Albatross
Albatrosses are very large seabirds in the family Diomedeidae. Albatrosses, They range widely in the Southern Ocean and the North Pacific. They are absent from the North Atlantic, although fossil remains show they once occurred there and occasional vagrants are found. Albatrosses are among the largest of flying birds, and species of the genus Diomedea (great albatrosses) have the longest wingspans of any extant birds, reaching up to 3.7 m (12 ft). The albatrosses are usually regarded as falling into four genera, but disagreement exists over the number of species.
| Albatross | |
|---|---|
 | |
| Short-tailed albatross (Phoebastria albatrus) | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Aves |
| Order: | Procellariiformes |
| Family: | Diomedeidae G.R. Gray 1840[1] |
| Genera | |
|
Diomedea | |
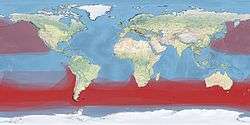 | |
| Global range density (in red) | |
Albatrosses are highly efficient in the air, using dynamic soaring and slope soaring to cover great distances with little exertion. It is the only bird that can fly 10,000 miles without landing. They feed on squid, fish, and krill by either scavenging, surface seizing, or diving. Albatrosses are colonial, nesting for the most part on remote oceanic islands, often with several species nesting together. Pair bonds between males and females form over several years, with the use of "ritualised dances", and last for the life of the pair. A breeding season can take over a year from laying to fledging, with a single egg laid in each breeding attempt. A Laysan albatross, named Wisdom, on Midway Island is recognised as the oldest wild bird in the world; she was first banded in 1956 by Chandler Robbins.
Of the 22 species of albatrosses recognised by the IUCN, all are listed as at some level of concern; three species are critically endangered, five species are endangered, seven species are near threatened, and seven species are vulnerable. Numbers of albatrosses have declined in the past due to harvesting for feathers. Albatrosses are threatened by introduced species, such as rats and feral cats that attack eggs, chicks, and nesting adults; by pollution; by a serious decline in fish stocks in many regions largely due to overfishing; and by longline fishing. Longline fisheries pose the greatest threat, as feeding birds are attracted to the bait, become hooked on the lines, and drown. Identified stakeholders such as governments, conservation organisations, and people in the fishing industry are all working toward reducing this bycatch.
Science
Taxonomy and evolution
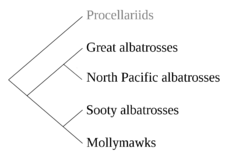
The "albatross" designation comprises between 13 and 24 species (the number is still a matter of some debate, 21 being the most commonly accepted number) in four genera. These genera are the great albatrosses (Diomedea), the mollymawks (Thalassarche), the North Pacific albatrosses (Phoebastria), and the sooty albatrosses or sooties (Phoebetria). The North Pacific albatrosses are considered to be a sister taxon to the great albatrosses, while the sooty albatrosses are considered closer to the mollymawks.[2]
The taxonomy of the albatross group has been a source of much debate. The Sibley-Ahlquist taxonomy places seabirds, birds of prey, and many others in a greatly enlarged order, the Ciconiiformes, whereas the ornithological organisations in North America, Europe, South Africa, Australia, and New Zealand retain the more traditional order Procellariiformes. The albatrosses can be separated from the other Procellariiformes both genetically and through morphological characteristics, size, their legs, and the arrangement of their nasal tubes (see below: Morphology and flight).[2]
Within the family, the assignment of genera has been debated for over 100 years. Originally placed into a single genus, Diomedea, they were rearranged by Reichenbach into four different genera in 1852, then lumped back together and split apart again several times, acquiring 12 different genus names in total (though never more than eight at one time) by 1965 (Diomedea, Phoebastria, Thalassarche, Phoebetria, Thalassageron, Diomedella, Nealbatrus, Rhothonia, Julietata, Galapagornis, Laysanornis, and Penthirenia).[3]

By 1965, in an attempt to bring some order back to the classification of albatrosses, they were lumped into two genera, Phoebetria (the sooty albatrosses which most closely seemed to resemble the procellarids and were at the time considered "primitive" ) and Diomedea (the rest).[4] Though a case was made for the simplification of the family (particularly the nomenclature), the classification was based on the morphological analysis by Elliott Coues in 1866, and paid little attention to more recent studies and even ignored some of Coues's suggestions.[3]
More recent research by Gary Nunn of the American Museum of Natural History (1996) and other researchers around the world studied the mitochondrial DNA of all 14 accepted species, finding four, not two, monophyletic groups within the albatrosses.[5] They proposed the resurrection of two of the old genus names, Phoebastria for the North Pacific albatrosses and Thalassarche for the mollymawks, with the great albatrosses retaining Diomedea and the sooty albatrosses staying in Phoebetria. Both the British Ornithologists' Union and the South African authorities split the albatrosses into four genera as Nunn suggested, and the change has been accepted by the majority of researchers.
While some agree on the number of genera, fewer agree on the number of species. Historically, up to 80 different taxa have been described by different researchers; most of these were incorrectly identified juvenile birds.[6]
Based on the work on albatross genera, Robertson and Nunn went on in 1998 to propose a revised taxonomy with 24 different species,[3] compared to the 14 then accepted. This expanded taxonomy elevated many established subspecies to full species, but was criticised for not using, in every case, peer reviewed information to justify the splits. Since then, further studies have in some instances supported or disproved the splits; a 2004 paper analysing the mitochondrial DNA and microsatellites agreed with the conclusion that the Antipodean albatross and the Tristan albatross were distinct from the wandering albatross, per Robertson and Nunn, but found that the suggested Gibson's albatross, Diomedea gibsoni, was not distinct from the Antipodean albatross.[7] For the most part, an interim taxonomy of 21 species is accepted by ITIS and many other researchers, though by no means all—in 2004 Penhallurick and Wink called for the number of species to be reduced to 13 (including the lumping of the Amsterdam albatross with the wandering albatross),[8] although this paper was itself controversial.[6][9] On all sides is the widespread agreement on the need for further research to clarify the issue.
Sibley and Ahlquist's molecular study of the evolution of the bird families has put the radiation of the Procellariiformes in the Oligocene period 35–30 million years ago (Mya), though this group probably originated earlier, with a fossil sometimes attributed to the order, a seabird known as Tytthostonyx, being found in late Cretaceous rocks (70 Mya). The molecular evidence suggests that the storm petrels were the first to diverge from the ancestral stock, and the albatrosses next, with the procellarids and diving petrels separating later. The earliest fossil albatrosses were found in Eocene to Oligocene rocks, although some of these are only tentatively assigned to the family and none appear to be particularly close to the living forms. They are Murunkus (Middle Eocene of Uzbekistan), Manu (early Oligocene of New Zealand), and an undescribed form from the Late Oligocene of South Carolina. The oldest widely accepted fossil albatross is Tydea septentrionalis from the early Oligocene of Belgium.[10] Diomedavus knapptonensis is smaller than all extant albatrosses and was found in late Oligocene strata of Washington State, USA.[11] Plotornis was formerly often considered a petrel but is now accepted as an albatross. It is from the Middle Miocene of France, a time when the split between the four modern genera was already underway as evidenced by Phoebastria californica and Diomedea milleri, both being mid-Miocene species from Sharktooth Hill, California. These show that the split between the great albatrosses and the North Pacific albatrosses occurred by 15 Mya. Similar fossil finds in the Southern Hemisphere put the split between the sooties and mollymawks at 10 Mya.[2] The fossil record of the albatrosses in the Northern Hemisphere is more complete than that of the Southern, and many fossil forms of albatross have been found in the North Atlantic, which today has no albatrosses. The remains of a colony of short-tailed albatrosses have been uncovered on the island of Bermuda,[12] and the majority of fossil albatrosses from the North Atlantic have been of the genus Phoebastria (the North Pacific albatrosses); one, Phoebastria anglica, has been found in deposits in both North Carolina and England. Due to convergent evolution in particular of the leg and foot bones, remains of the prehistoric pseudotooth birds (Pelagornithidae) may be mistaken for those of extinct albatrosses; Manu may be such a case, and quite certainly the supposed giant albatross femur from the Early Pleistocene[13] Dainichi Formation at Kakegawa, Japan, actually is from one of the last pseudotooth birds. Aldiomedes angustirostris was a uniquely narrow-beaked species from the Pliocene of New Zealand.[10] The articles on Diomedea, Phoebastria, and Thalassarche contain more data on fossil species of living albatross genera.
Morphology and flight

The albatrosses are a group of large to very large birds; they are the largest of the Procellariiformes. The bill is large, strong, and sharp-edged, with the upper mandible terminating in a large hook. This bill is composed of several horny plates, and along the sides are the two "tubes", long nostrils that give the order its former name. The tubes of all albatrosses are along the sides of the bill, unlike the rest of the Procellariiformes, where the tubes run along the top of the bill. These tubes allow the albatrosses to measure the exact airspeed in flight; the nostrils are analogous to the pitot tubes in modern aircraft. The albatross needs accurate airspeed measurement in order to perform dynamic soaring. Like other Procellariiformes, they use their uniquely developed sense of smell to locate potential food sources, whereas most birds depend on eyesight.[14] The feet have no hind toe and the three anterior toes are completely webbed. The legs are strong for Procellariiformes, making them and the giant petrels the only members of that order that can walk well on land.[15]
Albatrosses, along with all Procellariiformes, must excrete the salts they ingest in drinking sea water and eating marine invertebrates. All birds have an enlarged nasal gland at the base of the bill, above their eyes. This gland is inactive in species that do not require it, but in the Procellariiformes, it acts as a salt gland. Scientists are uncertain as to its exact processes, but do know in general terms that it removes salt by secreting a 5% saline solution that drips out of their noses or is forcibly ejected.[16]

The adult plumage of most of the albatrosses is usually some variation of dark upper-wing and back with white undersides, often compared to that of a gull.[15] The extent of colouration varies: the southern royal albatross is almost completely white except for the ends and trailing edges of the wings in fully mature males, while the Amsterdam albatross has an almost juvenile-like breeding plumage with a great deal of brown, particularly a strong brown band around the chest. Several species of mollymawks and North Pacific albatrosses have face markings like eye patches or have grey or yellow on the head and nape. Three albatross species, the black-footed albatross and the two sooty albatrosses, vary completely from the usual patterns and are almost entirely dark brown (or dark grey in places in the case of the light-mantled albatross). Albatrosses take several years to get their full adult breeding plumage.[2]
The wingspans of the largest great albatrosses (genus Diomedea) are the largest of any bird, exceeding 340 cm (11.2 ft), although the other species' wingspans are considerably smaller at, down to 1.75 m (5.7 ft).[17] The wings are stiff and cambered, with thickened streamlined leading edges. Albatrosses travel huge distances with two techniques used by many long-winged seabirds: dynamic soaring and slope soaring. Dynamic soaring involves repeatedly rising into wind and descending downwind, thus gaining energy from the vertical wind gradient. The only effort expended is in the turns at the top and bottom of every such loop. This maneuver allows the bird to cover almost 1000 km/day without flapping its wings. Slope soaring uses the rising air on the windward side of large waves. Albatross have high glide ratios, around 22:1 to 23:1, meaning that for every metre they drop, they can travel forward 22 metres.[2] They are aided in soaring by a shoulder-lock, a sheet of tendon that locks the wing when fully extended, allowing the wing to be kept outstretched without any muscle expenditure, a morphological adaptation they share with the giant petrels.[18]

Albatrosses combine these soaring techniques with the use of predictable weather systems: albatrosses in the Southern Hemisphere flying north from their colonies take a clockwise route, and those flying south fly counterclockwise.[15] Albatrosses are so well adapted to this lifestyle that their heart rates while flying are close to their basal heart rate when resting. This efficiency is such that the most energetically demanding aspect of a foraging trip is not the distance covered, but the landings, take-offs and hunting they undertake having found a food source.[19] A common assumption is that albatrosses must be able to sleep in flight, although no direct evidence has ever been obtained.[20]
This efficient long-distance travelling underlies the albatross's success as a long-distance forager, covering great distances and expending little energy looking for patchily distributed food sources. Their adaptation to gliding flight makes them dependent on wind and waves, but their long wings are ill-suited to powered flight and most species lack the muscles and energy to undertake sustained flapping flight. Albatrosses in calm seas rest on the ocean's surface until the wind picks up again, as using powered flight is not energetically worthwhile though they are capable of flight to avoid danger.[21] The North Pacific albatrosses can use a flight style known as flap-gliding, where the bird progresses by bursts of flapping followed by gliding.[22] When taking off, albatrosses need to take a run up to allow enough air to move under the wing to provide lift.[15]
The dynamic soaring of albatrosses is inspiring to airplane designers: German aerospace engineer Johannes Traugott and colleagues have charted the albatross's nuanced flight pattern and are looking for ways to apply this to aircraft, especially in the area of drones and unmanned aircraft.[23]
Distribution and range at sea

Most albatrosses range in the Southern Hemisphere from Antarctica to Australia, South Africa and South America. The exceptions to this are the four North Pacific albatrosses, of which three occur exclusively in the North Pacific, from Hawaii to Japan, California, and Alaska; and one, the waved albatross, breeds in the Galápagos Islands and feeds off the coast of South America. The need for wind to enable gliding is the reason albatrosses are for the most part confined to higher latitudes: being unsuited to sustained flapping flight makes crossing the doldrums extremely difficult. The exception, the waved albatross, is able to live in the equatorial waters around the Galápagos Islands because of the cool waters of the Humboldt Current and the resulting winds.[2]
It is not known for certain why the albatrosses became extinct in the North Atlantic, although rising sea levels due to an interglacial warming period are thought to have submerged the site of a short-tailed albatross colony that has been excavated in Bermuda.[12] Some southern species have occasionally turned up as vagrants in the North Atlantic and can become exiled, remaining there for decades. One of these exiles, a black-browed albatross, returned to gannet colonies in Scotland for many years in an attempt to breed.[24]
The use of satellite tracking is teaching scientists a great deal about the way albatrosses range across the ocean to find food. They undertake no annual migration, but disperse widely after breeding—Southern Hemisphere species often undertake circumpolar trips.[25] Evidence also exists of separate ranges for different species at sea. A comparison of the foraging niches of two related species that breed on Campbell Island, the Campbell albatross and the grey-headed albatross, showed the Campbell albatross primarily fed over the Campbell Plateau, whereas the grey-headed albatross fed in more pelagic, oceanic waters. Wandering albatrosses also react strongly to bathymetry, feeding only in waters deeper than 1000 m (3281 ft); so rigidly did the satellite plots match this contour that one scientist remarked, "It almost appears as if the birds notice and obey a 'No Entry' sign where the water shallows to less than 1000 m".[2] Also, evidence shows different ranges for the two sexes of the same species; a study of Tristan albatrosses breeding on Gough Island showed that males foraged to the west of Gough and females to the east.[2]
Birdlife has satellite tracking records for each of the 22 species of albatross in their seabird-tracking database.
Diet

The albatross diet is predominantly cephalopods, fish, crustaceans, and offal (organ meat),[15] although they will also scavenge carrion and feed on other zooplankton.[15] For most species a comprehensive understanding of diet is known for only the breeding season, when the albatrosses regularly return to land and study is possible. The importance of each of these food sources varies from species to species, and even from population to population; some concentrate on squid alone, others take more krill or fish. Of the two albatross species found in Hawaii, one, the black-footed albatross, takes mostly fish while the Laysan feeds on squid.[15]
The use of data loggers at sea that record ingestion of water against time (providing a likely time of feeding) suggests that albatrosses predominantly feed during the day. Analysis of the squid beaks regurgitated by albatrosses has shown that many of the squid eaten are too large to have been caught alive, and include mid-water species likely to be beyond the reach of albatross, suggesting that, for some species (like the wandering albatross), scavenged squid may be an important part of the diet. The source of these dead squid is a matter of debate; some certainly comes from squid fisheries, but in nature it primarily comes from the die-off that occurs after squid spawning and the vomit of squid-eating whales (sperm whales, pilot whales and southern bottlenose whales).[26] The diet of other species, like the black-browed albatross or the grey-headed albatross, is rich with smaller species of squid that tend to sink after death, and scavenging is not assumed to play a large role in their diet.[2] The waved albatross has been observed practising kleptoparasitism, harassing boobies to steal their food, making it the only member of its order to do so regularly.[27]
Until recently, albatrosses were thought to be predominantly surface feeders, swimming at the surface and snapping up squid and fish pushed to the surface by currents, predators, or death. The deployment of capillary depth recorders, which record the maximum dive depth undertaken by a bird, has shown that while some species, like the wandering albatross, do not dive deeper than a metre, some species, like the light-mantled albatross, have a mean diving depth of almost 5 m and can dive as deep as 12.5 m.[28] In addition to surface feeding and diving, they have also been observed plunge diving from the air to snatch prey.[29]
Breeding and dancing
%E2%80%93Wandering_albatross_(Diomedea_exulans)_06.jpg)
Albatrosses are colonial, usually nesting on isolated islands; where colonies are on larger landmasses, they are found on exposed headlands with good approaches from the sea in several directions, like the colony on the Otago Peninsula in Dunedin, New Zealand. Many Buller's albatrosses and black-footed albatrosses nest under trees in open forest.[30] Colonies vary from the very dense aggregations favoured by the mollymawks (black-browed albatross colonies on the Falkland Islands have densities of 70 nests per 100 m2) to the much looser groups and widely spaced individual nests favoured by the sooty and great albatrosses. All albatross colonies are on islands that historically were free of land mammals. Albatrosses are highly philopatric, meaning they usually return to their natal colony to breed. This tendency is so strong that a study of Laysan albatrosses showed that the average distance between hatching site and the site where a bird established its own territory was 22 m (72 ft).[31]
Albatrosses live much longer than other birds; they delay breeding for longer and invest more effort into fewer young. Most species survive upwards of 50 years, the oldest recorded being a Laysan albatross named Wisdom that was ringed in 1956 as a mature adult and hatched another chick in February 2017, making her at least 66 years old. She is the oldest confirmed wild bird and the oldest banded bird in the world.[32][33]
Albatrosses reach sexual maturity slowly, after about five years, but even once they have reached maturity, they do not begin to breed for another few years (even up to 10 years for some species). Young nonbreeders attend a colony prior to beginning to breed, spending many years practising the elaborate breeding rituals and "dances" for which the family is famous.[34] Birds arriving back at the colony for the first time already have the stereotyped behaviours that compose albatross language, but can neither "read" that behaviour as exhibited by other birds nor respond appropriately.[15] After a period of trial and error learning, the young birds learn the syntax and perfect the dances. This language is mastered more rapidly if the younger birds are around older birds.
The repertoire of behaviour involves synchronised performances of various actions such as preening, pointing, calling, bill clacking, staring, and combinations of such behaviours (such as the sky-call).[35] When a bird first returns to the colony, it dances with many partners, but after a number of years, the number of interactions drops, until one partner is chosen and a pair is formed. They then continue to perfect an individual language that will eventually be unique to that one pair. Having established a pair bond that will last for life, however, most of that dance will never be used again.

Albatrosses are held to undertake these elaborate and painstaking rituals to ensure that the appropriate partner has been chosen and to perfect partner recognition, as egg laying and chick rearing is a huge investment. Even species that can complete an egg-laying cycle in under a year seldom lay eggs in consecutive years.[2] The great albatrosses (i.e. wandering albatross) take over a year to raise a chick from laying to fledging. Albatrosses lay a single subelliptical[17] egg, white with reddish-brown spots,[30] in a breeding season; if the egg is lost to predators or accidentally broken, then no further breeding attempts are made that year. The larger eggs weigh from 200 to 510 g (7.1 to 18.0 oz).[30] The "divorce" of a pair is a rare occurrence, due to the diminished lifetime reproductive success it causes, and usually happens only after several years of breeding failure.[2]
All the southern albatrosses create large nests for their egg, using grass, shrubs, soil, peat, and even penguin feathers,[30] whereas the three species in the North Pacific make more rudimentary nests. The waved albatross, though, makes no nest and even moves its egg around the pair's territory, as much as 50 m (160 ft), sometimes causing it to lose the egg.[36] In all albatross species, both parents incubate the egg in stints that last between one day and three weeks. Incubation lasts around 70 to 80 days (longer for the larger albatrosses), the longest incubation period of any bird. It can be an energetically demanding process, with the adult losing as much as 83 g (2.9 oz) of body weight a day.[37]
_01.jpg)
After hatching, the chick, which is semialtricial,[17] is brooded and guarded for three weeks until it is large enough to defend and thermoregulate itself. During this period, the parents feed the chick small meals when they relieve each other from duty. After the brooding period is over, the chick is fed in regular intervals by both parents. The parents adopt alternative patterns of short and long foraging trips, providing meals that weigh around 12% of their body weight (around 600 g (21 oz)). The meals are composed of fresh squid, fish, and krill, as well as stomach oil, an energy-rich food that is lighter to carry than undigested prey items. This oil is created in a stomach organ known as a proventriculus from digested prey items by most Procellariiformes, and gives them their distinctive musty smell.[38]
Albatross chicks take a long time to fledge. In the case of the great albatrosses, it can take up to 280 days; even for the smaller albatrosses, it takes between 140 and 170 days.[39] Like many seabirds, albatross chicks will gain enough weight to be heavier than their parents, and prior to fledging they use these reserves to build up body condition (particularly growing all their flight feathers), usually fledging at the same weight as their parents. Between 15% and 65% of those fledged survive to breed.[30] Albatross chicks fledge on their own and receive no further help from their parents, which return to the nest after fledging, unaware their chick has left. Studies of juveniles dispersing at sea have suggested an innate migration behaviour, a genetically coded navigation route, which helps young birds when they are first out at sea.[40]
Hybridization is rare in albatrosses, largely due to the low incidence of breeding-site vagrancy.[41]
Albatrosses and humans
Etymology
The name "albatross" is derived from the Arabic al-qādūs or al-ḡaṭṭās (a pelican; literally, "the diver"), which travelled to English via the Portuguese form alcatraz ("gannet"), which is also the origin of the name of the former prison, Alcatraz. The Oxford English Dictionary notes that the word alcatraz was originally applied to the frigatebird; the modification to albatross was perhaps influenced by Latin albus, meaning "white", in contrast to frigatebirds, which are black.[15] In modern Portuguese, the word used for the bird, albatroz, is in turn derived from the English albatross.
They were once commonly known as goonie birds or gooney birds, particularly those of the North Pacific. In the Southern Hemisphere, the name mollymawk is still well established in some areas, which is a corrupted form of malle-mugge, an old Dutch name for the northern fulmar. The name Diomedea, assigned to the albatrosses by Linnaeus, references the mythical metamorphosis of the companions of the Greek warrior Diomedes into birds. Finally, the name for the order, Procellariiformes, comes from the Latin word procella meaning "a violent wind" or "a storm".[42]
In culture

Albatrosses have been described as "the most legendary of all birds".[39] An albatross is the central emblem in The Rime of the Ancient Mariner by Samuel Taylor Coleridge, representing the innocence and beauty of God's creation. The albatross metaphor is derived from this poem; someone bearing a burden or facing an obstacle is said to have "an albatross around his neck", the punishment given to the mariner who killed the albatross. A widespread myth holds that sailors believe shooting or harming an albatross is disastrous, due in part to the poem; in truth, sailors regularly killed and ate them,[24] as reported by James Cook in 1772. However, other sailors reportedly caught the birds but let them free again, possibly believing that albatrosses were the souls of lost sailors,[43] so killing them would bring bad luck.[44] The head of an albatross being caught with a hook is used as the emblem of the Cape Horners, i.e. sailors who have rounded Cape Horn on freighters under sail; captains of such ships even received themselves the title "albatrosses" in the Cape Horners' organisation.[44]
A captive albatross tormented by jeering sailors is also a metaphor for the social travails of the sensitive poète maudit in Charles Baudelaire's poem L'Albatros:
Le Poète est semblable au prince des nuées
Qui hante la tempête et se rit de l'archer;
Exilé sur le sol au milieu des huées,
Ses ailes de géant l'empêchent de marcher.
(The poet is like this prince of the clouds, who haunts the storm and mocks the archer; but exiled on earth surrounded by jeers, his giant wings make him helpless to walk.)
In golf, shooting three under par on a single hole has recently been termed scoring an "albatross", as a continuation on the birdie and eagle theme.[45]
Non-European mythologies
The Maori used the wing bones of the albatross to carve flutes.[46] In Hawaiian mythology, Laysan albatrosses are considered aumakua, being a sacred manifestation of the ancestors, and quite possibly also the sacred bird of Kāne.[47] Japanese mythology, by contrast, refers to the short-tailed albatross as aho-dori, "idiot bird", due to its insular habitat making it disregard terrestrial predators, and it was considered an excellent target for consumption.[48]
Birdwatching
Albatrosses are popular birds for birdwatchers, and their colonies are popular destinations for ecotourists. Regular birdwatching trips are taken out of many coastal towns and cities, such as Monterey, Dunedin, Kaikoura, Wollongong, Sydney, Port Fairy, Hobart, and Cape Town, to see pelagic seabirds. Albatrosses are easily attracted to these sightseeing boats by the deployment of fish oil and burley into the sea. Visits to colonies can be very popular: the northern royal albatross colony at Taiaroa Head in Dunedin, New Zealand, attracts 40,000 visitors a year,[2] and more isolated colonies are regular attractions on cruises to subantarctic islands.
Threats and conservation

In spite of often being accorded legendary status, albatrosses have not escaped either indirect or direct pressure from humans. Early encounters with albatrosses by Polynesians and Aleuts resulted in hunting and in some cases extirpation from some islands (such as Easter Island). As Europeans began sailing the world, they, too, began to hunt albatross, "fishing" for them from boats to serve at the table or blasting them for sport.[49] This sport reached its peak on emigration lines bound for Australia, and only died down when ships became too fast to fish from, and regulations forbade the discharge of weapons for safety reasons. In the 19th century, albatross colonies, particularly those in the North Pacific, were harvested for the feather trade, leading to the near-extinction of the short-tailed albatross.[15]
Of the 21 albatross species recognised by IUCN on their Red List, 19 are threatened, and the other two are near threatened.[50] Three species (as recognised by the IUCN) are considered critically endangered: the Amsterdam albatross, Tristan albatross, and waved albatross. One of the main threats is commercial longline fishing, as the albatrosses and other seabirds—which will readily feed on offal—are attracted to the set bait, become hooked on the lines and drown. An estimated 100,000 albatross per year are killed in this fashion. Unregulated pirate fisheries exacerbate the problem.[2][51]
On Midway Atoll, collisions between Laysan albatrosses and aircraft have resulted in human and bird deaths, as well as severe disruptions in military flight operations. Studies were made in the late 1950s and early 1960s that examined the results of control methods such as the killing of birds, the levelling and clearing of land to eliminate updrafts, and the destruction of annual nesting sites.[52] Tall structures such as traffic control and radio towers killed 3000 birds in flight collisions during 1964–1965 before the towers were taken down. Closure of Naval Air Facility Midway in 1993 eliminated the problem of collisions with military aircraft. Recent reductions in human activity on the island have helped reduce bird deaths, though lead paint pollution near military buildings continues to poison birds by ingestion.[53] Albatross plumes were popular in the early 20th century. In 1909 alone, over 300,000 albatrosses were killed on Midway Island and Laysan Island for their plumes.[17]
Another threat to albatrosses is introduced species, such as rats or feral cats, which directly attack albatrosses or their chicks and eggs. Albatrosses have evolved to breed on islands where land mammals are absent and have not developed defences against them. Even species as small as mice can be detrimental; on Gough Island, the chicks of Tristan albatrosses are attacked and eaten alive by introduced house mice.[54] Introduced species can have other indirect effects; cattle overgrazed essential cover on Amsterdam Island, threatening the Amsterdam albatross; on other islands, introduced plants reduce potential nesting habitat.[2]

Ingestion of plastic flotsam is another problem, one faced by many seabirds. The amount of plastic in the seas has increased dramatically since the first record in the 1960s, coming from waste discarded by ships, offshore dumping, litter on beaches, and waste washed to sea by rivers. It is impossible to digest and takes up space in the stomach or gizzard that should be used for food, or can cause an obstruction that starves the bird directly. Studies of birds in the North Pacific have shown that ingestion of plastics results in declining body weight and body condition.[55] This plastic is sometimes regurgitated and fed to chicks; a study of Laysan albatross chicks on Midway Atoll showed large amounts of ingested plastic in naturally dead chicks compared to healthy chicks killed in accidents. While not the direct cause of death, this plastic causes physiological stress and causes the chick to feel full during feedings, reducing its food intake and the chances of survival.[56]
Scientists and conservationists (most importantly BirdLife International and their partners, who run the Save the Albatross campaign) are working with governments and fishermen to find solutions to the threats albatrosses face. Techniques such as setting longline bait at night, dyeing the bait blue, setting the bait underwater, increasing the amount of weight on lines, and using bird scarers can all reduce the seabird bycatch.[57] For example, a collaborative study between scientists and fishermen in New Zealand successfully tested an underwater setting device for longliners, which set the lines below the reach of vulnerable albatross species.[58] The use of some of these techniques in the Patagonian toothfish fishery in the Falkland Islands is thought to have reduced the number of black-browed albatrosses taken by the fleet in the last 10 years.[59] Conservationists have also worked on the field of island restoration, removing introduced species that threaten native wildlife, which protects albatrosses from introduced predators.
One important step towards protecting albatrosses and other seabirds is the 2001 treaty, the Agreement on the Conservation of Albatrosses and Petrels, which came into force in 2004 and has been ratified by thirteen countries, Argentina, Australia, Brazil, Chile, Ecuador, France, New Zealand, Norway, Peru, South Africa, Spain, the United Kingdom and Uruguay.[60] The treaty requires these countries to take specific actions to reduce bycatch, pollution and to remove introduced species from nesting islands.[61]
Species
Current thinking divides the albatrosses into four genera. The number of species is a matter of debate. The IUCN and BirdLife International recognise 22 extant species (listed below),[62] ITIS recognise 21 (the 22 below minus T. steadi),[63] and one recent paper proposed a reduction to 13 (indicated in parentheses below), comprising the traditional 14 species minus D. amsterdamensis.[8]
- Great albatrosses (Diomedea)
- Wandering albatross (D. exulans)
- Antipodean albatross (D. (exulans) antipodensis)
- Amsterdam albatross (D. (exulans) amsterdamensis)
- Tristan albatross (D. (exulans) dabbenena)
- Northern royal albatross (D. (epomophora) sanfordi)
- Southern royal albatross (D. epomophora)
- North Pacific albatrosses (Phoebastria)
- Waved albatross (P. irrorata)
- Short-tailed albatross (P. albatrus)
- Black-footed albatross (P. nigripes)
- Laysan albatross (P. immutabilis)
- Mollymawks (Thalassarche)
- Black-browed albatross (T. melanophris )
- Campbell albatross (T. (melanophris) impavida)
- Shy albatross (T. cauta)
- White-capped albatross (T. (cauta) steadi)
- Chatham albatross (T. (cauta) eremita)
- Salvin's albatross (T. (cauta) salvini)
- Grey-headed albatross (T. chrysostoma)
- Atlantic yellow-nosed albatross (T. chlororhynchos
- Indian yellow-nosed albatross (T. (chlororhynchos) carteri)
- Buller's albatross (T. bulleri)
- Sooty albatrosses (Phoebetria)
- Sooty albatross (P. fusca)
- Light-mantled albatross (P. palpebrata)
See also
References
- Brands, Sheila (14 August 2008). "Systema Naturae 2000 / Classification – Family Diomedeidae". Project: The Taxonomicon. Archived from the original on 16 June 2009. Retrieved 17 February 2009.
- Brooke, M. (2004). Albatrosses and Petrels Across the World Oxford University Press, Oxford, UK ISBN 0-19-850125-0
- Robertson, C. J. R. and Nunn, G. B. (1998) "Towards a new taxonomy for albatrosses" in: Proceedings First International Conference on the Biology and Conservation of Albatrosses, G.Robertson & R.Gales (Eds), Chipping Norton:Surrey Beatty & Sons, 13–19,
- Alexander, W. B.; Fleming, C. A.; Falla, R. A.; Kuroda, N. H.; Jouanin, C.; Rowan, M. K.; Murphy, R. C.; Serventy, D. L.; Salomonsen, F.; et al. (1965). "Correspondence: The families and genera of the petrels and their names". Ibis. 107 (3): 401–5. doi:10.1111/j.1474-919X.1965.tb07326.x.
- Nunn, G. B.; Cooper, J.; Jouventin, P.; Robertson, C. J. R.; Robertson, G. G. (1996). "Evolutionary relationships among extant albatrosses (Procellariiformes: Diomedeidae) established from complete cytochrome-b gene sequences" (PDF). Auk. 113 (4): 784–801. doi:10.2307/4088857. JSTOR 4088857.
- Double, M.C. & Chambers, G.K., (2004). "The need for the parties to the Agreement on Albatrosses and Petrels (ACAP) to establish a robust, defendable and transparent decision-making process for the construction and maintenance of their species lists ". Proceedings of the Scientific Meeting of Agreement on Albatrosses and Petrels (ACAP), Hobart, Australia, 8–9 November 2004
- Burg, T.M.; Croxall, J.P. (2004). "Global population structure and taxonomy of the wandering albatross species complex" (PDF). Molecular Ecology. 13 (8): 2345–2355. doi:10.1111/j.1365-294X.2004.02232.x. PMID 15245406.
- Penhallurick, J.; Wink, M. (2004). "Analysis of the taxonomy and nomenclature of the Procellariiformes based on complete nucleotide sequences of the mitochondrial cytochrome b gene" (PDF). Emu. 104 (2): 125–147. doi:10.1071/MU01060.
- Rheindt, F. E.; Austin, J. (2005). "Major analytical and conceptual shortcomings in a recent taxonomic revision of the Procellariiformes – A reply to Penhallurick and Wink (2004)" (PDF). Emu. 105 (2): 181–186. doi:10.1071/MU04039.
- Mayr, G.; Tennyson, A.J.D. (2019). A small, narrow-beaked albatross from the Pliocene of New Zealand demonstrates a higher past diversity in the feeding ecology of the Diomedeidae. Ibis
- Mayr, Gerald; Goedert, James L. (2017). "Oligocene and Miocene albatross fossils from Washington State (USA) and the evolutionary history of North Pacific Diomedeidae". The Auk. 134 (3): 659–671. doi:10.1642/AUK-17-32.1.
- Olson, S.L.; Hearty, P.J. (2003). "Probable extirpation of a breeding colony of Short-tailed Albatross (Phoebastria albatrus) on Bermuda by Pleistocene sea-level rise". Proceedings of the National Academy of Sciences. 100 (22): 12825–12829. Bibcode:2003PNAS..10012825O. doi:10.1073/pnas.1934576100. PMC 240703. PMID 14566060.
- Gelasian, formerly Late Pliocene
- Lequette, B.; Verheyden, C.; Jowentin, P. (1989). "Olfaction in Subantarctic seabirds: Its phylogenetic and ecological significance" (PDF). The Condor. 91 (3): 732–735. doi:10.2307/1368131. JSTOR 1368131.
- Tickell, W. L. N. (2000). Albatrosses. Sussex: Pica Press. ISBN 1-873403-94-1.
- Ehrlich, Paul R.; Dobkin, David, S.; Wheye, Darryl (1988). The Birders Handbook (First ed.). New York, NY: Simon & Schuster. pp. 29–31. ISBN 978-0-671-65989-9.
- Humann, Alec; Brinkley, Edward S. (2001). "Albatrosses". In Elphick, Chris; Dunning Jr., John B.; Sibley, David Allen (eds.). The Sibley Guide to Bird Life and Behavior. illustrated by David Allen Sibley (1st. ed.). New York: Alfred A. Knopf. pp. 132–135. ISBN 978-0-679-45123-5.
- Pennycuick, C. J. (1982). "The flight of petrels and albatrosses (Procellariiformes), observed in South Georgia and its vicinity". Philosophical Transactions of the Royal Society of London B. 300 (1098): 75–106. Bibcode:1982RSPTB.300...75P. doi:10.1098/rstb.1982.0158. JSTOR 2395926.
- Weimerskirch, H; Guionnet, T; Martin, J; Shaffer, SA; Costa, DP. (2000). "Fast and fuel efficient? Optimal use of wind by flying albatrosses". Proceedings of the Royal Society of London. Series B: Biological Sciences. 267 (1455): 1869–74. doi:10.1098/rspb.2000.1223. PMC 1690761. PMID 11052538.
- "Animal Facts – Albatross: Do albatrosses sleep while flying?". OnlineMathLearning.com. Retrieved 6 April 2012.
- Peter Ryan (February 1997). "The Wandering Albatross" (PDF).
- Warham, J. (1996). The Behaviour, Population, Biology and Physiology of the Petrels. London: Academic Press. ISBN 0-12-735415-8.
- Daniel Stone (7 September 2012). "Albatross's Effortless Flight Decoded—May Influence Future Planes". National Geographic.
- Cocker, M., & Mabey, R., (2005) Birds Britannica London:Chatto & Windus, ISBN 0-7011-6907-9
- Croxall, J. P.; Silk, J.R.D.; Phillips, R.A.; Afanasyev, V.; Briggs, D.R. (2005). "Global Circumnavigations: Tracking year-round ranges of nonbreeding Albatrosses". Science. 307 (5707): 249–250. Bibcode:2005Sci...307..249C. doi:10.1126/science.1106042. PMID 15653503.
- Croxall, J.P.; Prince, P.A. (1994). "Dead or alive, night or day: how do albatrosses catch squid?". Antarctic Science. 6 (2): 155–162. Bibcode:1994AntSc...6..155C. doi:10.1017/S0954102094000246.
- Spear, Larry; David G. Ainley (1993). "Kleptoparasitism by Kermadec Petrels, Jaegers, and Skuas in the Eastern Tropical Pacific: Evidence of Mimicry by Two Species of Pterodroma". The Auk. 110 (2): 222–233. JSTOR 4088550.
- Prince, P.A.; Huin, N.; Weimerskirch, H. (1994). "Diving depths of albatrosses". Antarctic Science. 6 (3): 353–354. Bibcode:1994AntSc...6..353P. doi:10.1017/S0954102094000532.
- Cobley, N.D. (1996). "An observation of live prey capture by a Black-browed Albatross Diomedea melanophrys" (PDF). Marine Ornithology. 24: 45–46.
- Robertson, C. J. R. (2003). "Albatrosses (Diomedeidae)". In Hutchins, Michael (ed.). Grzimek's Animal Life Encyclopedia. 8 Birds: 1. (2 ed.). Farmington Hills, MI: Gale Group. pp. 113–116. ISBN 0-7876-5784-0.
- Fisher, H.I. (1976). "Some dynamics of a breeding colony of Laysan Albatrosses". Wilson Bulletin. 88 (1): 121–142. JSTOR 4160718.
- Dell'Amore, Christine (22 February 2013). "Oldest Known Wild Bird Hatches Chick at 62". National Geographic. Retrieved 18 February 2017.
- Arnold, Carrie (17 February 2017). "World's Oldest Wild Bird Has Baby at 66". National Geographic. Retrieved 18 February 2017.
- Jouventin, P.; Monicault, G. de; Blosseville, J.M. (1981). "La danse de l'albatros, Phoebetria fusca". Behaviour. 78 (1–2): 43–80. doi:10.1163/156853981X00257.
- Pickering, S.P.C.; Berrow, S.D. (2001). "Courtship behaviour of the Wandering Albatross Diomedea exulans at Bird Island, South Georgia" (PDF). Marine Ornithology. 29: 29–37.
- Anderson, D.J. & Cruz, F. (1998) "Biology and management of the Waved Albatross at the Galapagos Islands, pp. 105–109 in Albatross Biology and Conservation (Robertson, G. & Gales, R. eds) Chipping Norton:Surrey Beatty and & Sons ISBN 0-949324-82-5
- Warham, J. (1990) The Petrels – Their Ecology and Breeding Systems London: Academic Press.
- Warham, J. (1976). "The incidence, function and ecological significance of petrel stomach oils" (PDF). Proceedings of the New Zealand Ecological Society. 24 (3): 84–93. Archived from the original (PDF) on July 24, 2006.
- Carboneras, C. (1992) "Family Diomedeidae (Albatross)" in Handbook of Birds of the World Vol 1. Barcelona:Lynx Edicions, ISBN 84-87334-10-5
- Åkesson, S.; Weimerskirch, H. (2005). "Albatross Long-Distance Navigation: Comparing Adults And Juveniles" (PDF). Journal of Navigation. 58 (3): 365–373. Bibcode:2005JNav...58..365A. doi:10.1017/S0373463305003401.
- Phillips, R.A..; Cooper, J.; Burg, T.M. (2018). "Breeding‐site vagrancy and hybridization in albatrosses". Ibis. 160 (4): 907–913. doi:10.1111/ibi.12622.
- Gotch, A. F. (1995) [1979]. "Albatrosses, Fulmars, Shearwaters, and Petrels". Latin Names Explained. A Guide to the Scientific Classifications of Reptiles, Birds & Mammals. New York, NY: Facts on File. p. 190. ISBN 0-8160-3377-3.
- Eyers, Jonathan (2011). Don't Shoot the Albatross!: Nautical Myths and Superstitions. A&C Black, London, UK. ISBN 978-1-4081-3131-2.
- From the website of the Cape Horners' organisation caphorniers.cl: A.I.C.H. Emblem. Archived 2011-06-15 at the Wayback Machine (retrieved 24 February 2011). Synthesis of an article written by the International Secretary General of the A.I.C.H., Captain Roger GHYS, as published in LE COURRIER DU CAP N°3 of December, 1999. By Rear Admiral Roberto BENAVENTE, President, Chilean Section AICH
- The Players Championship – What Golf Betting Is All About. scottishgolfhistory.net
- Mclean, Mervyn (1982). "A Chronological and Geographical Sequence of Maori Flute Scales". Man. 17 (1): 123–157. doi:10.2307/2802105. JSTOR 2802105.
- Hor Osterlund, Holy Mōlī: Albatross and Other Ancestors, Oregon State University Press
- "Archived copy". Archived from the original on 2017-11-13. Retrieved 2017-11-13.CS1 maint: archived copy as title (link)
- Safina, C. (2002) Eye of the Albatross: Visions of Hope and Survival New York: Henry Holt & Company ISBN 0-8050-6229-7
- IUCN (2004). Red List: Albatross Species Archived April 27, 2006, at the Wayback Machine. Retrieved July 27, 2007.
- Brothers, NP. (1991). "Albatross mortality and associated bait loss in the Japanese longline fishery in the southern ocean". Biological Conservation. 55 (3): 255–268. doi:10.1016/0006-3207(91)90031-4.
- Fisher, Harvey I. (May 1966). "Airplane-Albatross Collisions on Midway Atoll". The Condor. 68 (3): 229–242. doi:10.2307/1365556. JSTOR 1365556. S2CID 88307541.
- Audubon Watchlist. Laysan Albatross (Phoebastria immutabilis) Archived 2008-02-03 at the Wayback Machine. audubon2.org
- Amos, Jonathan (24 July 2005). Albatross chicks attacked by mice. BBC News.
- Spear, L.B.; Ainley, D.G.; Ribic, C.A. (1995). "Incidence of plastic in seabirds from the tropical Pacific, 1984–91: relation with distribution of species, sex, age, season, year and body weight". Marine Environmental Research. 40 (2): 123–146. doi:10.1016/0141-1136(94)00140-K.
- Auman, H.J., Ludwig, J.P., Giesy, J.P., Colborn, T., (1997) "Plastic ingestion by Laysan Albatross chicks on Sand Island, Midway Atoll, in 1994 and 1995." Archived 2005-10-30 at the Wayback Machine in Albatross Biology and Conservation, (ed by G. Robinson and R. Gales). Surrey Beatty & Sons:Chipping Norton. pp. 239–44
- Food and Agriculture Organisation (1999) "The incidental catch of seabirds by longline fisheries: worldwide review and technical guidelines for mitigation" Archived June 29, 2006, at the Wayback Machine. FAO Fisheries Circular No. 937. Food and Agriculture Organisation of the United Nations, Rome.
- O'Toole, Decland; Molloy, Janice (2000). "Preliminary performance assessment of an underwater line setting device for pelagic longline fishing". New Zealand Journal of Marine and Freshwater Research. 34 (3): 455–461. doi:10.1080/00288330.2000.9516947.
- Reid, A.T.; Sullivan, B.J.; Pompert, J.; Enticott, J.W.; Black, A.D. (2004). "Seabird mortality associated with Patagonian Toothfish (Dissostichus eleginoides) longliners in Falkland Islands waters". Emu. 104 (4): 317–325. doi:10.1071/MU03002.
- Agreement on the Conservation of Albatrosses and Petrels. "Parties To ACAP". Agreement on the Conservation of Albatrosses and Petrels. Retrieved 22 January 2011.
- ACAP (2009). "Amended, Third Session of the Meeting of the Parties". Agreement on the Conservation of Albatrosses and Petrels. Retrieved 22 January 2011.
- "IUCN Red List of Threatened Species: Version 2014.2 (search for "DIOMEDEIDAE")". Retrieved 30 October 2014.
- "Integrated Taxonomic Information System (ITIS)". Retrieved 30 October 2014.
External links
| Library resources about Albatross |
| Look up albatross in Wiktionary, the free dictionary. |
| Wikimedia Commons has media related to Diomedeidae. |
| Wikispecies has information related to Diomedeidae |
- "Diomedeidae". Integrated Taxonomic Information System. Retrieved 4 May 2006.
- HANZAB complete species list (Handbook of Australian, New Zealand and Antarctic Birds.)
- BirdLife International Save the Albatross campaign
- The Agreement for the Conservation of Albatrosses and Petrels (ACAP)
- Albatross: Don Roberson's family page
- Tracking Ocean Wanderers The global distribution of albatrosses and petrels: Results from the Global Procellariiform Tracking Workshop, 1–5 September 2003, Gordon's Bay, South Africa. BirdLife International
- Albatross videos on the Internet Bird Collection
- Albatross species profile at the WWF
- "Recovery plan for albatrosses in the Chatham Islands 2001–2011" (PDF). Department of Conservation, Wellington, New Zealand. 2001. Archived from the original (PDF) on 2008-04-14. Retrieved 2007-09-28.

- "Albatross". New International Encyclopedia. 1905.
- "Albatross". Encyclopædia Britannica (11th ed.). 1911.
- "Albatross". Encyclopedia Americana. 1920.
- . Collier's New Encyclopedia. 1921.