Luteinizing hormone/choriogonadotropin receptor
The luteinizing hormone/choriogonadotropin receptor (LHCGR), also lutropin/choriogonadotropin receptor (LCGR) or luteinizing hormone receptor (LHR) is a transmembrane receptor found predominantly in the ovary and testis, but also many extragonadal organs such as the uterus and breasts. The receptor interacts with both luteinizing hormone (LH) and chorionic gonadotropins (such as hCG in humans) and represents a G protein-coupled receptor (GPCR). Its activation is necessary for the hormonal functioning during reproduction.




LHCGR gene
The gene for the LHCGR is found on chromosome 2 p21 in humans, close to the FSH receptor gene. It consists of 70 kbp (versus 54 kpb for the FSHR).[5] The gene is similar to the gene for the FSH receptor and the TSH receptor.
Receptor structure
The LHCGR consists of 674 amino acids and has a molecular mass of about 85–95 kDA based on the extent of glycosylation.[6]
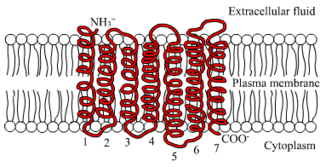
Like other GPCRs, the LHCG receptor possess seven membrane-spanning domains or transmembrane helices.[7] The extracellular domain of the receptor is heavily glycosylated. These transmembrane domains contain two highly conserved cysteine residues, which build disulfide bonds to stabilize the receptor structure. The transmembrane part is highly homologous with other members of the rhodopsin family of GPCRs.[8] The C-terminal domain is intracellular and brief, rich in serine and threonine residues for possible phosphorylation.
Ligand binding and signal transduction
Upon binding of LH to the external part of the membrane spanning receptor, a transduction of the signal takes place that activates the G protein that is bound to the receptor internally. With LH attached, the receptor shifts conformation and thus mechanically activates the G protein, which detaches from the receptor and activates the cAMP system.[9]
It is believed that a receptor molecule exists in a conformational equilibrium between active and inactive states. The binding of LH (or CG) to the receptor shifts the equilibrium between active and inactive receptors. LH and LH-agonists shift the equilibrium in favor of active states; LH antagonists shift the equilibrium in favor of inactive states. For a cell to respond to LH only a small percentage (≈1%) of receptor sites need to be activated.
Phosphorylation by cAMP-dependent protein kinases
Cyclic AMP-dependent protein kinases (protein kinase A) are activated by the signal chain coming from the G protein (that was activated by the LHCG-receptor) via adenylate cyclase and cyclic AMP (cAMP). These protein kinases are present as tetramers with two regulatory units and two catalytic units. Upon binding of cAMP to the regulatory units, the catalytic units are released and initiate the phosphorylation of proteins leading to the physiologic action. The cyclic AMP-regulatory dimers are degraded by phosphodiesterase and release 5’AMP. DNA in the cell nucleus binds to phosphorylated proteins through the cyclic AMP response element (CRE), which results in the activation of genes.[5]
The signal is amplified by the involvement of cAMP and the resulting phosphorylation. The process is modified by prostaglandins. Other cellular regulators that participate are the intracellular calcium concentration modified by phospholipase, nitric acid, and other growth factors.
In a feedback mechanism, these activated kinases phosphorylate the receptor. The longer the receptor remains active the more kinases are activated and the more receptors are phosphorylated.
Other pathways of signaling exist for the LHCGR.[6]
Action

It specifically acts to up-regulate the enzyme cholesterol side chain cleaving enzyme, which leads to the greater conversion of cholesterol into androgen precursors required to make many steroid hormones, including testosterone and estrogens.[10]
Ovary
In the ovary, the LHCG receptor is necessary for follicular maturation and ovulation, as well as luteal function. Its expression requires appropriate hormonal stimulation by FSH and estradiol. The LHCGR is present on granulosa cells, theca cells, luteal cells, and interstitial cells[6] The LCGR is restimulated by increasing levels of chorionic gonadotropins in case a pregnancy is developing. In turn, luteal function is prolonged and the endocrine milieu is supportive of the nascent pregnancy.
Testis
In the male the LHCGR has been identified on the Leydig cells that are critical for testosterone production, and support spermatogenesis.
Normal LHCGR functioning is critical for male fetal development, as the fetal Leydig cells produce testosterone to induce masculinization.
Extragonadal
LHCGR have been found in many types of extragonadal tissues, and the physiologic role of some has remained largely unexplored. Thus receptors have been found in the uterus, sperm, seminal vesicles, prostate, skin, breast, adrenals, thyroid, neural retina, neuroendocrine cells, and (rat) brain.[6]
Receptor regulation
Upregulation
Upregulation refers to the increase in the number of receptor sites on the membrane. Estrogen and FSH upregulate LHCGR sites in preparation for ovulation. After ovulation, the luteinized ovary maintains LHCGR s that allow activation in case there is an implantation.
Desensitization
The LHCGRs become desensitized when exposed to LH for some time. A key reaction of this downregulation is the phosphorylation of the intracellular (or cytoplasmic) receptor domain by protein kinases. This process uncouples Gs protein from the LHCGR. Another way to desensitize is to uncouple the regulatory and catalytic units of the cAMP system.
Downregulation
Downregulation refers to the decrease in the number of receptor sites. This can be accomplished by metabolizing bound LHCGR sites. The bound LCGR complex is brought by lateral migration to a coated pit, where such units are concentrated and then stabilized by a framework of clathrins. A pinched-off coated pit is internalized and degraded by lysosomes. Proteins may be metabolized or the receptor can be recycled. Use of long-acting agonists will downregulate the receptor population.
Modulators
Antibodies to LHCGR can interfere with LHCGR activity.
LHCGR abnormalities
Loss-of-function mutations in females can lead to infertility. In 46, XY individuals severe inactivation can cause male pseudohermaphroditism, as fetal Leydig cells during may not respond and thus interfere with masculinization.[11] Less severe inactivation can result in hypospadias or a micropenis.[6]
History
Alfred G. Gilman and Martin Rodbell received the 1994 Nobel Prize in Medicine and Physiology for the discovery of the G Protein System.
I interactions
Luteinizing hormone/choriogonadotropin receptor has been shown to interact with GIPC1.[12]
References
- GRCh38: Ensembl release 89: ENSG00000138039 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000024107 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Simoni M, Gromoll J, Nieschlag E (Dec 1997). "The follicle-stimulating hormone receptor: biochemistry, molecular biology, physiology, and pathophysiology". Endocrine Reviews. 18 (6): 739–73. doi:10.1210/er.18.6.739. PMID 9408742.
- Ascoli M, Fanelli F, Segaloff DL (Apr 2002). "The lutropin/choriogonadotropin receptor, a 2002 perspective". Endocrine Reviews. 23 (2): 141–74. doi:10.1210/er.23.2.141. PMID 11943741.
- Dufau ML (1998). "The luteinizing hormone receptor". Annual Review of Physiology. 60: 461–96. doi:10.1146/annurev.physiol.60.1.461. PMID 9558473.
- Jiang X, Dias JA, He X (Jan 2014). "Structural biology of glycoprotein hormones and their receptors: insights to signaling". Molecular and Cellular Endocrinology. 382 (1): 424–51. doi:10.1016/j.mce.2013.08.021. PMID 24001578.
- Ryu KS, Gilchrist RL, Koo YB, Ji I, Ji TH (Apr 1998). "Gene, interaction, signal generation, signal divergence and signal transduction of the LH/CG receptor". International Journal of Gynaecology and Obstetrics. 60 Suppl 1: S9-20. doi:10.1016/S0020-7292(98)80001-5. PMID 9833610.
- Dufau ML, Cigorraga S, Baukal AJ, Sorrell S, Bator JM, Neubauer JF, Catt KJ (Dec 1979). "Androgen biosynthesis in Leydig cells after testicular desensitization by luteinizing hormone-releasing hormone and human chorionic gonadotropin". Endocrinology. 105 (6): 1314–21. doi:10.1210/endo-105-6-1314. PMID 227658.
- Wu SM, Chan WY (1999). "Male pseudohermaphroditism due to inactivating luteinizing hormone receptor mutations". Archives of Medical Research. 30 (6): 495–500. doi:10.1016/S0188-4409(99)00074-0. PMID 10714363.
- Hirakawa T, Galet C, Kishi M, Ascoli M (Dec 2003). "GIPC binds to the human lutropin receptor (hLHR) through an unusual PDZ domain binding motif, and it regulates the sorting of the internalized human choriogonadotropin and the density of cell surface hLHR". The Journal of Biological Chemistry. 278 (49): 49348–57. doi:10.1074/jbc.M306557200. PMID 14507927.
Further reading
- Ji TH, Ryu KS, Gilchrist R, Ji I (1997). "Interaction, signal generation, signal divergence, and signal transduction of LH/CG and the receptor". Recent Progress in Hormone Research. 52: 431–53, discussion 454. PMID 9238862.
- Dufau ML (1998). "The luteinizing hormone receptor". Annual Review of Physiology. 60: 461–96. doi:10.1146/annurev.physiol.60.1.461. PMID 9558473.
- Ascoli M, Fanelli F, Segaloff DL (Apr 2002). "The lutropin/choriogonadotropin receptor, a 2002 perspective". Endocrine Reviews. 23 (2): 141–74. doi:10.1210/er.23.2.141. PMID 11943741.
- Amsterdam A, Hanoch T, Dantes A, Tajima K, Strauss JF, Seger R (Feb 2002). "Mechanisms of gonadotropin desensitization". Molecular and Cellular Endocrinology. 187 (1–2): 69–74. doi:10.1016/S0303-7207(01)00701-8. PMID 11988313.
- Fanelli F, Puett D (Aug 2002). "Structural aspects of luteinizing hormone receptor: information from molecular modeling and mutagenesis". Endocrine. 18 (3): 285–93. doi:10.1385/ENDO:18:3:285. PMID 12450321.
- Latronico AC, Segaloff DL (Jan 2007). "Insights learned from L457(3.43)R, an activating mutant of the human lutropin receptor". Molecular and Cellular Endocrinology. 260-262: 287–93. doi:10.1016/j.mce.2005.11.053. PMC 1785107. PMID 17055147.
- Nagayama Y, Russo D, Wadsworth HL, Chazenbalk GD, Rapoport B (Aug 1991). "Eleven amino acids (Lys-201 to Lys-211) and 9 amino acids (Gly-222 to Leu-230) in the human thyrotropin receptor are involved in ligand binding". The Journal of Biological Chemistry. 266 (23): 14926–30. PMID 1651314.
- Jia XC, Oikawa M, Bo M, Tanaka T, Ny T, Boime I, Hsueh AJ (Jun 1991). "Expression of human luteinizing hormone (LH) receptor: interaction with LH and chorionic gonadotropin from human but not equine, rat, and ovine species". Molecular Endocrinology. 5 (6): 759–68. doi:10.1210/mend-5-6-759. PMID 1922095.
- Minegishi T, Nakamura K, Takakura Y, Miyamoto K, Hasegawa Y, Ibuki Y, Igarashi M, Minegishi T (Nov 1990). "Cloning and sequencing of human LH/hCG receptor cDNA". Biochemical and Biophysical Research Communications. 172 (3): 1049–54. doi:10.1016/0006-291X(90)91552-4. PMID 2244890.
- Rousseau-Merck MF, Misrahi M, Atger M, Loosfelt H, Milgrom E, Berger R (1991). "Localization of the human luteinizing hormone/choriogonadotropin receptor gene (LHCGR) to chromosome 2p21". Cytogenetics and Cell Genetics. 54 (1–2): 77–9. doi:10.1159/000132962. PMID 2249480.
- Xie YB, Wang H, Segaloff DL (Dec 1990). "Extracellular domain of lutropin/choriogonadotropin receptor expressed in transfected cells binds choriogonadotropin with high affinity". The Journal of Biological Chemistry. 265 (35): 21411–4. PMID 2254302.
- Frazier AL, Robbins LS, Stork PJ, Sprengel R, Segaloff DL, Cone RD (Aug 1990). "Isolation of TSH and LH/CG receptor cDNAs from human thyroid: regulation by tissue specific splicing". Molecular Endocrinology. 4 (8): 1264–76. doi:10.1210/mend-4-8-1264. PMID 2293030.
- Keutmann HT, Charlesworth MC, Mason KA, Ostrea T, Johnson L, Ryan RJ (Apr 1987). "A receptor-binding region in human choriogonadotropin/lutropin beta subunit". Proceedings of the National Academy of Sciences of the United States of America. 84 (7): 2038–42. doi:10.1073/pnas.84.7.2038. PMC 304579. PMID 3470775.
- Jiang X, Dreano M, Buckler DR, Cheng S, Ythier A, Wu H, Hendrickson WA, el Tayar N (Dec 1995). "Structural predictions for the ligand-binding region of glycoprotein hormone receptors and the nature of hormone-receptor interactions". Structure. 3 (12): 1341–53. doi:10.1016/S0969-2126(01)00272-6. PMID 8747461.
- Atger M, Misrahi M, Sar S, Le Flem L, Dessen P, Milgrom E (Jun 1995). "Structure of the human luteinizing hormone-choriogonadotropin receptor gene: unusual promoter and 5' non-coding regions". Molecular and Cellular Endocrinology. 111 (2): 113–23. doi:10.1016/0303-7207(95)03557-N. PMID 7556872.
- Latronico AC, Anasti J, Arnhold IJ, Mendonça BB, Domenice S, Albano MC, Zachman K, Wajchenberg BL, Tsigos C (Aug 1995). "A novel mutation of the luteinizing hormone receptor gene causing male gonadotropin-independent precocious puberty". The Journal of Clinical Endocrinology and Metabolism. 80 (8): 2490–4. doi:10.1210/jc.80.8.2490. PMID 7629248.
- Shenker A, Laue L, Kosugi S, Merendino JJ, Minegishi T, Cutler GB (Oct 1993). "A constitutively activating mutation of the luteinizing hormone receptor in familial male precocious puberty". Nature. 365 (6447): 652–4. doi:10.1038/365652a0. PMID 7692306.
- Yano K, Saji M, Hidaka A, Moriya N, Okuno A, Kohn LD, Cutler GB (Apr 1995). "A new constitutively activating point mutation in the luteinizing hormone/choriogonadotropin receptor gene in cases of male-limited precocious puberty". The Journal of Clinical Endocrinology and Metabolism. 80 (4): 1162–8. doi:10.1210/jc.80.4.1162. PMID 7714085.
- Kremer H, Kraaij R, Toledo SP, Post M, Fridman JB, Hayashida CY, van Reen M, Milgrom E, Ropers HH, Mariman E (Feb 1995). "Male pseudohermaphroditism due to a homozygous missense mutation of the luteinizing hormone receptor gene". Nature Genetics. 9 (2): 160–4. doi:10.1038/ng0295-160. hdl:2066/20602. PMID 7719343.
- Kosugi S, Van Dop C, Geffner ME, Rabl W, Carel JC, Chaussain JL, Mori T, Merendino JJ, Shenker A (Feb 1995). "Characterization of heterogeneous mutations causing constitutive activation of the luteinizing hormone receptor in familial male precocious puberty". Human Molecular Genetics. 4 (2): 183–8. doi:10.1093/hmg/4.2.183. PMID 7757065.
- Kremer H, Mariman E, Otten BJ, Moll GW, Stoelinga GB, Wit JM, Jansen M, Drop SL, Faas B, Ropers HH (Nov 1993). "Cosegregation of missense mutations of the luteinizing hormone receptor gene with familial male-limited precocious puberty". Human Molecular Genetics. 2 (11): 1779–83. doi:10.1093/hmg/2.11.1779. PMID 8281137.
External links
- "Glycoprotein Hormone Receptors: LH". IUPHAR Database of Receptors and Ion Channels. International Union of Basic and Clinical Pharmacology.
- SSFA-GPHR: Sequence Structure Function Analysis of Glycoprotein Hormone Receptors
- GRIS: Glycoprotein-hormone Receptor Information System
- LH-hCG+Receptors at the US National Library of Medicine Medical Subject Headings (MeSH)