cm (4 ft 4 in) for P. robustus males and 110 cm (3 ft 7 in) for females.[18]
In 2001, two more femoral fragments were discovered, which would give an average of about 42 kg (93 lb) for males and 30 kg (66 lb) for females. If these four individuals are representative of the entire species, this degree of sexual dimorphism is greater than what is exhibited in humans and chimps, but less than orangutans and gorillas. Female P. robustus were about the same estimated weight as female H. erectus in Swartkrans, but male H. erectus were much bigger at 55 kg (121 lb).[19]
Torso
Few vertebrae are assigned to P. robustus. The only thoracolumbar series (thoracic and lumbar series) preserved belongs to the juvenile SKW 14002, and either represents L1–L4 or L2–L5. Much like that of A. africanus, the T12 (last thoracic vertebra) of SK 3981a is relatively elongated, and the top surface is kidney-shaped. The T12 is more compressed in height than that of other australopithecines and modern apes. The shape of the lumbar vertebrae is much more similar to that of Turkana boy (H. ergaster/H. erectus) and humans than other australopithecines. The pedicles (which jut out diagonally from the vertebra) of the lower lumbar vertebra SK 3981b are much more robust than other australopithecines and are within the range of humans, and the transverse processes indicate powerful iliolumbar ligaments. These could have bearing on the amount of time spent upright compared to other australopithecines.[20]
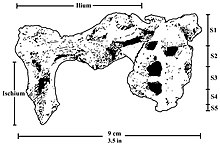
The pelvis is similar to the pelvises of A. africanus and A. afarensis, but it has a wider iliac blade and smaller acetabulum and hip joint.[21] Like modern humans, the ilium of P. robustus features development of the surface and thickening of the posterior superior iliac spine, which are important in stabilising the sacrum, and indicates lumbar lordosis (curvature of the lumbar vertebrae) and thus bipedalism. The anatomy of the sacrum and the first lumbar vertebra (at least the vertebral arch), preserved in DNH 43, are similar to those of other australopithecines.[22] The pelvis seems to indicate a more-or-less humanlike hip joint consistent with bipedalism, though differences in overall pelvic anatomy may indicate P. robustus used different muscles to generate force and perhaps had a different mechanism to direct force up the spine. This is similar to the condition seen in A. africanus. This could potentially indicate the lower limbs had a wider range of motion than modern humans.[23]
Limbs
The distal (lower) humerus of P. robustus falls within the variation of both modern humans and chimps, as the distal humerus is quite similar between humans and chimps.[24] The radius of P. robustus is the same as in Australopithecus species. The wrist joint has the same manoeuverability as that of modern humans rather than the greater flexion achieved by non-human apes, but the head of radius (the elbow) seems to have been quite capable of maintaining stability when the forearm is flexed like non-human apes. It is possible this reflects some arboreal activity in the trees as is controversially postulated in other australopithecines.[25] SKX 3602 exhibits a robust radial styloid processes near the hand which indicates strong brachioradialis muscles and extensor retinaculae. Like humans, the finger bones are uncurved and have weaker muscle attachment than non-human apes, though the proximal phalanges are smaller than in humans. The intermediate phalanges are stout and straight like humans, but have stouter bases and better developed flexor impressions. The distal phalanges seem to be essentially humanlike.[26] The hand is consistent with a humanlike precision grip which would have made possible the production or usage of tools requiring greater motor functions than non-human primate tools.[27]
The femur, like P. boisei and H. habilis, is flattened anteroposteriorly (on the front and back side). This may indicate a walking gait more similar to early hominins than to modern humans (less efficient gait).[21] Four femora assigned to P. robustus—SK 19, SK 82, SK 97, and SK 3121—exhibit an apparently high anisotropic trabecular bone (at the hip joint) structure, which could indicate reduced mobility of the hip joint compared to non-human apes, and the ability to produce forces consistent with humanlike bipedalism.[28] The femoral head StW 311, which either belongs to P. robustus or early Homo, seems to have habitually been placed in highly flexed positions, which would be consistent with frequent climbing activity. It is unclear if frequent squatting could be a valid alternative interpretation.[29] The big toe bone of P. robustus is not dextrous, which indicates a humanlike foot posture and range of motion, but the more distal ankle joint would have inhibited the modern human toe-off gait cycle. P. robustus and H. habilis may have achieved about the same grade of bipedality.[30]
Palaeobiology
Diet and technology
In 1954, Robinson suggested that the heavily built skull of P. robustus was indicative of a specialist diet specifically adapted for processing a narrow band of foods. Because of this, the predominant model of Paranthropus extinction for the latter half of the 20th century was that they were unable to adapt to the volatile climate of the Pleistocene, unlike the much more adaptable Homo.[31] However, the microwear patterns in P. robustus suggest hard food was infrequently consumed, and the heavy build of the skull was only relevant when eating less desirable fallback foods.[31] Such a strategy is similar to that used by modern gorillas, which can sustain themselves entirely on lower quality fallback foods year-round, as opposed to lighter built chimps (and presumably gracile australopithecines) which require steady access to high quality foods.[32]
P. robustus is now thought to have been an omnivore, with a diet similar to early Homo.[31] Using Carbon isotope analysis, P. robustus likely subsisted on mainly C4 savanna plants or C3 forest plants depending on the season, which could indicate either seasonal shifts in diet or seasonal migration from forest to savanna.[33] H. ergaster appears to have consumed about the same proportion of C3 to C4 based foods as P. robustus.[34] P. robustus likely also consumed seeds.[33][35] A high cavity rate could indicate honey consumption.[36] Juvenile P. robustus may have relied more on tubers than adults, given the elevated levels of Strontium compared to adults in teeth from Swartkrans Cave, which, in the area, was most likely sourced from tubers. Dentin exposure on juvenile teeth could indicate early weaning, or a more abrasive diet than adults which wore away the cementum and enamel coatings, or both. It is also possible juveniles were less capable of removing grit from dug-up food rather than purposefully seeking out more abrasive foods.[37]
Bone tools dating between 2.3 and 0.6 mya—found in abundance in Swartkrans,[38] Kromdraai, and Drimolen Caves—are often associated with P. robustus, and may have been used to cut or process vegetation,[27] process fruits (namely marula fruit), strip tree bark,[39] or dig up tubers or termites.[38][39][40] Though Homo is also known from these caves, their remains are comparatively scarce to Paranthropus, making Homo-attribution less likely.[40] The form of P. robustus incisors appears to be intermediate between H. erectus and modern humans, which could indicate less food processing done by the teeth due to preparation with simple tools.[37] The tools also cooccur with two Oldawan or possibly Acheulian stone tool industries typically attributed to Homo, but two stone tools from Kromdraai B could possibly be attributed to P. robustus as Homo has not been confidently identified in this layer yet. However, it is unclear if the stone tools were reworked or not. The bone tools were typically sourced from the shaft of long bones from medium- to large-sized mammals, but tools sourced from mandibles, ribs, and horn cores have also been found.[40] The bone tools were not manufactured or purposefully shaped for a task. However, since the bones display no weathering, and there is a preference displayed for certain bones, raw materials were likely specifically hand picked.[40]
Burnt bones were also associated with the inhabitants of Swartkrans, which could indicate some of the earliest fire usage.[41] Out of about 60,000 bones, 270 were burned. However, these bones were found in Member 3, where Paranthropus remains are rarer than H. erectus. Some specimens display cutting or percussion, and burning appears to be a recurring event, which suggest attribution to hominin activity, but it is nonetheless also plausible that the bones were simply burned in a natural wildfire.[42][43]
Social structure
Given the marked anatomical and physical differences with modern great apes, there may be no modern analogue for australopithecine societies, so comparisons drawn with modern primates are highly speculative.[44][45] P. robustus appears to have had pronounced sexual dimorphism, with males notably larger than females. This is commonly correlated with a male-dominated polygamous society, such as the harem society of modern forest-dwelling silverback gorillas where one male has exclusive breeding rights to a group of females. Estimated male-female size disparity in P. robustus is comparable to gorillas (based on facial dimensions), and younger males were less robust than older males (delayed maturity is also exhibited in gorillas).[46]
However, among primates, delayed maturity is also exhibited in the rhesus monkey which has a multi-male society, and may not be an accurate indicator of social structure. If P. robustus preferred a savanna habitat, a multi-male society would have been more productive to better defend the troop from predators in the more exposed environment, much like savanna baboons.[45] A 2011 Strontium isotope study of P. robustus teeth from the dolomite Sterkfontein Valley found that, like other hominins, but unlike other great apes, P. robustus females were more likely to leave their place of birth (patrilocal). This also discounts the plausibility of a harem society, which would have resulted in a matrilocal society due to heightened male–male competition. Males did not seem to have ventured very far from the valley, which could either indicate small home ranges, or that they preferred dolomitic landscapes due to perhaps cave abundance or factors related to vegetation growth.[44] Nonetheless, because male P. robustus appear to have had a higher mortality rate than females, it is possible that males were more likely to be kicked out of a group, and these lone males had a higher risk of predation.[45] A female moving out of her birth group may have spent little time alone and transferred immediately to another established group.[46]
Development
Australopithecines are generally considered to have had a faster, apelike growth rate than modern humans. While growing, the front part of the jaw in P. robustus is depository whereas the sides are resorptive. For comparison, chimp jaws are generally depository reflecting prognathism, and modern humans resorptive reflecting a flat face. In Paranthropus, this may have functioned to thicken the palate. Unlike apes and gracile australopithecines, but like humans, the premaxillary suture between the premaxilla and the maxilla (on the palate) formed early in development. At early stages, the P. robustus jawbone is somewhat similar to that of modern humans, but the breadth grows in P. robustus, as to be expected from its incredible robustness in adulthood. By the time the first permanent molar erupts, the body of the mandible and the front jaw broaden, and the ramus of the mandible elongates, diverging from the modern human trajectory. Because the ramus is so tall, it is suggested that P. robustus experienced more anterior face rotation than modern humans and apes. Growth is most marked between the eruptions of the first and second permanent molars, most notably in terms of the distance from the back of the mouth to the front of the mouth, probably to make room for the massive postcanine teeth. Like humans, jaw robustness decreases with age, though it decreases slower in P. robustus.[47]

Dental development seems to have followed about the same timeframe as it does in modern humans and most other hominins, but, since P. robustus molars are markedly larger, rate of tooth eruption would have been accelerated.[48] The early age of the eruption and the fast rate of tooth root extension for the permanent first molar of the 3–4 year old specimen SK 63 is consistent with that of non-human apes.[49]
Females may have reached skeletal maturity by the time the third molar erupted, but males appear to have continued growing after reaching dental maturity, during which time they become markedly more robust than females (sexual bimaturism). Similarly, male gorillas complete dental development about the same time as females, but continue growing for up to 5 or 6 years; and male mandrills complete dental development before females, but continue growing for several years more.[46] It is debated whether or not P. robustus had a defined growth spurt in terms of overall height during adolescence, an event unique to humans among modern apes.[47]
Pathology
P. robustus seems to have had notably high rates of pitting enamel hypoplasia (PEH), where tooth enamel formation is spotty instead of mostly uniform. In P. robustus, about 47% of baby teeth and 14% of adult teeth were affected, in comparison to about 6.7% and 4.3% respectively in any other tested hominin species. The condition of these holes covering the entire tooth is consistent with the modern human ailment amelogenesis imperfecta. However, since circular holes in enamel coverage are uniform in size, only present on the molar teeth, and have the same severity across individuals, the PEH may have been a genetic condition. It is possible that the coding region concerned with thickening enamel also increased the risk of developing PEH.[50]
As many as four P. robustus individuals have been identified as having had dental cavities, indicating a rate similar to non-agricultural modern humans (1–5%). This is odd as P. robustus is thought to have had a diet high in gritty foods, and gritty foods should decrease cavity incidence rate, so P. robustus may have often consumed high-sugar cavity-causing foods. PEH may have also increased susceptibility to cavities.[51] A molar from Drimolen showed a cavity on the tooth root, a rare occurrence in fossil great apes. In order for cavity-creating bacteria to reach this area, the individual would have had to have also presented either alveolar resportion, which is commonly associated with gum disease; or super-eruption of teeth which occurs when teeth become worn down and have to erupt a bit more in order to maintain a proper bite, exposing the root in the process. The latter is most likely, and the exposed root seems to have caused hypercementosis to anchor the tooth in place. The cavity seems to have been healing, possibly due to a change in diet or mouth microbiome, or the loss of the adjacent molar.[36]
In a sample of 15 P. robustus specimens, all of them exhibited mild to moderate alveolar bone loss resulting from periodontal disease (the wearing away of the bone which supports the teeth due to gum disease). In contrast, in a sample of 10 A. africanus specimens, three exhibited no pathologies of the alveolar bone. Measuring the distance between the alveolar bone and the cementoenamel junction, P. robustus possibly suffered from a higher rate of tooth-attachment loss, unless P. robustus had a higher cervical height (the slightly narrowed area where the crown meets the root) in which case these two species had the same rate of tooth-attachment loss. If the former is correct, then the difference may be due to different dietary habits, chewing strategies, more pathogenic mouth microflora in P. robustus, or some immunological difference which made P. robustus somewhat more susceptible to gum disease.[52]
Palaeoecology

The Pleistocene Cradle of Humankind was mainly dominated by the springbok Antidorcas recki, but other antelope, giraffes, and elephants were also seemingly abundant megafauna. Other known primates are early Homo, the Hamadryas baboon, and the extinct colobine monkey Cercopithecoides williamsi. The carnivore assemblage comprises the sabertoothed cats Dinofelis and Megantereon, and the hyena Lycyaenops silberbergi. These animals indicate a mixed, open-to-closed landscape featuring perhaps montane grasslands and shrublands[53] Australopithecines and early Homo likely preferred cooler conditions than later Homo, as there are no australopithecine sites that were below 1,000 m (3,300 ft) in elevation at the time of deposition. This would mean that, like chimps, they often inhabited areas with an average diurnal temperature of 25 °C (77 °F), dropping to 10 or 5 °C (50 or 41 °F) at night.[54]
Australopithecine bones may have accumulated in caves due to large carnivore activity, dragging in carcasses. The juvenile P. robustus skullcap SK 54 has two puncture marks consistent with the lower canines of the leopard specimen SK 349 from the same deposits. In 1983, South African palaeontologist Charles Kimberlin Brain hypothesised that Dinofelis and perhaps also hunting hyenas specialised on killing australopithecines.[55] However, carbon isotope analysis indicates these species predominantly ate large grazers, and the leopard, the sabertoothed Megantereon, and the spotted hyena were more likely to have regularly consumed P. robustus.[56] Brain was unsure if these predators actively sought them out and brought them back to the cave den to eat, or inhabited deeper recesses of caves and ambushed them when they entered. Baboons in this region modern day often shelter in sinkholes especially on cold winter nights, though Brain proposed that australopithecines seasonally migrated out of the Highveld and into the warmer Bushveld, only taking up cave shelters in spring and autumn.[55]
P. robustus also cohabited the Cradle of Humankind with H. ergaster/H. erectus.[19][42][9] In addition, these two species resided alongside A. sediba from 2.04–1.95 million years ago. Around 2.07 million years ago, just before the arrival of P. robustus and H. erectus, A. africanus went extinct from the area. It is possible that South Africa was a refugium for Australopithecus until about 2 million years ago with the beginning of major climatic variability and volatility, and potentially competition with Homo and Paranthropus.[9]
See also
- African archaeology
- Australopithecus africanus – Extinct hominid from South Africa
- Australopithecus sediba – 2 million year old hominin from the Cradle of Humankind
- Homo ergaster – Extinct species or subspecies of archaic human
- Homo habilis – Archaic human species from 2.1 to 1.5 mya
- Homo naledi – Small-brained South African archaic human
- Homo rudolfensis – Extinct hominin from the Early Pleistocene of East Africa
- Paranthropus aethiopicus – Extinct species of hominin of East Africa
- Paranthropus boisei – Extinct species of hominin of East Africa
References
- Broom, R. (1938). "The Pleistocene Anthropoid Apes of South Africa". Nature. 142: 377–339. doi:10.1038/142377a0.
- "Paranthropus". Merriam–Webster Dictionary. Retrieved 20 June 2020.
- Broom, R. (1939). "The dentition of the Transvaal Pleistocene anthropoids, Plesianthropus and Paranthropus" (PDF). Annals of the Transvaal Museum. 19 (3): 303–314.
- Broom, R. (1948). "Another new type of fossil ape-man". Nature. 162 (4132): 57. doi:10.1038/163057a0. PMID 18106151.
- Constantino, P. J.; Wood, B. A. (2004). "Paranthropus Paleobiology". Miscelanea en Homenaje a Emiliano Aguirre. Paleoantropologia. III. Museo Arqueológico Regional.
- Washburn, S. L.; Patterson, B. (1951). "Evolutionary Importance of the South African 'Man-apes'". Nature. 167: 650–651. doi:10.1038/167650a0.
- Wood, Bernard; Constantino, Paul (2007). "Paranthropus boisei: Fifty years of evidence and analysis". American Journal of Physical Anthropology. 134 (Suppl 45): 117–121. doi:10.1002/ajpa.20732. PMID 18046746.
- Keyser, A. W. (2000). "The Drimolen skull: the mostcomplete australopithecinecranium and mandible to date". South African Journal of Science. 96: 189–197.
- Herries, A. I. R.; Martin, J. M.; et al. (2020). "Contemporaneity of Australopithecus, Paranthropus, and early Homo erectus in South Africa". Science. 368 (6486): eaaw7293. doi:10.1126/science.aaw7293.
- Herries, A. I. R.; Curnoe, D.; Adams, J. W. (2009). "A multi-disciplinary seriation of early Homo and Paranthropus bearing palaeocaves in southern Africa". Quaternary International. 202 (1–2): 14–28. Bibcode:2009QuInt.202...14H. doi:10.1016/j.quaint.2008.05.017.
- Holloway, R. L. (1972). "New australopithecine endocast, SK 1585, from Swartkrans, South Africa". American Journal of Physical Anthropology. 37 (2): 173–185. doi:10.1002/ajpa.1330370203.
- Falk, D.; Redmond, Jr., J. C.; Guyer, J.; et al. (2000). "Early hominid brain evolution: a new look at old endocasts" (PDF). Journal of Human Evolution. 38 (5): 695–717. doi:10.1006/jhev.1999.0378.
- Beaudet, A.; Clarke, R. J.; Bruxelles, L.; et al. (2019). "The bony labyrinth of StW 573 ("Little Foot"): Implications for early hominin evolution and paleobiology". Journal of Human Evolution. 127: 67–80. doi:10.1016/j.jhevol.2018.12.002.
- Cartmill, M.; Smith, F.H. (2009). The Human Lineage. John Wiley and Sons. pp. 152–157. ISBN 978-0-471-21491-5.
- Wolpoff, M. H. (1974). "Reviewed Work: Early Hominid Posture and Locomotion by John T. Robinson". Human Biology. 46 (4): 719–724. JSTOR 41462378.
- McHenry, H. M. (1991). "Petite bodies of the "robust" australopithecines". American Journal of Physical Anthropology. 86 (4): 445–454. doi:10.1002/ajpa.1330860402.
- Braga, J.; Thackeray, J. F.; Bruxelles, L.; Dumoncel, J.; Fourvel, J.-P. (2017). "Stretching the time span of hominin evolution at Kromdraai (Gauteng, South Africa): Recent discoveries". Comptes Rendus Palevol. 16 (1): 58–70. doi:10.1016/j.crpv.2016.03.003.
- McHenry, H. M. (1991). "Femoral lengths and stature in Plio-Pleistocene hominids". American Journal of Physical Anthropology. 85 (2): 149–158. doi:10.1002/ajpa.1330850204. PMID 1882979.
- Susman, R. L.; de Ruiter, D.; Brain, C. K. (2001). "Recently identified postcranial remains of Paranthropus and Early Homo from Swartkrans Cave, South Africa". Journal of Human Evolution. 41 (6): 607–629. doi:10.1006/jhev.2001.0510. PMID 11782111.
- Sanders, W. J. (1998). "Comparative morphometric study of the australopithecine vertebral series Stw-H8/H41" (PDF). Journal of Human Evolution. 34 (3): 249–302. doi:10.1006/jhev.1997.0193. PMID 9547457.
- Wood B, Richmond BG (2000). "Human evolution: taxonomy and paleobiology". Journal of Anatomy. 197 (1): 35–36. doi:10.1046/j.1469-7580.2000.19710019.x. PMC 1468107. PMID 10999270.
- Gommery, D.; Senut, B.; Keyser, A. (2002). "A fragmentary pelvis of Paranthropus robustus of the Plio-Pleistocene site of Drimolen Republic of South Africa". Geobios (in French). 35 (2): 265–281. doi:10.1016/S0016-6995(02)00022-0.
- Macchiarelli R, Bondioli L, Galichon V, Tobias PV (February 1999). "Hip bone trabecular architecture shows uniquely distinctive locomotor behaviour in South African australopithecines". Journal of Human Evolution. 36 (2): 211–32. doi:10.1006/jhev.1998.0267. PMID 10068067.
- Straus, Jr., W. L. (1948). "The humerus of Paranthropus robustus". American Journal of Physical Anthropology. 6 (3): 285–313. doi:10.1002/ajpa.1330060305.
- Grine, F. E.; Susman, R. L. (1991). "Radius of Paranthropus robustus from member 1, Swartkrans Formation, South Africa". American Journal of Physical Anthropology. 84 (3): 229–248. doi:10.1002/ajpa.1330840302.
- Susman, R. L. (1989). "New hominid fossils from the Swartkrans formation (1979–1986 excavations): Postcranial specimens". American Journal of Physical Anthropology. 79 (4): 451–474. doi:10.1002/ajpa.1330790403.
- Susman, R. L. (1988). "Hand of Paranthropus robustus From Member 1, Swartkrans: Fossil Evidence for Tool Behavior". Science. 240 (4853): 781–784. doi:10.1126/science.3129783. JSTOR 1701549.
- Ryan, T. M.; Carlson, K. J.; Gordon, A. D.; et al. (2018). "Human-like hip joint loading in Australopithecus africanus and Paranthropus robustus". Journal of Human Anthropology. 121: 12–24. doi:10.1016/j.jhevol.2018.03.008. PMID 29706230.
- Georgiou, L.; Dunmore, C. J.; Bardo, A. (2020). "Evidence for habitual climbing in a Pleistocene hominin in South Africa". Proceedings of the National Academy of Sciences. 117 (15): 8416–8423. doi:10.1073/pnas.1914481117.
- Susman, R. L.; Brain, T. M. (1988). "New first metatarsal (SKX 5017) from Swartkrans and the gait of Paranthropus robustus". American Journal of Physical Anthropology. 77 (1): 7–15. doi:10.1002/ajpa.1330770103. PMID 3189526.
- Wood, B.; Strait, D. (2004). "Patterns of resource use in early Homo and Paranthropus". Journal of Human Evolution. 46 (2): 119–162. doi:10.1016/j.jhevol.2003.11.004. PMID 14871560.
- Laden, G.; Wrangham, R. (2005). "The rise of the hominids as an adaptive shift in fallback foods: Plant underground storage organs (USOs) and australopith origins". Journal of Human Evolution. 49 (4): 482–498. doi:10.1016/j.jhevol.2005.05.007.
- Sponheimer, M.; Passey, B. H.; de Ruiter, D. J.; et al. (2006). "Isotopic Evidence for Dietary Variability in the Early Hominin Paranthropus robustus". Science. 314 (5801): 980–982. Bibcode:2006Sci...314..980S. doi:10.1126/science.1133827. PMID 17095699.
- Lee-Thorp, J.; Thackeray, J. F.; der Merwe, N. V. (2000). "The hunters and the hunted revisited". Journal of Human Evolution. 39 (6): 565–576. doi:10.1006/jhev.2000.0436. PMID 11102267.
- Constantino, P. J.; Borrero-Lopez, O.; Lawn, B. R. (2018). "Mechanisms of Tooth Damage in Paranthropus Dietary Reconstruction". Biosurface and Biotribology. 4 (3): 73–78. doi:10.1049/bsbt.2018.0017.
- Towle, I.; Riga, A.; Irish, J. D.; et al. (2019). "Root caries on a Paranthropus robustus third molar from Drimolen" (PDF). American Journal of Physical Anthropology. 170 (2): 319–323. doi:10.1002/ajpa.23891. PMID 31265762.
- Williams, F. L. (2015). "Dietary proclivities of Paranthropus robustus from Swartkrans, South Africa". Anthropological Review. 78 (1): 1–19. doi:10.1515/anre-2015-0001.
- Backwell, L. R.; d'Errico, F. (2001). "Evidence of termite foraging by Swartkrans early hominids". Proceedings of the National Academy of Sciences. 98 (4): 1358–1363. doi:10.1073/pnas.021551598. PMC 29261. PMID 11171955.
- d'Errico, F.; Backwell, L. (2009). "Assessing the function of early hominin bone tools". Journal of Archaeological Science. 36 (8): 1764–1773. doi:10.1016/j.jas.2009.04.005.
- Stammers, R. C.; Caruana, M.; Herries, A. I. R. (2018). "The first bone tools from Kromdraai and stone tools from Drimolen, and the place of bone tools in the South African Earlier Stone Age". Quaternary International. 495: 87–101. Bibcode:2018QuInt.495...87S. doi:10.1016/j.quaint.2018.04.026.
- Brain, C. K.; Sillent, A. (1988). "Evidence from the Swartkrans cave for the earliest use of fire". Nature. 336 (6198): 464–466. Bibcode:1988Natur.336..464B. doi:10.1038/336464a0.
- Pickering, T. R. (2012). "What's new is old: comments on (more) archaeological evidence of one-million-year-old fire from South Africa". South African Journal of Science. 108 (5–6): 1–2. doi:10.4102/sajs.v108i5/6.1250.
- Gowlett, J. A. J.; Wrangham, R. W. (2013). "Earliest fire in Africa: towards the convergence of archaeological evidence and the cooking hypothesis". Azania: Archaeological Research in Africa. 48 (1): 16–17. doi:10.1080/0067270X.2012.756754.
- Copeland, S. R.; Sponheimmer, M.; de Ruiter, D. J.; Lee-Thorp, J. (2011). "Strontium isotope evidence for landscape use by early hominins". Nature. 474 (7349): 76–78. doi:10.1038/nature10149. PMID 21637256.
- Kaszycka, K. A. (2016). "Australopithecus robustus societies - one-male or multimale?". South African Journal of Science. 112 (1–2): 124–131. doi:10.17159/sajs.2016/20150165.
- Lockwood, C. A.; Menter, C. G.; Moggi-Cecchi, J.; Keyser, A. W. (2007). "Extended male growth in a fossil hominin species". Science. 318 (5855): 1443–1446. Bibcode:2007Sci...318.1443L. doi:10.1126/science.1149211. PMID 18048687.
- Cofran, Z. (2014). "Mandibular development in Australopithecus robustus". American Journal of Physical Anthropology. 154 (3): 436–446. doi:10.1002/ajpa.22527. PMID 24820665.
- Dean, M. C. (1985). "The eruption pattern of the permanent incisors and first permanent molars in Australopithecus (Paranthropus) robustus". American Journal of Physical Anthropology. 67 (3): 251–257. doi:10.1002/ajpa.1330670310. PMID 3933358.
- Dean, M. C. (1993). "Histological reconstruction of dental development and age at death of a juvenile Paranthropus robustus specimen, SK 63, from Swartkrans, South Africa". American Journal of Physical Anthropology. 91 (4): 401–419. doi:10.1002/ajpa.1330910402.
- Towle, I.; Irish, J. D. (2019). "A probable genetic origin for pitting enamel hypoplasia on the molars of Paranthropus robustus" (PDF). Journal of Human Evolution. 129: 54–61. doi:10.1016/j.jhevol.2019.01.002. PMID 30904040.
- Towle, I.; Irish, J. D.; et al. (2019). "Dental caries in human evolution: frequency of carious lesions in South African fossil hominins". BioRxiv. doi:10.1101/597385.
- Ripamonti, U. (1989). "The Hard Evidence of Alveolar Bone Loss in Early Hominids of Southern Africa". Journal of Periodontology. 60 (2): 118–120. doi:10.1902/jop.1989.60.2.118.
- Adams, J. W.; Rovinsky, D. S.; Herries, A. I. R.; Menter, C. G. (2016). "Macromammalian faunas, biochronology and palaeoecology of the early Pleistocene Main Quarry hominin-bearing deposits of the Drimolen Palaeocave System, South Africa". PeerJ. 4: e1941. doi:10.7717/peerj.1941. PMC 4841245. PMID 27114884.
- Dávid-Barrett, T.; Dunbar, R. I. M. (2016). "Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling". Journal of Human Evolution. 94. doi:10.1016/j.jhevol.2016.02.006. PMC 4874949. PMID 27178459.
- Brain, C. K. (1983). "Who Were the Hunters and Who the Hunted". The Hunters Or the Hunted?: An Introduction to African Cave Taphonomy. University of Chicago Press. ISBN 978-0-226-07090-2.
- Lee-Thorp, J.; Thackeray, J. F.; van der Merwe, N. (2010). "The hunters and the hunted revisited". Journal of Human Evolution. 39 (6): 565–576. doi:10.1006/jhev.2000.0436.
External links
| Wikimedia Commons has media related to Paranthropus robustus. |
| Wikispecies has information related to Paranthropus robustus |
- Meet Australopithecus robustus — John D. Hawk's website
- Paranthropus robustus - The Smithsonian's Human Origins Program
- Human Timeline (Interactive) – Smithsonian
| Taxonomy (Hominins) |
| ||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ancestors |
| ||||||||||||||||||||||||||
| Models |
| ||||||||||||||||||||||||||
| Timelines | |||||||||||||||||||||||||||
| Others |
| ||||||||||||||||||||||||||
| |||||||||||||||||||||||||||