Interleukin 15
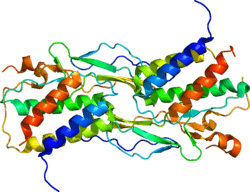
Interleukin-15 (IL-15) is a cytokine with structural similarity to Interleukin-2 (IL-2). Like IL-2, IL-15 binds to and signals through a complex composed of IL-2/IL-15 receptor beta chain (CD122) and the common gamma chain (gamma-C, CD132). IL-15 is secreted by mononuclear phagocytes (and some other cells) following infection by virus(es). This cytokine induces the proliferation of natural killer cells, i.e. cells of the innate immune system whose principal role is to kill virally infected cells.




Expression
IL-15 was discovered in 1994 by two different laboratories, and characterized as T cell growth factor.[5] Together with Interleukin-2 (IL-2), Interleukin-4 (IL-4), Interleukin-7 (IL-7), Interleukin-9 (IL-9), granulocyte colony-stimulating factor (G-CSF), and granulocyte-macrophage colony-stimulating factor (GM-CSF), IL-15 belongs to the four α-helix bundle family of cytokines.[5][6]
IL-15 is constitutively expressed by a large number of cell types and tissues, including monocytes, macrophages, dendritic cells (DC), keratinocytes, fibroblasts, myocyte and nerve cells.[7] As a pleiotropic cytokine, it plays an important role in innate and adaptive immunity.[8]
Gene

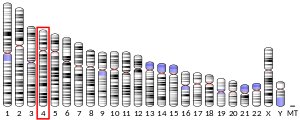
IL-15 is 14-15 kDa glycoprotein encoded by the 34 kb region of chromosome 4q31 in humans, and at the central region of chromosome 8 in mice.[9] The human IL-15 gene comprises nine exons (1 - 8 and 4A) and eight introns, four of which (exons 5 through 8) code for the mature protein (Figure 1).[5]
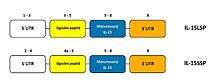
Two alternatively spliced transcript variants of this gene encoding the same protein have been reported.[10] The originally identified isoform, with long signal peptide of 48 amino acids (IL-15 LSP) consisted of a 316 bp 5’-untranslated region (UTR), 486 bp coding sequence and the C-terminus 400 bp 3’-UTR region. The other isoform (IL-15 SSP) has a short signal peptide of 21 amino acids encoded by exons 4A and 5.[5] Both isoforms shared 11 amino acids between signal sequences of the N-terminus.[11] Although both isoforms produce the same mature protein, they differ in their cellular trafficking.[5] IL-15 LSP isoform was identified in Golgi apparatus [GC], early endosomes and in the endoplasmic reticulum (ER). It exists in two forms, secreted and membrane-bound particularly on dendritic cells. On the other hand, IL-15 SSP isoform is not secreted and it appears to be restricted to the cytoplasm and nucleus where plays an important role in the regulation of cell cycle.[5]
It has been demonstrated that two isoforms of IL-15 mRNA are generated by alternatively splicing in mice. The isoform which had an alternative exon 5 containing another 3’ splicing site, exhibited a high translational efficiency, and the product lack hydrophobic domains in the signal sequence of the N-terminus. This suggests that the protein derived from this isoform is located intracellulary. The other isoform with normal exon 5, which is generated by integral splicing of the alternative exon 5, may be released extracellulary.[12]
Although IL-15 mRNA can be found in many cells and tissues including mast cells, cancer cells or fibroblasts, this cytokine is produced as a mature protein mainly by dendritic cells, monocytes and macrophages. This discrepancy between the wide appearance of IL-15 mRNA and limited production of protein might be explained by the presence of the twelve in humans and five in mice upstream initiating codons, which can repress translation of IL-15 mRNA. Translational inactive mRNA is stored within the cell and can be induced upon specific signal.[13] Expression of IL-15 can be stimulated by cytokine such as GM-CSF, double-strand mRNA, unmethylated CpG oligonucleotides, lipopolysaccharide (LPS) through Toll-like receptors (TLR), interferon gamma (IFN-γ) or after infection of monocytes herpes virus, Mycobacterium tuberculosis and Candida albicans (Figure 2).[14]
Signaling
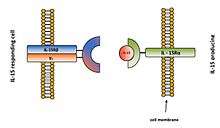
The prevailing mechanism of IL-15 action seems to be juxtacrine signaling or also determined as cell-to-cell contact. It also includes intracrine and reverse signaling. IL-15 was initially characterized as a soluble molecule. Later it was shown that IL-15 also exists as a membrane-bound form which represents the major form of IL-15 protein. In membrane-bound form it could be bound directly to cellular membrane or presented by IL-15Rα receptor.[13]
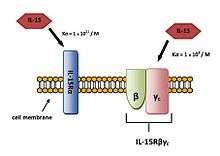
The main mechanism of IL-15 signaling is trans-presentation which is mediated by membrane-bound complex IL-15/IL-15Rα (Figure 3).[15] IL-15 bind to IL-15Rα receptor alone with affinity (Ka = 1.1011/M). It can also bind to IL-15Rβγc signaling complex with lower affinity (Ka = 1.109/M) (Figure 4).[8]
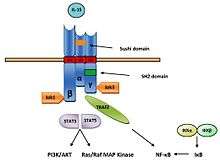
Signaling pathway of IL-15 begins with binding to IL-15Rα receptor, with subsequent presentation to surrounding cells bearing IL-15Rβγc complex on their cell surface. Upon binding IL-15β subunit activates Janus kinase 1 (Jak1) and γc subunit Janus kinase 3 (Jak3), which leads to phosphorylation and activation of signal transducer and activator of transcription 3 (STAT3) and STAT5.[16] Due to sharing of receptor subunits between IL-2 and IL-15, both of these cytokines have similar downstream effects including the induction of Bcl-2, MAP (mitogen-activated protein kinase) kinase pathway and the phosphorylation of Lck (lymphocyte-activated protein tyrosine kinase) and Syk (spleen tyrosine kinase) kinases, which leads to cell proliferation and maturation (Figure 5).[8][17]
In mast cells, the IL-15R signaling pathway has been found to include Jak2 and STAT5 instead Jak1/3 and STAT3/5. Phosphorylation STATs form transcription factors and activate transcription of appropriate genes. The β chain of IL-15R recruits and also activates protein tyrosine kinases of the Src family including Lck, Fyn and Lyn kinase. It also activates phosphatidylinositol 3-kinase (PI3K) and AKT signaling pathway and induce expression of transcription factors including c-Fos, c-Jun, c-Myc and NF-κB.[13]
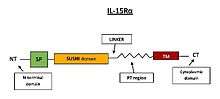
IL-15 is also able to bind to the 15Rβγc signaling complex with intermediate affinity without requirement for IL-15Rα receptor. Upon binding IL-15 to signaling complex, kinases of the Src family including Lck and Fyn are activated, and subsequently activates PI3K and MAPK signaling pathway.[18] The second mechanism of IL-15 action is cis-presentation, when IL-15 is presented by IL-15Rα to 15Rβγc signaling complex on the same cell. This mechanism is mediated by the C-terminus flexibility which is mediated by 32 amino acids linker and/or 74 amino acids long PT region (Figure 6).[15]
Function
IL-15 regulates the activation and proliferation of T and natural killer (NK) cells. Survival signals that maintain memory T cells in the absence of antigen are provided by IL-15. This cytokine is also implicated in NK cell development. In rodent lymphocytes, IL-15 prevents apoptosis by inducing BCL2L1/BCL-x(L), an inhibitor of the apoptosis pathway.[10] In humans with celiac disease IL-15 similarly suppresses apoptosis in T-lymphocytes by inducing Bcl-2 and/or Bcl-xL.[19]
A hematopoietin receptor, the IL-15 receptor, that binds IL-15 propagates its function. Some subunits of the IL-15 receptor are shared in common with the receptor for a structurally related cytokine called Interleukin 2 (IL-2) allowing both cytokines to compete for and negatively regulate each other's activity. CD8+ memory T cell number is controlled by a balance between IL-15 and IL-2. When IL-15 binds its receptor, JAK kinase, STAT3, STAT5, and STAT6 transcription factors are activated to elicit downstream signaling events.
IL-15 and its receptor subunit alpha (IL-15Rα) are also produced by skeletal muscle in response to different exercise doses (myokine), playing significant roles in visceral (intra-abdominal or interstitial) fat reduction [20][21] and myofibrillar protein synthesis (hypertrophy).[22]
Disease
Epstein-Barr virus
In humans with history of acute infectious mononucleosis (the syndrome associated with primary Epstein-Barr virus infection), IL-15R expressing lymphocytes are not detected even 14 years after infection.[23]
Celiac disease
There have been recent studies suggesting that suppression of IL-15 may be a potential treatment for celiac disease and even presents the possibility of preventing its development. In one study with mice blocking IL-15 with an antibody led to the reversal of autoimmune intestinal damage.[24] In another study mice used were able to eat gluten without developing symptoms.[25]
Non-alcoholic fatty liver disease
A recent report indicated IL-15 promotes non-alcoholic fatty liver disease.[26]
Immunotherapy
Metastatic cancer
IL-15 has been shown to enhance the anti-tumor immunity of CD8+ T cells in pre-clinical models.[27][28] A phase I clinical trial to evaluate the safety, dosing, and anti-tumor efficacy of IL-15 in patients with metastatic melanoma and renal cell carcinoma (kidney cancer) has begun to enroll patients at the National Institutes of Health.[29]
Vaccines Adjuvants
Vector-based therapy – Nonlytic Newcastle Disease Virus (NDV) was engineered to express recombinant IL-15 protein to generate an NDV-modified tumor vaccine. Preclinical results of NDV-modified tumor vaccine showed promise by controlling melanoma tumor growth in mice.[30] A recombinant vaccinia virus expressing influenza A proteins and IL-15 promoted cross protection by CD4+ T cells.[31] A Brucella DNA vaccine containing IL-15 gene enhanced the CD8+ T cell immune response in mice.[32] IL-15 was needed for CD4+ T cell heterosubtypic protection while using a multivalent influenza vaccine using vaccinia-based vector.[31] While influenza A virus expressing IL-15 stimulates both innate and adaptive immune cells to decrease tumor growth mice.[33]
Transpresentation complexes
Currently there are two varieties of IL-15 superagonist available. One combines IL-15 and IL-15Rα-Fc (R&D Systems) in vitro to generate the complex. It is referred to as IL-15 SA. A second IL-15 superagonist complex called ALT-803 is offered by Altor BioScience.
IL-15 SA
IL-15 SA is currently being evaluated for antiviral and anticancer activities, in addition to enhancing immunotherapy and vaccination.[34][35] One potential shortcoming of IL-15 SA was its enhancement of septic shock in mice.[36]
ALT-803
ALT-803 is an IL-15 superagonist complex IL-15N72D:IL-15RαSu/Fc that includes an IL-15 mutant (IL-15N72D) and a dimeric IL-15 receptor α sushi domain-IgG1 Fc fusion protein.[37][38]
ALT-803 was given fast track status by the FDA in 2017 and at that time, Phase III trials in bladder cancer were being prepared.[39]
RLI-15
RLI-15 is an fusion protein consisting of the NH2-terminal (amino acids 1–77, sushi+) cytokine-binding domain of IL-15Rα coupled to IL-15 via a 20-amino acid flexible linker. This fusion protein, referred to as protein receptor-linker-IL-15 (RLI) acts as an IL-15 superagonists that has an increased serum half-life and biological activity similar to complexed IL-15/IL-15Rα-Fc. RLI demonstrated a strong anti-tumor effect in two different tumor models.[40] RLI15 is being produced and tested by Cytune Pharma affiliated company of Sotio which renamed it to SO-C101.[41] Phase 1 trial was initiated in 2019.[42]
References
- GRCh38: Ensembl release 89: ENSG00000164136 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000031712 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Steel JC, Waldmann TA, Morris JC (January 2012). "Interleukin-15 biology and its therapeutic implications in cancer". Trends in Pharmacological Sciences. 33 (1): 35–41. doi:10.1016/j.tips.2011.09.004. PMC 3327885. PMID 22032984.
- Di Sabatino A, Calarota SA, Vidali F, Macdonald TT, Corazza GR (February 2011). "Role of IL-15 in immune-mediated and infectious diseases". Cytokine & Growth Factor Reviews. 22 (1): 19–33. doi:10.1016/j.cytogfr.2010.09.003. PMID 21074481.
- Grabstein KH, Eisenman J, Shanebeck K, Rauch C, Srinivasan S, Fung V, Beers C, Richardson J, Schoenborn MA, Ahdieh M (May 1994). "Cloning of a T cell growth factor that interacts with the beta chain of the interleukin-2 receptor". Science. 264 (5161): 965–8. doi:10.1126/science.8178155. PMID 8178155.
- Lodolce JP, Burkett PR, Koka RM, Boone DL, Ma A (December 2002). "Regulation of lymphoid homeostasis by interleukin-15". Cytokine & Growth Factor Reviews. 13 (6): 429–39. doi:10.1016/S1359-6101(02)00029-1. PMID 12401478.
- Waldmann TA, Tagaya Y (1999). "The multifaceted regulation of interleukin-15 expression and the role of this cytokine in NK cell differentiation and host response to intracellular pathogens". Annual Review of Immunology. 17: 19–49. doi:10.1146/annurev.immunol.17.1.19. PMID 10358752.
- "Entrez Gene: IL15 interleukin 15".
- Tagaya Y, Kurys G, Thies TA, Losi JM, Azimi N, Hanover JA, Bamford RN, Waldmann TA (December 1997). "Generation of secretable and nonsecretable interleukin 15 isoforms through alternate usage of signal peptides". Proceedings of the National Academy of Sciences of the United States of America. 94 (26): 14444–9. doi:10.1073/pnas.94.26.14444. PMC 25016. PMID 9405632.
- Nishimura H, Yajima T, Naiki Y, Tsunobuchi H, Umemura M, Itano K, Matsuguchi T, Suzuki M, Ohashi PS, Yoshikai Y (January 2000). "Differential roles of interleukin 15 mRNA isoforms generated by alternative splicing in immune responses in vivo". The Journal of Experimental Medicine. 191 (1): 157–70. doi:10.1084/jem.191.1.157. PMC 2195806. PMID 10620614.
- Jakobisiak M, Golab J, Lasek W (April 2011). "Interleukin 15 as a promising candidate for tumor immunotherapy". Cytokine & Growth Factor Reviews. 22 (2): 99–108. doi:10.1016/j.cytogfr.2011.04.001. PMID 21531164.
- Bamford RN, DeFilippis AP, Azimi N, Kurys G, Waldmann TA (May 1998). "The 5' untranslated region, signal peptide, and the coding sequence of the carboxyl terminus of IL-15 participate in its multifaceted translational control". Journal of Immunology. 160 (9): 4418–26. PMID 9574546.
- Olsen SK, Ota N, Kishishita S, Kukimoto-Niino M, Murayama K, Uchiyama H, Toyama M, Terada T, Shirouzu M, Kanagawa O, Yokoyama S (December 2007). "Crystal Structure of the interleukin-15.interleukin-15 receptor alpha complex: insights into trans and cis presentation". The Journal of Biological Chemistry. 282 (51): 37191–204. doi:10.1074/jbc.M706150200. PMID 17947230.
- Okada S, Han S, Patel ES, Yang LJ, Chang LJ (2015). "STAT3 signaling contributes to the high effector activities of interleukin-15-derived dendritic cells". Immunology and Cell Biology. 93 (5): 461–71. doi:10.1038/icb.2014.103. PMC 4450366. PMID 25582338.
- Schluns KS, Stoklasek T, Lefrançois L (August 2005). "The roles of interleukin-15 receptor alpha: trans-presentation, receptor component, or both?". The International Journal of Biochemistry & Cell Biology. 37 (8): 1567–71. doi:10.1016/j.biocel.2005.02.017. PMID 15896666.
- Perera PY, Lichy JH, Waldmann TA, Perera LP (March 2012). "The role of interleukin-15 in inflammation and immune responses to infection: implications for its therapeutic use". Microbes and Infection. 14 (3): 247–61. doi:10.1016/j.micinf.2011.10.006. PMC 3270128. PMID 22064066.
- Malamut G, El Machhour R, Montcuquet N, Martin-Lannerée S, Dusanter-Fourt I, Verkarre V, Mention JJ, Rahmi G, Kiyono H, Butz EA, Brousse N, Cellier C, Cerf-Bensussan N, Meresse B (June 2010). "IL-15 triggers an antiapoptotic pathway in human intraepithelial lymphocytes that is a potential new target in celiac disease-associated inflammation and lymphomagenesis". The Journal of Clinical Investigation. 120 (6): 2131–43. doi:10.1172/JCI41344. PMC 2877946. PMID 20440074.
- Pedersen BK (January 2011). "Muscles and their myokines". The Journal of Experimental Biology. 214 (Pt 2): 337–46. doi:10.1242/jeb.048074. PMID 21177953.
- Pérez-López A, Valadés D, Vázquez Martínez C, de Cos Blanco AI, Bujan J, García-Honduvilla N (March 2018). "Serum IL-15 and IL-15Rα levels are decreased in lean and obese physically active humans". Scandinavian Journal of Medicine & Science in Sports. 28 (3): 1113–1120. doi:10.1111/sms.12983. PMID 28940555.
- Pérez-López A, McKendry J, Martin-Rincon M, Morales-Alamo D, Pérez-Köhler B, Valadés D, Buján J, Calbet JA, Breen L (January 2018). "Skeletal muscle IL-15/IL-15Rα and myofibrillar protein synthesis after resistance exercise" (PDF). Scandinavian Journal of Medicine & Science in Sports. 28 (1): 116–125. doi:10.1111/sms.12901. PMID 28449327.
- Sauce D, Larsen M, Curnow SJ, Leese AM, Moss PA, Hislop AD, Salmon M, Rickinson AB (July 2006). "EBV-associated mononucleosis leads to long-term global deficit in T-cell responsiveness to IL-15". Blood. 108 (1): 11–8. doi:10.1182/blood-2006-01-0144. PMID 16543467.
- DePaolo RW, Abadie V, Tang F, Fehlner-Peach H, Hall JA, Wang W, Marietta EV, Kasarda DD, Waldmann TA, Murray JA, Semrad C, Kupfer SS, Belkaid Y, Guandalini S, Jabri B (March 2011). "Co-adjuvant effects of retinoic acid and IL-15 induce inflammatory immunity to dietary antigens". Nature. 471 (7337): 220–4. doi:10.1038/nature09849. PMC 3076739. PMID 21307853. Lay summary – WebMD Health News.
- Yokoyama S, Watanabe N, Sato N, Perera PY, Filkoski L, Tanaka T, Miyasaka M, Waldmann TA, Hiroi T, Perera LP (September 2009). "Antibody-mediated blockade of IL-15 reverses the autoimmune intestinal damage in transgenic mice that overexpress IL-15 in enterocytes". Proceedings of the National Academy of Sciences of the United States of America. 106 (37): 15849–54. doi:10.1073/pnas.0908834106. PMC 2736142. PMID 19805228.
- Cepero-Donates Y, Lacraz G, Ghobadi F, Rakotoarivelo V, Orkhis S, Mayhue M, Chen YG, Rola-Pleszczynski M, Menendez A, Ilangumaran S, Ramanathan S (June 2016). "Interleukin-15-mediated inflammation promotes non-alcoholic fatty liver disease". Cytokine. 82: 102–11. doi:10.1016/j.cyto.2016.01.020. PMID 26868085.
- Klebanoff CA, Finkelstein SE, Surman DR, Lichtman MK, Gattinoni L, Theoret MR, Grewal N, Spiess PJ, Antony PA, Palmer DC, Tagaya Y, Rosenberg SA, Waldmann TA, Restifo NP (February 2004). "IL-15 enhances the in vivo antitumor activity of tumor-reactive CD8+ T cells". Proceedings of the National Academy of Sciences of the United States of America. 101 (7): 1969–74. doi:10.1073/pnas.0307298101. PMC 357036. PMID 14762166.
- Teague RM, Sather BD, Sacks JA, Huang MZ, Dossett ML, Morimoto J, Tan X, Sutton SE, Cooke MP, Ohlén C, Greenberg PD (March 2006). "Interleukin-15 rescues tolerant CD8+ T cells for use in adoptive immunotherapy of established tumors". Nature Medicine. 12 (3): 335–41. doi:10.1038/nm1359. PMID 16474399.
- "A Phase I Study of Intravenous Recombinant Human IL-15 in Adults With Refractory Metastatic Malignant Melanoma and Metastatic Renal Cell Cancer". ClinicalTrials.gov.
- Xu X, Sun Q, Yu X, Zhao L (April 2017). "Rescue of nonlytic Newcastle Disease Virus (NDV) expressing IL-15 for cancer immunotherapy". Virus Research. 233: 35–41. doi:10.1016/j.virusres.2017.03.003. PMID 28286036.
- Valkenburg SA, Li OT, Mak PW, Mok CK, Nicholls JM, Guan Y, Waldmann TA, Peiris JS, Perera LP, Poon LL (April 2014). "IL-15 adjuvanted multivalent vaccinia-based universal influenza vaccine requires CD4+ T cells for heterosubtypic protection". Proceedings of the National Academy of Sciences of the United States of America. 111 (15): 5676–81. doi:10.1073/pnas.1403684111. PMC 3992686. PMID 24706798.
- Hu XD, Chen ST, Li JY, Yu DH, Cai H (March 2010). "An IL-15 adjuvant enhances the efficacy of a combined DNA vaccine against Brucella by increasing the CD8+ cytotoxic T cell response". Vaccine. 28 (12): 2408–15. doi:10.1016/j.vaccine.2009.12.076. PMID 20064480.
- Hock K, Laengle J, Kuznetsova I, Egorov A, Hegedus B, Dome B, Wekerle T, Sachet M, Bergmann M (March 2017). "Oncolytic influenza A virus expressing interleukin-15 decreases tumor growth in vivo". Surgery. 161 (3): 735–746. doi:10.1016/j.surg.2016.08.045. PMID 27776794.
- Ahmad A, Ahmad R, Iannello A, Toma E, Morisset R, Sindhu ST (July 2005). "IL-15 and HIV infection: lessons for immunotherapy and vaccination". Current HIV Research. 3 (3): 261–70. doi:10.2174/1570162054368093. PMID 16022657.
- Suck G, Oei VY, Linn YC, Ho SH, Chu S, Choong A, Niam M, Koh MB (September 2011). "Interleukin-15 supports generation of highly potent clinical-grade natural killer cells in long-term cultures for targeting hematological malignancies". Experimental Hematology. 39 (9): 904–14. doi:10.1016/j.exphem.2011.06.003. PMID 21703984.
- Guo Y, Luan L, Patil NK, Wang J, Bohannon JK, Rabacal W, Fensterheim BA, Hernandez A, Sherwood ER (February 2017). "IL-15 Enables Septic Shock by Maintaining NK Cell Integrity and Function". Journal of Immunology. 198 (3): 1320–1333. doi:10.4049/jimmunol.1601486. PMC 5263185. PMID 28031340.
- "Altor BioScience". altorbioscience.com. Retrieved 2018-11-08.
- Liu B, Kong L, Han K, Hong H, Marcus WD, Chen X, et al. (November 2016). "A Novel Fusion of ALT-803 (Interleukin (IL)-15 Superagonist) with an Antibody Demonstrates Antigen-specific Antitumor Responses". The Journal of Biological Chemistry. 291 (46): 23869–23881. doi:10.1074/jbc.M116.733600. PMC 5104912. PMID 27650494.
- Adams B (June 28, 2017). "Altor shareholders revolt against Soon-Shiong buyout". FierceBiotech.
- Robinson TO, Schluns KS (October 2017). "The potential and promise of IL-15 in immuno-oncogenic therapies". Immunology Letters. 190: 159–168. doi:10.1016/j.imlet.2017.08.010. PMC 5774016. PMID 28823521.
- "SO-C101 - Sotio". www.sotio.com. Retrieved 16 August 2019.
- "SOTIO initiates first-in-human clinical trial with IL-15 superagonist SO-C101". www.sotio.com. Retrieved 16 August 2019.
Further reading
- Ren H, Han R, Chen X, Liu X, Wan J, Wang L, Yang X, Wang J (May 2020). "Potential therapeutic targets for intracerebral hemorrhage-associated inflammation: An update". J Cereb Blood Flow Metab: 0271678X2092355. doi:10.1177/0271678X20923551. PMID 32423330.
- Maślińska D (2001). "The cytokine network and interleukin-15 (IL-15) in brain development". Folia Neuropathologica. 39 (2): 43–7. PMID 11680634.
- Liew FY, McInnes IB (November 2002). "Role of interleukin 15 and interleukin 18 in inflammatory response". Annals of the Rheumatic Diseases. 61 Suppl 2 (Suppl 2): ii100-2. doi:10.1136/ard.61.suppl_2.ii100. PMC 1766710. PMID 12379638.
- Lodolce JP, Burkett PR, Koka RM, Boone DL, Ma A (December 2002). "Regulation of lymphoid homeostasis by interleukin-15". Cytokine & Growth Factor Reviews. 13 (6): 429–39. doi:10.1016/S1359-6101(02)00029-1. PMID 12401478.
- Mattei F, Schiavoni G, Belardelli F, Tough DF (August 2001). "IL-15 is expressed by dendritic cells in response to type I IFN, double-stranded RNA, or lipopolysaccharide and promotes dendritic cell activation". Journal of Immunology. 167 (3): 1179–87. doi:10.4049/jimmunol.167.3.1179. PMID 11466332.