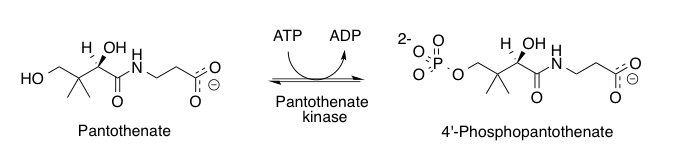
Pantothenate kinase
Pantothenate kinase (EC 2.7.1.33, PanK; CoaA) is the first enzyme in the Coenzyme A (CoA) biosynthetic pathway. It phosphorylates pantothenate (vitamin B5) to form 4'-phosphopantothenate at the expense of a molecule of adenosine triphosphate (ATP). It is the rate-limiting step in the biosynthesis of CoA.[1][2]
| Pantothenate kinase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| EC number | 2.7.1.33 | ||||||||
| CAS number | 9026-48-6 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
CoA is a necessary cofactor in all living organisms. It acts as the major acyl group carrier in many important cellular processes, such as the citric acid cycle (tricarboxylic acid cycle) and fatty acid metabolism. Consequently, pantothenate kinase is a key regulatory enzyme in the CoA biosynthetic pathway.[3]
Types
Three distinct types of PanK has been identified - PanK-I (found in bacteria), PanK-II (mainly found in eukaryotes, but also in the Staphylococci) and PanK-III, also known as CoaX (found in bacteria). Eukaryotic PanK-II enzymes often occur as different isoforms, such as PanK1, PanK2, PanK3 and PanK4. In humans, multiple PanK isoforms are expressed by four genes. PANK1 gene encodes the PanK1α and PanK1β forms, and PANK2 and PANK3 encode PanK2 and PanK3, respectively.[4]
Structure
PanK-II
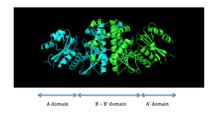
PanK-II contains two protein domains, as illustrated in Figure 1. The A domain and A' domain each has a glycine-rich loop (sequence GXXXXGKS; P loop) that is characteristic of nucleotide-binding sites; this is where ATP is assumed to bind.[5] located between residues 95 and 102 on the A domain
The two ATP binding sites display cooperative behavior. The dimerization interface consists of two long helices, one from each monomer, that interact with each other. The C-terminal ends of the helices are held together by van der Waals interactions between valine and methionine residues of each monomer. The middle of the helices is attached by hydrogen bonds between asparagine residues. At the N-terminal end, each helix widens and forms a four-helix bundle with two shorter helices. This bundle consists of a hydrophobic core formed by non-polar residues that utilize van der Waals forces to further stabilize the dimer.[4]
In the active site, pantothenate is oriented by hydrogen bonds between pantothenate and the side chains of aspartate, tyrosine, histidine, tyrosine, and asparagine residues.[6] Asparagine, histidine, and arginine residues are involved in catalysis.
Human PanK-II isoforms PanK1α, PanK1β, PanK2, and PanK3 have a common, highly homologous catalytic core of approximately 355 residues.[4] PanK1α and PanK1β are both encoded by the PANK1 gene and have the same catalytic domain of 363 amino acids, encoded by exons 2 through 7. The PanK1α transcript starts with exon 1α that encodes a 184-residue regulatory domain at the N-terminus. This region allows for feedback inhibition by free CoA and acyl-CoA and regulation by acetyl-CoA and malonyl-CoA. On the other hand, the PanK1β transcript starts with exon 1β, which produces a 10-residue N-terminus that does not include a feedback regulatory domain.[7]
PanK-III
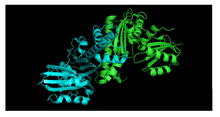
PanK-III also contains two protein domains, and the key catalytic residues of PanK-II are conserved. The monomer units of PanK-II and PanK-III are virtually identical, but they have distinctly different dimer assemblies. A study between the structures of Staphylococcus aureus type II and the Pseudomonas aeruginosa type III demonstrate that the PanK-II monomer has a loop region that is absent from the PanK-III monomer, and the PanK-III monomer has a loop region that is absent from the PanK-II monomer.[8] This minor variation has a crucial difference on the dimerization interface in which the helices of the PanK-II dimer coil around one another and the helices of the PanK-III dimer interact at a 70° angle (Figure 2).[9]
As a result of this difference in dimerization interface between PanK-II and PanK-III, the conformations of the substrate binding sites for ATP and pantothenate are also distinct.[10][11]
Catalytic Mechanism
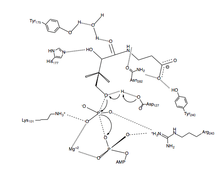
PanK-II
A proposed mechanism of the phosphoryl transfer reaction of PanK-II is a concerted mechanism with a dissociative transition state.
First, the ATP binds at the binding groove created by residues of the P loop and nearby residues. Here, the conserved lysine (Lys-101) is the key residue required for ATP binding.[12][13] Additionally, the side chains of residues Lys-101, Ser-102, Glu-199, and Arg-243 orient the nucleotide in the binding groove. The pantothenate is bound and oriented by forming hydrogen bond interactions with residues Asp-127, Tyr-240, Asn-282, Tyr-175, and His-177.[6] When both ATP and pantothenate are bound, Asp-127 deprotonates the C1 hydroxyl group of pantothenate. The oxygen from the pantothenate then attacks the γ-phosphate of the bound ATP. Here, charge stabilization of β- and γ-phosphate groups is achieved by Arg-243, Lys-101, and a coordinated Mg2+ ion.[14] In this concerted mechanism, the planar phosphorane of the γ-phosphate is transferred in-line to the attacking oxygen of pantothenate.[6] Finally, 4'-phosphopantothenate dissociates from PanK, followed by ADP.
Regulation of pantothenate kinase
PanK-II
The regulation of pantothenate kinase is essential to controlling the intracellular CoA concentration.[15] Pantothenate kinase is regulated via feedback inhibition by CoA and its thioesters (i.e., acetyl-CoA, malonyl-CoA).[16] CoA inhibits PanK activity by competitively binding to the ATP binding site and preventing ATP binding to Lys-101.[12][13]
Although CoA binds at the same site as ATP, they bind in distinct orientations, and their adenine moieties interact with the enzyme with nonoverlapping sets of residues. His-177, Phe-247, and Arg-106 are necessary for CoA recognition but not for ATP, and while Asn-43 and His-307 interact with the adenine base of ATP, His-177 and Phe-247 interact with the adenine base of CoA.[14] Both molecules use Lys-101 to neutralize the charge on their respective phosphodiesters.
Nonesterified CoA has more potent inhibition than its thioesters. This phenomenon is best explained by the tight fit of the thiol group with the surrounding aromatic residues, Phe-244, Phe-259, Tyr-262, and Phe-252. Free CoA has an optimal fit, but when an acyl group is attached to CoA, the steric hindrance makes it difficult for the thioester to interact with Phe-252. Thus, the inhibition by thioesters is less effective than that by nonesterified CoA.[14]
Genes
In humans:
The PANK2 gene encodes for PanK2, which regulates the formation of CoA in mitochondria, the cell’s energy-producing centers.[17] PANK2 mutation is the cause of Pantothenate kinase-associated neurodegeneration, formerly called Hallervorden-Spatz syndrome. This rare disease presents with profound dystonia, spasticity and is often fatal.
References
- Robishaw JD, Berkich D, Neely JR (September 1982). "Rate-limiting step and control of coenzyme A synthesis in cardiac muscle". The Journal of Biological Chemistry. 257 (18): 10967–72. PMID 7107640.
- Yang K, Eyobo Y, Brand LA, Martynowski D, Tomchick D, Strauss E, Zhang H (August 2006). "Crystal structure of a type III pantothenate kinase: insight into the mechanism of an essential coenzyme A biosynthetic enzyme universally distributed in bacteria". Journal of Bacteriology. 188 (15): 5532–40. doi:10.1128/JB.00469-06. PMC 1540032. PMID 16855243.
- Leonardi R, Zhang YM, Rock CO, Jackowski S (2005-03-01). "Coenzyme A: back in action". Progress in Lipid Research. 44 (2–3): 125–53. doi:10.1016/j.plipres.2005.04.001. PMID 15893380.
- Hong BS, Senisterra G, Rabeh WM, Vedadi M, Leonardi R, Zhang YM, Rock CO, Jackowski S, Park HW (September 2007). "Crystal structures of human pantothenate kinases. Insights into allosteric regulation and mutations linked to a neurodegeneration disorder". The Journal of Biological Chemistry. 282 (38): 27984–93. doi:10.1074/jbc.M701915200. PMID 17631502.
- Saraste M, Sibbald PR, Wittinghofer A (November 1990). "The P-loop--a common motif in ATP- and GTP-binding proteins". Trends in Biochemical Sciences. 15 (11): 430–4. doi:10.1016/0968-0004(90)90281-F. PMID 2126155.
- Ivey RA, Zhang YM, Virga KG, Hevener K, Lee RE, Rock CO, Jackowski S, Park HW (August 2004). "The structure of the pantothenate kinase.ADP.pantothenate ternary complex reveals the relationship between the binding sites for substrate, allosteric regulator, and antimetabolites". The Journal of Biological Chemistry. 279 (34): 35622–9. doi:10.1074/jbc.M403152200. PMID 15136582.
- Rock CO, Karim MA, Zhang YM, Jackowski S (2002). "The murine pantothenate kinase (Pank1) gene encodes two differentially regulated pantothenate kinase isozymes". Gene. 291 (1–2): 35–43. doi:10.1016/S0378-1119(02)00564-4. PMID 12095677.
- Yang K, Strauss E, Huerta C, Zhang H (February 2008). "Structural basis for substrate binding and the catalytic mechanism of type III pantothenate kinase". Biochemistry. 47 (5): 1369–80. doi:10.1021/bi7018578. PMID 18186650.
- Hong BS, Yun MK, Zhang YM, Chohnan S, Rock CO, White SW, Jackowski S, Park HW, Leonardi R (August 2006). "Prokaryotic type II and type III pantothenate kinases: The same monomer fold creates dimers with distinct catalytic properties". Structure. 14 (8): 1251–61. doi:10.1016/j.str.2006.06.008. PMID 16905099.
- Brand LA, Strauss E (May 2005). "Characterization of a new pantothenate kinase isoform from Helicobacter pylori". The Journal of Biological Chemistry. 280 (21): 20185–8. doi:10.1074/jbc.C500044200. PMID 15795230.
- Choudhry AE, Mandichak TL, Broskey JP, Egolf RW, Kinsland C, Begley TP, Seefeld MA, Ku TW, Brown JR, Zalacain M, Ratnam K (June 2003). "Inhibitors of pantothenate kinase: novel antibiotics for staphylococcal infections". Antimicrobial Agents and Chemotherapy. 47 (6): 2051–5. doi:10.1128/AAC.47.6.2051-2055.2003. PMC 155856. PMID 12760898.
- Song WJ, Jackowski S (October 1994). "Kinetics and regulation of pantothenate kinase from Escherichia coli". The Journal of Biological Chemistry. 269 (43): 27051–8. PMID 7929447.
- Song WJ, Jackowski S (October 1992). "Cloning, sequencing, and expression of the pantothenate kinase (coaA) gene of Escherichia coli". Journal of Bacteriology. 174 (20): 6411–7. doi:10.1128/jb.174.20.6411-6417.1992. PMC 207592. PMID 1328157.
- Yun M, Park CG, Kim JY, Rock CO, Jackowski S, Park HW (September 2000). "Structural basis for the feedback regulation of Escherichia coli pantothenate kinase by coenzyme A". The Journal of Biological Chemistry. 275 (36): 28093–9. doi:10.1074/jbc.M003190200. PMID 10862768.
- Jackowski S, Rock CO (December 1981). "Regulation of coenzyme A biosynthesis". Journal of Bacteriology. 148 (3): 926–32. PMC 216294. PMID 6796563.
- Rock CO, Park HW, Jackowski S (June 2003). "Role of feedback regulation of pantothenate kinase (CoaA) in control of coenzyme A levels in Escherichia coli". Journal of Bacteriology. 185 (11): 3410–5. doi:10.1128/JB.185.11.3410-3415.2003. PMC 155388. PMID 12754240.
- "PANK2 gene". Genetics Home Reference. 2016-02-22. Retrieved 2016-02-29.
External links
- Pantothenate+kinase at the US National Library of Medicine Medical Subject Headings (MeSH)
- EC 2.7.1.33