Oncostatin M
Oncostatin M, also known as OSM, is a protein that in humans is encoded by the OSM gene.[4]



OSM is a pleiotropic cytokine that belongs to the interleukin 6 group of cytokines.[5] Of these cytokines it most closely resembles leukemia inhibitory factor (LIF) in both structure and function.[5] As yet poorly defined, it is proving important in liver development, haematopoeisis, inflammation and possibly CNS development. It is also associated with bone formation and destruction.[6]
OSM signals through cell surface receptors that contain the protein gp130. The type I receptor is composed of gp130 and LIFR, the type II receptor is composed of gp130 and OSMR.[7]
Discovery, isolation and cloning
The human form of OSM was originally isolated in 1986 from the growth media of PMA treated U-937 histiocytic lymphoma cells by its ability to inhibit the growth of cell lines established from melanomas and other solid tumours.[8] A robust protein, OSM is stable between pH2 and 11 and resistant to heating for one hour at 56 °C. A partial amino acid sequence allowed the isolation of human OSM cDNA and subsequently genomic clones.[9] The full cDNA clone of hOSM encodes a 252 amino acid precursor, the first 25 amino acids of which functions as a secretory signal peptide, which on removal yields the soluble 227 amino acid pro-OSM. Cleavage of the C-terminal most 31 residues at a trypsin like cleavage site yields the fully active 196 residue form. Two potential N-glycosylation sites are present in hOSM both of which are retained in the mature form.[10][11]
The 196 residue OSM is the predominant form isolated form a variety of cell lines and corresponds to a glycoprotein of 28 KDa, although the larger 227 residue pro-OSM can be isolated from over transfected cells. Pro-OSM, although an order of magnitude less efficacious in growth inhibition assays, displays similar binding affinity toward cells in radio ligand binding assays.[11] Thus, post translational processing may play a significant role in the in vivo function of OSM. Like many cytokines OSM is produced from cells by de novo synthesis followed by release through the classical secretion pathway. However, OSM can be released from preformed stores within polymorphonuclear leukocytes on degranulation.[12] It still remains unclear how OSM is targeted to these intracellular compartments.
Structure
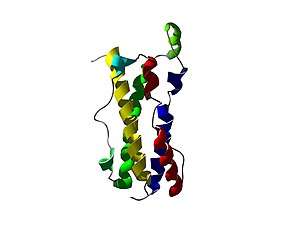
Primary sequence analysis of OSM allocates it to the gp130 group of cytokines. OSM most resembles LIF, bearing 22% sequence identity and 30% similarity. Incidentally the genes for OSM and LIF occur in tandem on human chromosome 22. Both LIF and OSM genes have very similar gene structures sharing similar promoter elements and intron-exon structure.[14] These data suggest that OSM and LIF arose relatively recently in evolutionary terms by gene duplication.[4] Of the five cysteine residues within the human OSM sequence four participate in disulfide bridges, one of these disulfide bonds namely between helices A and B is necessary for OSM activity. The free cysteine residue does not appear to mediate dimerisation of OSM.
The three-dimensional structure of human OSM has been solved to atomic resolution, confirming the predicted long chain four helix bundle topology.[13] Comparing this structure with the known structures of other known LC cytokines shows it to be most closely related to LIF (RMSD of 2.1 Å across 145 equivalent Cα). A distinctive kink in the A helix arises from departure of the classical alpha helical H-bonding pattern, a feature shared with all known structures of LIFR using cytokines. This “kink” results in a different special positioning of one extreme of the bundle to the other, significantly affecting the relative positioning of site III with sites I and II (see:Receptor recruitment sites)
Receptors
Receptors for OSM can be found on a variety of cells from a variety of tissues. In general cells derived from endothelial and tumour origins express high levels of OSM receptors, whereas cells of Haematopoietic origin tend to express lower numbers. Scatchard analysis of radio ligand binding data from 125I-OSM binding to a variety of OSM responsive cell lines produced curvilinear graphs which the authors interpreted as the presence of two receptor species, a high affinity form with an approximate dissociation constant Kd of 1-10 pM, and a low affinity form of 0.4-1 nM.[15] Subsequently it was shown that the presence of gp130 alone was sufficient to reproduce the low affinity form of the receptor, and co-transfection of COS-7 cells with LIFR and gp130 produced a high affinity receptor.[16] However further experiments demonstrated that not all actions of OSM could be replicated by LIF, that is certain cells that are irresponsive to LIF would respond to OSM.[17] This data hinted to the existence of an additional ligand specific receptor chain which led to the cloning of OSMR.[18] These two receptor complexes, namely gp130/LIFR and gp130/OSMR, were termed the type I and type II Oncostatin-M receptors. The ability of OSM to signal via two receptor complexes conveniently offers a molecular explanation to the shared and unique effects of OSM with respect to LIF. Thus common biological activities of LIF and OSM are mediated through the type I receptor and OSM specific activities are mediated through the type II receptor.
The murine homologue of OSM was not discovered until 1996,[19] whereas the murine OSMR homologue was not cloned until 1998.[20] Until recently, it was thought that mOSM only signals through the murine type II receptor, namely through mOSMR/mgp130 complexes, because of a low affinity for the type I receptor counterpart.[21] However, it is now known that, in bone at least, mOSM is able to signal through both mOSMR/mgp130 and mLIFR/mgp130.[6]
Receptor recruitment sites
Oncostatin M triggers the formation of receptor complexes by binding to receptors via two binding sites named site II and site III. The nomenclature of these sites is taken by direct analogy to Growth Hormone, probably the best studied of four helix bundle cytokines.
Site II consists of exposed residues within the A and C helices, and confers binding to gp130. The crucial residues of site III are located at the N-terminal extremity of the D-helix. This site is the most conserved amongst IL-6 like cytokines. OSM contains a conserved Phenylalanine and Lysine residues (F160 and K163). Cytokines that recruit LIFR via site 3 i.e. LIF, OSM, CNTF and CT-1 possess these conserved phenylalanine and lysine residues and is known as the FK motif.
Signal transduction through OSM receptors
Signalling by type I and type II OSM receptors have now been shown to be qualitatively distinct. These differences in signaling character, in addition to the tissue distribution profiles of OSMRb and LIFRb, offer another variable in the distinction between the common and specific cellular effects of OSM with respect to LIF. All IL-6 cytokines whether they homo- or heterodimerise gp130 seem to activate JAK1, JAK2 and to a lesser degree Tyk2.[7][22] JAK1, JAK2, and tyk2 are not interchangeable in the gp130 system, this has been demonstrated with the use of JAK1, Jak2 or Tyk2 deficient cell lines obtained from mutant mice. Cells from JAK1 deficient mice show reduced STAT activation and generation of biological responses in response to IL-6 and LIF.[23] In contrast, fibroblasts derived from JAK2 null mice can respond to IL-6, with demonstratable tyrosine phosphorylation of gp130, JAK1 and TYK2.[24] Thus it seems JAK1 is the critical JAK required for gp130 signalling. Activation of the same Jaks by all three receptor combinations (gp130/gp130, gp130/LIFR, gp130/OSMR) raises the question of how IL6, LIF and OSM can activate distinct intracellular signaling pathways. Selection of particular substrates, i.e. STAT isoform, depended not on which Jak is activated, but instead are determined by specific motifs, especially tyrosine-based motifs, within each receptor intracellular domain.
Aligning the intracellular domains of gp130, LIFR and hOSMR results in some interesting observations. Sequence identity is generally quite low across the group averaging at 4.6%. However, as with many Class I Haematopoeitin receptors, two short membrane proximal motifs, termed box 1 and box 2 are present. In addition these receptors also contain a serine rich region and a third more poorly conserved motif termed box 3. Box 1 is present in all signalling cytokine receptors. It is characteristically rich in proline residues and is essential for the association and activation of JAKs.[25] Box 2 is also important for association with JAKs. Gp130 contains box1 and box2 sequences within the membrane-proximal part of the cytoplasmic region, lying within the minimum 61 amino acids required for receptor activation.[26] Mutations within the box1 region reduce the ability of gp130 to associate with Jaks[27] and abolish ligand-induced activation of Jak1 and Jak2.[26][28] Box 2 also contributes to activation and binding of JAKs. Studies with various gp130 truncation mutants show a reduction of Jak2 binding and abrogation of certain biological effects upon deletion of box2.[26][29] However, Jaks are able to associate with gp130 devoid of box2 when overexpressed.[27]
LIFR and OSMR also contain the membrane-proximal box1/box2-like regions. The first 65 amino acid residues in the cytoplasmic domain of LIFR, in combination with full length gp130, can generate signalling on treatment with LIF.[30] Coprecipitation of Jak1, Jak2 and Tyk2 with receptors containing cytoplasmic parts of the LIFR[31] or OSMR.[7] All beta receptor subunits of the gp130 system also possess a box 3 region. This region corresponds to the C-terminal amino acids of the OSMR and LIFR receptors respectively. Box 3 is necessary for the action of OSMR; however Box3 is dispensable for the action of LIFR.[32] In the case of gp130 box 3 is dispensable for activity, however the presence of an intact box 3 sequence is required for certain aspects of gp130 signalling, i.e. stimulation of transcription through the STAT-3 response element. In addition to the poor sequence conservation amongst the intracellular domains of gp130 receptors, the number and position of conserved tyrosine residues are also poorly conserved. For example, LIFR and OSMR share three homologous tyrosines. In contrast none of the tyrosine residues present in the intracellular domain of gp130 share equivalents with LIFR or OSMR, even though the intracellular regions of LIFR and gp130 share more sequence identity than LIFR and OSMR.
Of the proteins recruited to type I cytokine receptors STAT proteins remain the best studied. Homodimerisation of gp130 has been shown to phosphorylate and activate both STAT1 and STAT3. gp130 preferentially activates STAT3 which it can do through four STAT3 activation consensus sequences YXXQ: (YRHQ), (YFKQ), Y905 (YLPQ), Y915 (YMPQ). The lower propensity for STAT1 activation may be a reflection of the lower number of STAT1 activation sequences, YZPQ (where X is any residue and Z is any uncharged residue), namely Y905 and Y915.[33] Cytokines that signal via homodimeric complexes of LIFR or OSMR (i.e. devoid of gp130) are currently unknown in nature. However, various investigators have attempted artificial homodimerisation of LIFR and OSMR intracellular domains, with conflicting results, by constructing receptor chimeras that fuse the extracellular domain of one cytokine receptor with the intracellular domain of LIFR or OSMR.
Signalling by LIFR intracellular domain homodimerisation has been demonstrated in hepatoma and neuroblastoma cells,[30] embryonic stem cells[34][35] and COS-1 cells[36] by using chimeric receptors that homodimerise upon stimulation with their cognate cytokines (i.e. GCSF, neurotrophin-3, EGF). However a GCSFR/LIFR chimera was not capable of signaling in M1 or Baf cells.[35]
Anti- or pro-inflammatory?
The role of OSM as an inflammatory mediator was clear as early as 1986.[8] Its precise effect on the immune system, as with any cytokine, is far from clear. However, two schools of thought are emerging: The first proposes that OSM is pro-inflammatory; whilst the other holds the opposite view, claiming OSM is anti-inflammatory. It is important to note that before 1997[37] differences in human and murine OSM receptor usage were unknown. As a result, several investigators used human OSM in mouse assays and thus any conclusion drawn from the results of these experiments will be representative of LIF, i.e. signalling through gp130/LIFR complexes.
OSM is synthesized by stimulated T-cells and monocytes.[9] The effects of OSM on endothelial cells suggest a pro-inflammatory role for OSM. Endothelial cells possess a large number of OSM receptors.[38] Stimulation of a primary endothelial culture (HUVEC) with hOSM results in delayed but prolonged upregulation of P-selectin,[39] which facilitates leukocyte adhesion and rolling, necessary for their extravasation. OSM also promotes the production of IL-6 from these cells.[38]
As mentioned above the action of OSM as a quencher of the inflammatory response is by no means established yet. For example, conflicting results exist as to the action of OSM on various models of arthritis. For example, OSM reduces the degree of joint destruction in an antibody induced model of rheumatoid arthritis.[40]
OSM is a major growth factor for Kaposi’s sarcoma “spindle cells”, which are of endothelial origin.[41] These cells do not express LIFR but do express OSMR at high levels.[42] For example, OSM can modulate the expression of IL-6, an important regulator of the host defence system.[38] OSM can regulate the expression of acute phase proteins.[43] OSM regulates the expression of various protease and protease inhibitors, for example Gelatinase and a1-chymotrypsin inhibitor.
See also
References
- GRCh38: Ensembl release 89: ENSG00000099985 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Rose TM, Bruce AG (October 1991). "Oncostatin M is a member of a cytokine family that includes leukemia-inhibitory factor, granulocyte colony-stimulating factor, and interleukin 6". Proc. Natl. Acad. Sci. U.S.A. 88 (19): 8641–5. doi:10.1073/pnas.88.19.8641. PMC 52565. PMID 1717982.
- Tanaka M, Miyajima A (2003). "Oncostatin M, a multifunctional cytokine". Rev. Physiol. Biochem. Pharmacol. Reviews of Physiology, Biochemistry and Pharmacology. 149: 39–52. doi:10.1007/s10254-003-0013-1. ISBN 978-3-540-20213-4. PMID 12811586.
- Walker EC, McGregor NE, Poulton IJ, Solano M, Pompolo S, Fernandes TJ, Constable MJ, Nicholson GC, Zhang JG, Nicola NA, Gillespie MT, Martin TJ, Sims NA (2010). "Oncostatin M promotes bone formation independently of resorption when signaling through leukemia inhibitory factor receptor in mice". J Clin Invest. 120 (2): 582–92. doi:10.1172/JCI40568. PMC 2810087. PMID 20051625. Lay summary – ScienceDaily.
- Auguste P, Guillet C, Fourcin M, Olivier C, Veziers J, Pouplard-Barthelaix A, Gascan H (June 1997). "Signaling of type II oncostatin M receptor". J. Biol. Chem. 272 (25): 15760–4. doi:10.1074/jbc.272.25.15760. PMID 9188471.
- Zarling JM, Shoyab M, Marquardt H, Hanson MB, Lioubin MN, Todaro GJ (December 1986). "Oncostatin M: a growth regulator produced by differentiated histiocytic lymphoma cells". Proc. Natl. Acad. Sci. U.S.A. 83 (24): 9739–43. doi:10.1073/pnas.83.24.9739. PMC 387216. PMID 3540948.
- Malik N, Kallestad JC, Gunderson NL, Austin SD, Neubauer MG, Ochs V, Marquardt H, Zarling JM, Shoyab M, Wei CM (July 1989). "Molecular cloning, sequence analysis, and functional expression of a novel growth regulator, oncostatin M". Mol. Cell. Biol. 9 (7): 2847–53. PMC 362750. PMID 2779549.
- Linsley PS, Kallestad J, Ochs V, Neubauer M (May 1990). "Cleavage of a hydrophilic C-terminal domain increases growth-inhibitory activity of oncostatin M". Mol. Cell. Biol. 10 (5): 1882–90. PMC 360533. PMID 2325640.
- Malik N, Graves D, Shoyab M, Purchio AF (1992). "Amplification and expression of heterologous oncostatin M in Chinese hamster ovary cells". DNA Cell Biol. 11 (6): 453–9. doi:10.1089/dna.1992.11.453. PMID 1524679.
- Grenier A, Dehoux M, Boutten A, Arce-Vicioso M, Durand G, Gougerot-Pocidalo MA, Chollet-Martin S (February 1999). "Oncostatin M production and regulation by human polymorphonuclear neutrophils". Blood. 93 (4): 1413–21. PMID 9949186.
- PDB: 1EVS; Deller MC, Hudson KR, Ikemizu S, Bravo J, Jones EY, Heath JK (August 2000). "Crystal structure and functional dissection of the cytostatic cytokine oncostatin M". Structure. 8 (8): 863–74. doi:10.1016/S0969-2126(00)00176-3. PMID 10997905.
- Rose TM, Lagrou MJ, Fransson I, Werelius B, Delattre O, Thomas G, de Jong PJ, Todaro GJ, Dumanski JP (July 1993). "The genes for oncostatin M (OSM) and leukemia inhibitory factor (LIF) are tightly linked on human chromosome 22". Genomics. 17 (1): 136–40. doi:10.1006/geno.1993.1294. PMID 8406444.
- Linsley PS, Bolton-Hanson M, Horn D, Malik N, Kallestad JC, Ochs V, Zarling JM, Shoyab M (March 1989). "Identification and characterization of cellular receptors for the growth regulator, oncostatin M". J. Biol. Chem. 264 (8): 4282–9. PMID 2538434.
- Gearing DP, Comeau MR, Friend DJ, Gimpel SD, Thut CJ, McGourty J, Brasher KK, King JA, Gillis S, Mosley B (March 1992). "The IL-6 signal transducer, gp130: an oncostatin M receptor and affinity converter for the LIF receptor". Science. 255 (5050): 1434–7. doi:10.1126/science.1542794. PMID 1542794.
- Thoma B, Bird TA, Friend DJ, Gearing DP, Dower SK (February 1994). "Oncostatin M and leukemia inhibitory factor trigger overlapping and different signals through partially shared receptor complexes". J. Biol. Chem. 269 (8): 6215–22. PMID 8119965.
- Mosley B, De Imus C, Friend D, Boiani N, Thoma B, Park LS, Cosman D (December 1996). "Dual oncostatin M (OSM) receptors. Cloning and characterization of an alternative signaling subunit conferring OSM-specific receptor activation". J. Biol. Chem. 271 (51): 32635–43. doi:10.1074/jbc.271.51.32635. PMID 8999038.
- Yoshimura A, Ichihara M, Kinjyo I, Moriyama M, Copeland NG, Gilbert DJ, Jenkins NA, Hara T, Miyajima A (March 1996). "Mouse oncostatin M: an immediate early gene induced by multiple cytokines through the JAK-STAT5 pathway". EMBO J. 15 (5): 1055–63. PMC 450003. PMID 8605875.
- Lindberg RA, Juan TS, Welcher AA, Sun Y, Cupples R, Guthrie B, Fletcher FA (June 1998). "Cloning and Characterization of a Specific Receptor for Mouse Oncostatin M". Mol. Cell. Biol. 18 (6): 3357–67. PMC 108917. PMID 9584176.
- Ichihara M, Hara T, Kim H, Murate T, Miyajima A (July 1997). "Oncostatin M and leukemia inhibitory factor do not use the same functional receptor in mice". Blood. 90 (1): 165–73. PMID 9207450.
- Stahl N, Boulton TG, Farruggella T, Ip NY, Davis S, Witthuhn BA, Quelle FW, Silvennoinen O, Barbieri G, Pellegrini S (January 1994). "Association and activation of Jak-Tyk kinases by CNTF-LIF-OSM-IL-6 beta receptor components". Science. 263 (5143): 92–5. doi:10.1126/science.8272873. PMID 8272873.
- Briscoe J, Rogers NC, Witthuhn BA, Watling D, Harpur AG, Wilks AF, Stark GR, Ihle JN, Kerr IM (February 1996). "Kinase-negative mutants of JAK1 can sustain interferon-gamma-inducible gene expression but not an antiviral state". EMBO J. 15 (4): 799–809. PMC 450278. PMID 8631301.
- Parganas E, Wang D, Stravopodis D, Topham DJ, Marine JC, Teglund S, Vanin EF, Bodner S, Colamonici OR, van Deursen JM, Grosveld G, Ihle JN (May 1998). "Jak2 is essential for signaling through a variety of cytokine receptors". Cell. 93 (3): 385–95. doi:10.1016/S0092-8674(00)81167-8. PMID 9590173.
- Taga T, Kishimoto T (1997). "Gp130 and the interleukin-6 family of cytokines". Annu. Rev. Immunol. 15: 797–819. doi:10.1146/annurev.immunol.15.1.797. PMID 9143707.
- Murakami M, Narazaki M, Hibi M, Yawata H, Yasukawa K, Hamaguchi M, Taga T, Kishimoto T (December 1991). "Critical cytoplasmic region of the interleukin 6 signal transducer gp130 is conserved in the cytokine receptor family". Proc. Natl. Acad. Sci. U.S.A. 88 (24): 11349–53. doi:10.1073/pnas.88.24.11349. PMC 53132. PMID 1662392.
- Tanner JW, Chen W, Young RL, Longmore GD, Shaw AS (March 1995). "The conserved box 1 motif of cytokine receptors is required for association with JAK kinases". J. Biol. Chem. 270 (12): 6523–30. doi:10.1074/jbc.270.12.6523. PMID 7896787.
- Narazaki M, Witthuhn BA, Yoshida K, Silvennoinen O, Yasukawa K, Ihle JN, Kishimoto T, Taga T (March 1994). "Activation of JAK2 kinase mediated by the interleukin 6 signal transducer gp130". Proc. Natl. Acad. Sci. U.S.A. 91 (6): 2285–9. doi:10.1073/pnas.91.6.2285. PMC 43355. PMID 8134389.
- Lai CF, Ripperger J, Morella KK, Wang Y, Gearing DP, Fey GH, Baumann H (June 1995). "Separate signaling mechanisms are involved in the control of STAT protein activation and gene regulation via the interleukin 6 response element by the box 3 motif of gp130". J. Biol. Chem. 270 (25): 14847–50. doi:10.1074/jbc.270.25.14847. PMID 7797460.
- Baumann H, Symes AJ, Comeau MR, Morella KK, Wang Y, Friend D, Ziegler SF, Fink JS, Gearing DP (January 1994). "Multiple regions within the cytoplasmic domains of the leukemia inhibitory factor receptor and gp130 cooperate in signal transduction in hepatic and neuronal cells". Mol. Cell. Biol. 14 (1): 138–46. PMC 358364. PMID 8264582.
- Heinrich PC, Behrmann I, Müller-Newen G, Schaper F, Graeve L (September 1998). "Interleukin-6-type cytokine signalling through the gp130/Jak/STAT pathway". Biochem. J. 334 (Pt 2): 297–314. doi:10.1042/bj3340297. PMC 1219691. PMID 9716487.
- Kuropatwinski KK, De Imus C, Gearing D, Baumann H, Mosley B (June 1997). "Influence of subunit combinations on signaling by receptors for oncostatin M, leukemia inhibitory factor, and interleukin-6". J. Biol. Chem. 272 (24): 15135–44. doi:10.1074/jbc.272.24.15135. PMID 9182534.
- Gerhartz C, Heesel B, Sasse J, Hemmann U, Landgraf C, Schneider-Mergener J, Horn F, Heinrich PC, Graeve L (May 1996). "Differential activation of acute phase response factor/STAT3 and STAT1 via the cytoplasmic domain of the interleukin 6 signal transducer gp130. I. Definition of a novel phosphotyrosine motif mediating STAT1 activation". J. Biol. Chem. 271 (22): 12991–8. doi:10.1074/jbc.271.22.12991. PMID 8662591.
- Ernst M, Novak U, Nicholson SE, Layton JE, Dunn AR (April 1999). "The carboxyl-terminal domains of gp130-related cytokine receptors are necessary for suppressing embryonic stem cell differentiation. Involvement of STAT3". J. Biol. Chem. 274 (14): 9729–37. doi:10.1074/jbc.274.14.9729. PMID 10092661.
- Starr R, Novak U, Willson TA, Inglese M, Murphy V, Alexander WS, Metcalf D, Nicola NA, Hilton DJ, Ernst M (August 1997). "Distinct roles for leukemia inhibitory factor receptor alpha-chain and gp130 in cell type-specific signal transduction". J. Biol. Chem. 272 (32): 19982–6. doi:10.1074/jbc.272.32.19982. PMID 9242667.
- Stahl N, Farruggella TJ, Boulton TG, Zhong Z, Darnell JE, Yancopoulos GD (March 1995). "Choice of STATs and other substrates specified by modular tyrosine-based motifs in cytokine receptors". Science. 267 (5202): 1349–53. doi:10.1126/science.7871433. PMID 7871433.
- Hermanns HM, Radtke S, Haan C, Schmitz-Van de Leur H, Tavernier J, Heinrich PC, Behrmann I (December 1999). "Contributions of leukemia inhibitory factor receptor and oncostatin M receptor to signal transduction in heterodimeric complexes with glycoprotein 130". J. Immunol. 163 (12): 6651–8. PMID 10586060.
- Brown TJ, Rowe JM, Liu JW, Shoyab M (October 1991). "Regulation of IL-6 expression by oncostatin M". J. Immunol. 147 (7): 2175–80. PMID 1918953.
- Yao L, Pan J, Setiadi H, Patel KD, McEver RP (July 1996). "Interleukin 4 or oncostatin M induces a prolonged increase in P- selectin mRNA and protein in human endothelial cells". J. Exp. Med. 184 (1): 81–92. doi:10.1084/jem.184.1.81. PMC 2192668. PMID 8691152.
- Wallace PM, MacMaster JF, Rouleau KA, Brown TJ, Loy JK, Donaldson KL, Wahl AF (May 1999). "Regulation of inflammatory responses by oncostatin M". J. Immunol. 162 (9): 5547–55. PMID 10228036.
- Nair BC, DeVico AL, Nakamura S, Copeland TD, Chen Y, Patel A, O'Neil T, Oroszlan S, Gallo RC, Sarngadharan MG (March 1992). "Identification of a major growth factor for AIDS-Kaposi's sarcoma cells as oncostatin M". Science. 255 (5050): 1430–2. doi:10.1126/science.1542792. PMID 1542792.
- Murakami-Mori K, Taga T, Kishimoto T, Nakamura S (September 1995). "AIDS-associated Kaposi's sarcoma (KS) cells express oncostatin M (OM)-specific receptor but not leukemia inhibitory factor/OM receptor or interleukin-6 receptor. Complete block of OM-induced KS cell growth and OM binding by anti-gp130 antibodies". J. Clin. Invest. 96 (3): 1319–27. doi:10.1172/JCI118167. PMC 185754. PMID 7657807.
- Heinrich PC, Horn F, Graeve L, Dittrich E, Kerr I, Müller-Newen G, Grötzinger J, Wollmer A (1998). "Interleukin-6 and related cytokines: effect on the acute phase reaction". Z Ernahrungswiss. 37 Suppl 1: 43–9. PMID 9558728.
Further reading
- Schieven GL, Kallestad JC, Brown TJ, Ledbetter JA, Linsley PS (September 1992). "Oncostatin M induces tyrosine phosphorylation in endothelial cells and activation of p62yes tyrosine kinase". J. Immunol. 149 (5): 1676–82. PMID 1324279.
- Hermanns HM, Radtke S, Schaper F, Heinrich PC, Behrmann I (December 2000). "Non-redundant signal transduction of interleukin-6-type cytokines. The adapter protein Shc is specifically recruited to the oncostatin M receptor". J. Biol. Chem. 275 (52): 40742–8. doi:10.1074/jbc.M005408200. PMID 11016927.
External links
- Oncostatin+M at the US National Library of Medicine Medical Subject Headings (MeSH)
PDB gallery | |
|---|---|
|