Chemokine
Chemokines (Greek -kinos, movement) are a family of small cytokines, or signaling proteins secreted by cells. Their name is derived from their ability to induce directed chemotaxis in nearby responsive cells; they are chemotactic cytokines.
| Small cytokines (intecrine/chemokine), interleukin-8 like | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
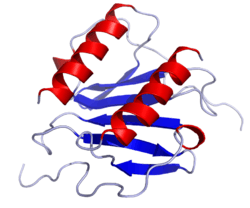 Solution structure of interleukin-8, a chemokine of the CXC subfamily | |||||||||||
| Identifiers | |||||||||||
| Symbol | IL8 | ||||||||||
| Pfam | PF00048 | ||||||||||
| InterPro | IPR001811 | ||||||||||
| PROSITE | PDOC00434 | ||||||||||
| SCOPe | 3il8 / SUPFAM | ||||||||||
| |||||||||||
Cytokine proteins are classified as chemokines according to behavior and structural characteristics. In addition to being known for mediating chemotaxis, chemokines are all approximately 8-10 kilodaltons in mass and have four cysteine residues in conserved locations that are key to forming their 3-dimensional shape.
These proteins have historically been known under several other names including the SIS family of cytokines, SIG family of cytokines, SCY family of cytokines, Platelet factor-4 superfamily or intercrines. Some chemokines are considered pro-inflammatory and can be induced during an immune response to recruit cells of the immune system to a site of infection, while others are considered homeostatic and are involved in controlling the migration of cells during normal processes of tissue maintenance or development. Chemokines are found in all vertebrates, some viruses and some bacteria, but none have been found in other invertebrates.
Chemokines have been classified into four main subfamilies: CXC, CC, CX3C and XC. All of these proteins exert their biological effects by interacting with G protein-linked transmembrane receptors called chemokine receptors, that are selectively found on the surfaces of their target cells.[1]
Function
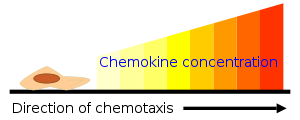
The major role of chemokines is to act as a chemoattractant to guide the migration of cells. Cells that are attracted by chemokines follow a signal of increasing chemokine concentration towards the source of the chemokine. Some chemokines control cells of the immune system during processes of immune surveillance, such as directing lymphocytes to the lymph nodes so they can screen for invasion of pathogens by interacting with antigen-presenting cells residing in these tissues. These are known as homeostatic chemokines and are produced and secreted without any need to stimulate their source cells. Some chemokines have roles in development; they promote angiogenesis (the growth of new blood vessels), or guide cells to tissues that provide specific signals critical for cellular maturation. Other chemokines are inflammatory and are released from a wide variety of cells in response to bacterial infection, viruses and agents that cause physical damage such as silica or the urate crystals that occur in gout. Their release is often stimulated by pro-inflammatory cytokines such as interleukin 1. Inflammatory chemokines function mainly as chemoattractants for leukocytes, recruiting monocytes, neutrophils and other effector cells from the blood to sites of infection or tissue damage. Certain inflammatory chemokines activate cells to initiate an immune response or promote wound healing. They are released by many different cell types and serve to guide cells of both innate immune system and adaptive immune system.
Types by function
Chemokines are functionally divided into two groups:[2]
- Homeostatic: are constitutively produced in certain tissues and are responsible for basal leukocyte migration. These include: CCL14, CCL19, CCL20, CCL21, CCL25, CCL27, CXCL12 and CXCL13. This classification is not strict; for example, CCL20 can act also as pro-inflammatory chemokine.[2]
- Inflammatory: these are formed under pathological conditions (on pro-inflammatory stimuli, such as IL-1, TNF-alpha, LPS, or viruses) and actively participate in the inflammatory response attracting immune cells to the site of inflammation. Examples are: CXCL-8, CCL2, CCL3, CCL4, CCL5, CCL11, CXCL10.[3]
Homing
The main function of chemokines is to manage the migration of leukocytes (homing) in the respective anatomical locations in inflammatory and homeostatic processes.
Basal: homeostatic chemokines are basal produced in the thymus and lymphoid tissues. Their homeostatic function in homing is best exemplified by the chemokines CCL19 and CCL21 (expressed within lymph nodes and on lymphatic endothelial cells) and their receptor CCR7 (expressed on cells destined for homing in cells to these organs). Using these ligands is possible routing antigen-presenting cells (APC) to lymph nodes during the adaptive immune response. Among other homeostatic chemokine receptors include: CCR9, CCR10, and CXCR5, which are important as part of the cell addresses for tissue-specific homing of leukocytes. CCR9 supports the migration of leukocytes into the intestine, CCR10 to the skin and CXCR5 supports the migration of B-cell to follicles of lymph nodes. As well CXCL12 (SDF-1) constitutively produced in the bone marrow promotes proliferation of progenitor B cells in the bone marrow microenvironment.[4][5]
Inflammatory: inflammatory chemokines are produced in high concentrations during infection or injury and determine the migration of inflammatory leukocytes into the damaged area. Typical inflammatory chemokines include: CCL2, CCL3 and CCL5, CXCL1, CXCL2 and CXCL8. A typical example is CXCL-8, which acts as a chemoattractant for neutrophils. In contrast to the homeostatic chemokine receptors, there is significant promiscuity (redundancy) associated with binding receptor and inflammatory chemokines. This often complicates research on receptor-specific therapeutics in this area.[5]
Types by cell attracted
- Monocytes / macrophages: the key chemokines that attract these cells to the site of inflammation include: CCL2, CCL3, CCL5, CCL7, CCL8, CCL13, CCL17 and CCL22.
- T-lymphocytes: the four key chemokines that are involved in the recruitment of T lymphocytes to the site of inflammation are: CCL2, CCL1, CCL22 and CCL17. Furthermore, CXCR3 expression by T-cells is induced following T-cell activation and activated T-cells are attracted to sites of inflammation where the IFN-y inducible chemokines CXCL9, CXCL10 and CXCL11 are secreted.[6]
- Mast cells: on their surface express several receptors for chemokines: CCR1, CCR2, CCR3, CCR4, CCR5, CXCR2, and CXCR4. Ligands of these receptors CCL2 and CCL5 play an important role in mast cell recruitment and activation in the lung. There is also evidence that CXCL8 might be inhibitory of mast cells.
- Eosinophils: the migration of eosinophils into various tissues involved several chemokines of CC family: CCL11, CCL24, CCL26, CCL5, CCL7, CCL13, and CCL3. Chemokines CCL11 (eotaxin) and CCL5 (RANTES) acts through a specific receptor CCR3 on the surface of eosinophils, and eotaxin plays an essential role in the initial recruitment of eosinophils into the lesion.
- Neutrophils: are regulated primarily by CXC chemokines. An example CXCL8 (IL-8) is chemoattractant for neutrophils and also activating their metabolic and degranulation.[7]
Structural characteristics
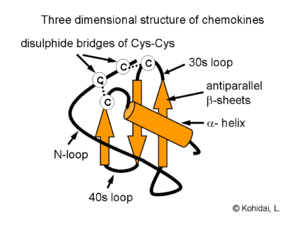
Proteins are classified into the chemokine family based on their structural characteristics, not just their ability to attract cells. All chemokines are small, with a molecular mass of between 8 and 10 kDa. They are approximately 20-50% identical to each other; that is, they share gene sequence and amino acid sequence homology. They all also possess conserved amino acids that are important for creating their 3-dimensional or tertiary structure, such as (in most cases) four cysteines that interact with each other in pairs to create a Greek key shape that is a characteristic of chemokines. Intramolecular disulfide bonds typically join the first to third, and the second to fourth cysteine residues, numbered as they appear in the protein sequence of the chemokine. Typical chemokine proteins are produced as pro-peptides, beginning with a signal peptide of approximately 20 amino acids that gets cleaved from the active (mature) portion of the molecule during the process of its secretion from the cell. The first two cysteines, in a chemokine, are situated close together near the N-terminal end of the mature protein, with the third cysteine residing in the centre of the molecule and the fourth close to the C-terminal end. A loop of approximately ten amino acids follows the first two cysteines and is known as the N-loop. This is followed by a single-turn helix, called a 310-helix, three β-strands and a C-terminal α-helix. These helices and strands are connected by turns called 30s, 40s and 50s loops; the third and fourth cysteines are located in the 30s and 50s loops.[8]
Types by structure
 | ||||
| CC chemokines | ||||
|---|---|---|---|---|
| Name | Gene | Other name(s) | Receptor | Uniprot |
| CCL1 | Scya1 | I-309, TCA-3 | CCR8 | |
| CCL2 | Scya2 | MCP-1 | CCR2 | P13500 |
| CCL3 | Scya3 | MIP-1a | CCR1 | P10147 |
| CCL4 | Scya4 | MIP-1β | CCR1, CCR5 | P13236 |
| CCL5 | Scya5 | RANTES | CCR5 | P13501 |
| CCL6 | Scya6 | C10, MRP-2 | CCR1 | P27784 |
| CCL7 | Scya7 | MARC, MCP-3 | CCR2 | P80098 |
| CCL8 | Scya8 | MCP-2 | CCR1, CCR2, CCR5 | P80075 |
| CCL9/CCL10 | Scya9 | MRP-2, CCF18, MIP-1? | CCR1 | P51670 |
| CCL11 | Scya11 | Eotaxin | CCR2, CCR3, CCR5 | P51671 |
| CCL12 | Scya12 | MCP-5 | Q62401 | |
| CCL13 | Scya13 | MCP-4, NCC-1, Ckβ10 | CCR2, CCR3, CCR5 | Q99616 |
| CCL14 | Scya14 | HCC-1, MCIF, Ckβ1, NCC-2, CCL | CCR1 | Q16627 |
| CCL15 | Scya15 | Leukotactin-1, MIP-5, HCC-2, NCC-3 | CCR1, CCR3 | Q16663 |
| CCL16 | Scya16 | LEC, NCC-4, LMC, Ckβ12 | CCR1, CCR2, CCR5, CCR8 | O15467 |
| CCL17 | Scya17 | TARC, dendrokine, ABCD-2 | CCR4 | Q92583 |
| CCL18 | Scya18 | PARC, DC-CK1, AMAC-1, Ckβ7, MIP-4 | P55774 | |
| CCL19 | Scya19 | ELC, Exodus-3, Ckβ11 | CCR7 | Q99731 |
| CCL20 | Scya20 | LARC, Exodus-1, Ckβ4 | CCR6 | P78556 |
| CCL21 | Scya21 | SLC, 6Ckine, Exodus-2, Ckβ9, TCA-4 | CCR7 | O00585 |
| CCL22 | Scya22 | MDC, DC/β-CK | CCR4 | O00626 |
| CCL23 | Scya23 | MPIF-1, Ckβ8, MIP-3, MPIF-1 | CCR1 | P55773 |
| CCL24 | Scya24 | Eotaxin-2, MPIF-2, Ckβ6 | CCR3 | O00175 |
| CCL25 | Scya25 | TECK, Ckβ15 | CCR9 | O15444 |
| CCL26 | Scya26 | Eotaxin-3, MIP-4a, IMAC, TSC-1 | CCR3 | Q9Y258 |
| CCL27 | Scya27 | CTACK, ILC, Eskine, PESKY, skinkine | CCR10 | Q9Y4X3 |
| CCL28 | Scya28 | MEC | CCR3, CCR10 | Q9NRJ3 |
| CXC chemokines | ||||
| Name | Gene | Other name(s) | Receptor | Uniprot |
| CXCL1 | Scyb1 | Gro-a, GRO1, NAP-3, KC | CXCR2 | P09341 |
| CXCL2 | Scyb2 | Gro-β, GRO2, MIP-2a | CXCR2 | P19875 |
| CXCL3 | Scyb3 | Gro-?, GRO3, MIP-2β | CXCR2 | P19876 |
| CXCL4 | Scyb4 | PF-4 | CXCR3B | P02776 |
| CXCL5 | Scyb5 | ENA-78 | CXCR2 | P42830 |
| CXCL6 | Scyb6 | GCP-2 | CXCR1, CXCR2 | P80162 |
| CXCL7 | Scyb7 | NAP-2, CTAPIII, β-Ta, PEP | P02775 | |
| CXCL8 | Scyb8 | IL-8, NAP-1, MDNCF, GCP-1 | CXCR1, CXCR2 | P10145 |
| CXCL9 | Scyb9 | MIG, CRG-10 | CXCR3 | Q07325 |
| CXCL10 | Scyb10 | IP-10, CRG-2 | CXCR3 | P02778 |
| CXCL11 | Scyb11 | I-TAC, β-R1, IP-9 | CXCR3, CXCR7 | O14625 |
| CXCL12 | Scyb12 | SDF-1, PBSF | CXCR4, CXCR7 | P48061 |
| CXCL13 | Scyb13 | BCA-1, BLC | CXCR5 | O43927 |
| CXCL14 | Scyb14 | BRAK, bolekine | O95715 | |
| CXCL15 | Scyb15 | Lungkine, WECHE | Q9WVL7 | |
| CXCL16 | Scyb16 | SRPSOX | CXCR6 | Q9H2A7 |
| CXCL17 | VCC-1 | DMC, VCC-1 | Q6UXB2 | |
| C chemokines | ||||
| Name | Gene | Other name(s) | Receptor | Uniprot |
| XCL1 | Scyc1 | Lymphotactin a, SCM-1a, ATAC | XCR1 | P47992 |
| XCL2 | Scyc2 | Lymphotactin β, SCM-1β | XCR1 | Q9UBD3 |
| CX3C chemokines | ||||
| Name | Gene | Other name(s) | Receptor | Uniprot |
| CX3CL1 | Scyd1 | Fractalkine, Neurotactin, ABCD-3 | CX3CR1 | P78423 |
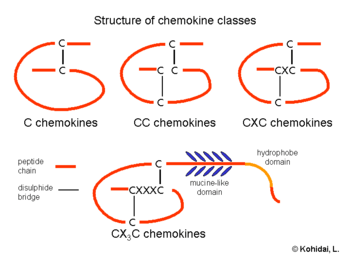
Members of the chemokine family are divided into four groups depending on the spacing of their first two cysteine residues. Thus the nomenclature for chemokines is, e.g.: CCL1 for the ligand 1 of the CC-family of chemokines, and CCR1 for its respective receptor.
CC chemokines
The CC chemokine (or β-chemokine) proteins have two adjacent cysteines (amino acids), near their amino terminus. There have been at least 27 distinct members of this subgroup reported for mammals, called CC chemokine ligands (CCL)-1 to -28; CCL10 is the same as CCL9. Chemokines of this subfamily usually contain four cysteines (C4-CC chemokines), but a small number of CC chemokines possess six cysteines (C6-CC chemokines). C6-CC chemokines include CCL1, CCL15, CCL21, CCL23 and CCL28.[9] CC chemokines induce the migration of monocytes and other cell types such as NK cells and dendritic cells.
Examples of CC chemokine include monocyte chemoattractant protein-1 (MCP-1 or CCL2) which induces monocytes to leave the bloodstream and enter the surrounding tissue to become tissue macrophages.
CCL5 (or RANTES) attracts cells such as T cells, eosinophils and basophils that express the receptor CCR5.
Increased CCL11 levels in blood plasma are associated with aging (and reduced neurogenesis) in mice and humans.[10]
CXC chemokines
The two N-terminal cysteines of CXC chemokines (or α-chemokines) are separated by one amino acid, represented in this name with an "X". There have been 17 different CXC chemokines described in mammals, that are subdivided into two categories, those with a specific amino acid sequence (or motif) of glutamic acid-leucine-arginine (or ELR for short) immediately before the first cysteine of the CXC motif (ELR-positive), and those without an ELR motif (ELR-negative). ELR-positive CXC chemokines specifically induce the migration of neutrophils, and interact with chemokine receptors CXCR1 and CXCR2. An example of an ELR-positive CXC chemokine is interleukin-8 (IL-8), which induces neutrophils to leave the bloodstream and enter into the surrounding tissue. Other CXC chemokines that lack the ELR motif, such as CXCL13, tend to be chemoattractant for lymphocytes. CXC chemokines bind to CXC chemokine receptors, of which seven have been discovered to date, designated CXCR1-7.
C chemokines
The third group of chemokines is known as the C chemokines (or γ chemokines), and is unlike all other chemokines in that it has only two cysteines; one N-terminal cysteine and one cysteine downstream. Two chemokines have been described for this subgroup and are called XCL1 (lymphotactin-α) and XCL2 (lymphotactin-β).
CX3C chemokines
A fourth group has also been discovered and members have three amino acids between the two cysteines and is termed CX3C chemokine (or d-chemokines). The only CX3C chemokine discovered to date is called fractalkine (or CX3CL1). It is both secreted and tethered to the surface of the cell that expresses it, thereby serving as both a chemoattractant and as an adhesion molecule.
Receptors
Chemokine receptors are G protein-coupled receptors containing 7 transmembrane domains that are found on the surface of leukocytes. Approximately 19 different chemokine receptors have been characterized to date, which are divided into four families depending on the type of chemokine they bind; CXCR that bind CXC chemokines, CCR that bind CC chemokines, CX3CR1 that binds the sole CX3C chemokine (CX3CL1), and XCR1 that binds the two XC chemokines (XCL1 and XCL2). They share many structural features; they are similar in size (with about 350 amino acids), have a short, acidic N-terminal end, seven helical transmembrane domains with three intracellular and three extracellular hydrophilic loops, and an intracellular C-terminus containing serine and threonine residues important for receptor regulation. The first two extracellular loops of chemokine receptors each has a conserved cysteine residue that allow formation of a disulfide bridge between these loops. G proteins are coupled to the C-terminal end of the chemokine receptor to allow intracellular signaling after receptor activation, while the N-terminal domain of the chemokine receptor determines ligand binding specificity.[11]
Signal transduction
Chemokine receptors associate with G-proteins to transmit cell signals following ligand binding. Activation of G proteins, by chemokine receptors, causes the subsequent activation of an enzyme known as phospholipase C (PLC). PLC cleaves a molecule called phosphatidylinositol (4,5)-bisphosphate (PIP2) into two second messenger molecules known as Inositol triphosphate (IP3) and diacylglycerol (DAG) that trigger intracellular signaling events; DAG activates another enzyme called protein kinase C (PKC), and IP3 triggers the release of calcium from intracellular stores. These events promote many signaling cascades (such as the MAP kinase pathway) that generate responses like chemotaxis, degranulation, release of superoxide anions and changes in the avidity of cell adhesion molecules called integrins within the cell harbouring the chemokine receptor.[11]
Infection control
The discovery that the β chemokines RANTES, MIP (macrophage inflammatory proteins) 1α and 1β (now known as CCL5, CCL3 and CCL4 respectively) suppress HIV-1 provided the initial connection and indicated that these molecules might control infection as part of immune responses in vivo,[12] and that sustained delivery of such inhibitors have the capacity of long-term infection control.[13] The association of chemokine production with antigen-induced proliferative responses, more favorable clinical status in HIV infection, as well as with an uninfected status in subjects at risk for infection suggests a positive role for these molecules in controlling the natural course of HIV infection.[14]
See also
- Paracrine signalling
References
- Mélik-Parsadaniantz S, Rostène W (July 2008). "Chemokines and neuromodulation". Journal of Neuroimmunology. 198 (1–2): 62–8. doi:10.1016/j.jneuroim.2008.04.022. PMID 18538863.
- Zlotnik A, Burkhardt AM, Homey B (August 2011). "Homeostatic chemokine receptors and organ-specific metastasis". Nature Reviews. Immunology. 11 (9): 597–606. doi:10.1038/nri3049. PMID 21866172.
- Zlotnik A, Yoshie O (May 2012). "The chemokine superfamily revisited". Immunity. 36 (5): 705–16. doi:10.1016/j.immuni.2012.05.008. PMC 3396424. PMID 22633458.
- Le Y, Zhou Y, Iribarren P, Wang J (April 2004). "Chemokines and chemokine receptors: their manifold roles in homeostasis and disease" (PDF). Cellular & Molecular Immunology. 1 (2): 95–104. PMID 16212895.
- Graham GJ, Locati M (January 2013). "Regulation of the immune and inflammatory responses by the 'atypical' chemokine receptor D6". The Journal of Pathology. 229 (2): 168–75. doi:10.1002/path.4123. PMID 23125030.
- Xie JH, Nomura N, Lu M, Chen SL, Koch GE, Weng Y, Rosa R, Di Salvo J, Mudgett J, Peterson LB, Wicker LS, DeMartino JA (June 2003). "Antibody-mediated blockade of the CXCR3 chemokine receptor results in diminished recruitment of T helper 1 cells into sites of inflammation". Journal of Leukocyte Biology. 73 (6): 771–80. doi:10.1189/jlb.1102573. PMID 12773510. S2CID 8175160.
- Ono SJ, Nakamura T, Miyazaki D, Ohbayashi M, Dawson M, Toda M (June 2003). "Chemokines: roles in leukocyte development, trafficking, and effector function". The Journal of Allergy and Clinical Immunology. 111 (6): 1185–99, quiz 1200. doi:10.1067/mai.2003.1594. PMID 12789214.
- Fernandez EJ, Lolis E (2002). "Structure, function, and inhibition of chemokines". Annual Review of Pharmacology and Toxicology. 42: 469–99. doi:10.1146/annurev.pharmtox.42.091901.115838. PMID 11807180.
- Laing KJ, Secombes CJ (May 2004). "Chemokines". Developmental and Comparative Immunology. 28 (5): 443–60. doi:10.1016/j.dci.2003.09.006. PMID 15062643.
- Villeda SA, Luo J, Mosher KI, Zou B, Britschgi M, Bieri G, Stan TM, Fainberg N, Ding Z, Eggel A, Lucin KM, Czirr E, Park JS, Couillard-Després S, Aigner L, Li G, Peskind ER, Kaye JA, Quinn JF, Galasko DR, Xie XS, Rando TA, Wyss-Coray T (August 2011). "The ageing systemic milieu negatively regulates neurogenesis and cognitive function". Nature. 477 (7362): 90–4. Bibcode:2011Natur.477...90V. doi:10.1038/nature10357. PMC 3170097. PMID 21886162.
- Murdoch C, Finn A (May 2000). "Chemokine receptors and their role in inflammation and infectious diseases". Blood. 95 (10): 3032–43. doi:10.1182/blood.V95.10.3032.010k17_3032_3043. PMID 10807766. Archived from the original on 2013-07-04.
- Cocchi F, DeVico AL, Garzino-Demo A, Arya SK, Gallo RC, Lusso P (December 1995). "Identification of RANTES, MIP-1 alpha, and MIP-1 beta as the major HIV-suppressive factors produced by CD8+ T cells". Science. 270 (5243): 1811–5. Bibcode:1995Sci...270.1811C. doi:10.1126/science.270.5243.1811. PMID 8525373.
- von Recum HA, Pokorski JK (May 2013). "Peptide and protein-based inhibitors of HIV-1 co-receptors". Experimental Biology and Medicine. 238 (5): 442–9. doi:10.1177/1535370213480696. PMC 3908444. PMID 23856897.
- Garzino-Demo A, Moss RB, Margolick JB, Cleghorn F, Sill A, Blattner WA, Cocchi F, Carlo DJ, DeVico AL, Gallo RC (October 1999). "Spontaneous and antigen-induced production of HIV-inhibitory beta-chemokines are associated with AIDS-free status". Proceedings of the National Academy of Sciences of the United States of America. 96 (21): 11986–91. Bibcode:1999PNAS...9611986G. doi:10.1073/pnas.96.21.11986. JSTOR 48922. PMC 18399. PMID 10518563.
External links
- The cytokine family database - Chemokines at kumamoto-u.ac.jp
- The correct chemokine nomenclature at rndsystems.com