Walker motifs
The Walker A and Walker B motifs are protein sequence motifs, known to have highly conserved three-dimensional structures. These were first reported in ATP-binding proteins by Walker and co-workers in 1982.[1]
Walker A motif
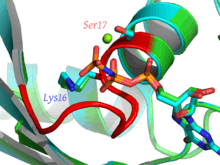
Walker A motif, also known as the Walker loop, or P-loop, or phosphate-binding loop, is a motif in proteins that is associated with phosphate binding. The motif has the pattern G-x(4)-GK-[TS], where G, K, T and S denote glycine, lysine, threonine and serine residues respectively, and x denotes any amino acid. It is present in many ATP or GTP utilizing proteins; it is the β phosphate of the nucleotide that is bound. The lysine (K) residue in the Walker A motif, together with the main chain NH atoms, are crucial for nucleotide-binding.[2] It is a glycine-rich loop preceded by a beta strand and followed by an alpha helix; these features are typically part of an α/β domain with four strands sandwiched between two helices on each side. The phosphate groups of the nucleotide are also coordinated to a divalent cation such as a magnesium, calcium, or manganese(II) ion.[3]
Apart from the conserved lysine, a feature of the P-loop used in phosphate binding is a compound LRLR nest[4] comprising the four residues xxGK, as above, whose main chain atoms form a phosphate-sized concavity with the NH groups pointing inwards. The synthetic hexapeptide SGAGKT has been shown[5] to bind inorganic phosphate strongly; since such a short peptide does not form an alpha helix, this suggests that it is the nest, rather than being at the N-terminus of a helix, that is the main phosphate binding feature.
The Walker A motif is best known for its presence in ATP- and GTP-binding proteins, and is also found in a variety of proteins with phosphorylated substrates. These include ATP synthase (α and β subunits), myosin, transducin, helicases, kinases, AAA proteins, G-proteins, RecA, protein tyrosine phosphatases (see below) and pyridoxal phosphate utilizing enzymes such as cysteine synthase.[6][7][8]
Upon nucleotide hydrolysis the loop does not significantly change the protein conformation, but stays bound to the remaining phosphate groups. Walker motif A-binding has been shown to cause structural changes in the bound nucleotide, along the line of the induced fit model of enzyme binding.
PTPs (protein tyrosine phosphatases) that catalyse the hydrolysis of an inorganic phosphate from a phosphotyrosine residue (the reverse of a tyrosine kinase reaction) contain a motif which folds into a P-loop-like structure with an arginine in the place of the conserved lysine. The conserved sequence of this motif is C-x(5)-R-[ST], where C and R denote cysteine and arginine residues respectively.[9]
A-loop
The A-loop (aromatic residue interacting with the adenine ring of ATP) refers to conserved aromatic amino acids, essential for ATP-binding, found in about 25 amino acids upstream of the Walker A motif in a subset of P-loop proteins.[10]
Walker B motif
Walker B motif is a motif in most P-loop proteins situated well downstream of the A-motif. The consensus sequence of this motif was reported to be [RK]-x(3)-G-x(3)-LhhhD, where R, K, G, L and D denote arginine, lysine, glycine, leucine and aspartic acid residues respectively, x represents any of the 20 standard amino acids and h denotes a hydrophobic amino acid.[1] This motif was changed to be hhhhDE, where E denotes a glutamate residue.[2] The aspartate and glutamate also form a part of the DEAD/DEAH motifs found in helicases. The aspartate residue co-ordinates magnesium ions, and the glutamate is essential for ATP hydrolysis.[2] There is considerable variability in the sequence of this motif, with the only invariant features being a negatively charged residue following a stretch of bulky, hydrophobic amino acids.[11]
See also
- Activation loop
- Autophosphorylation
- Ca2+/calmodulin-dependent protein kinase
- Cell signaling
- Cyclin-dependent kinase
- G protein-coupled receptor
- Nucleoside-diphosphate kinase
- Phosphatase
- Phosphatidylinositol phosphate kinases
- Phospholipid
- Phosphoprotein
- Phosphorylation
- Phosphotransferase
- Signal transduction
- Thymidine kinase
- Thymidine kinase in clinical chemistry
- Thymidylate kinase
- Wall-associated kinase
References
- Walker JE, Saraste M, Runswick MJ, Gay NJ (1982). "Distantly related sequences in the alpha- and beta-subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold". EMBO J. 1 (8): 945–51. doi:10.1002/j.1460-2075.1982.tb01276.x. PMC 553140. PMID 6329717.
- Hanson PI, Whiteheart SW (July 2005). "AAA+ proteins: have engine, will work". Nat. Rev. Mol. Cell Biol. 6 (7): 519–29. doi:10.1038/nrm1684. PMID 16072036.
- Bugreev, D. V.; Mazin, A. V. (2004). "Ca2+ activates human homologous recombination protein Rad51 by modulating its ATPase activity". Proceedings of the National Academy of Sciences of the United States of America. 101: 9988–9993. doi:10.1073/pnas.0402105101. PMC 454202. PMID 15226506.
- Watson, JD; Milner-White EJ (2002). "A novel main-chain anion binding site in proteins: the nest. A particular combination of phi,psi values for successive residues gives rise to anion binding sites that occur commonly and are found at functionally important regions". Journal of Molecular Biology. 315: 171–182. doi:10.1006/jmbi.2001.5227. PMID 11779237.
- Bianchi, A; Giorgi C; Ruzza P; Toniolo C; Milner-White EJ (2012). "A synthetic peptide designed to resemble a proteinaceous P-loop nest is shown to bind inorganic phosphate". Proteins. 80: 1418–1424. doi:10.1002/prot.24038. PMID 22275093.
- Stryer, Lubert; Berg, Jeremy Mark; Tymoczko, John L. (2002). Biochemistry. San Francisco: W.H. Freeman. ISBN 0-7167-4684-0.
- Ramakrishnan, C; Dani, VS; Ramasarma, T (October 2002). "A conformational analysis of Walker motif A [GXXXXGKT(S)] in nucleotide-binding and other proteins". Protein Engineering. 15 (10): 783–798. doi:10.1093/protein/15.10.783. PMID 12468712. Retrieved 16 October 2013.
- Saraste M, Sibbald PR, Wittinghofer A (November 1990). "The P-loop--a common motif in ATP- and GTP-binding proteins". Trends Biochem. Sci. 15 (11): 430–4. doi:10.1016/0968-0004(90)90281-f. PMID 2126155.
- Zhang M, Stauffacher CV, Lin D, Van Etten RL (August 1998). "Crystal structure of a human low molecular weight phosphotyrosyl phosphatase. Implications for substrate specificity". J. Biol. Chem. 273 (34): 21714–20. doi:10.1074/jbc.273.34.21714. PMID 9705307.
- Ambudkar SV, Kim IW, Xia D, Sauna ZE (February 2006). "The A-loop, a novel conserved aromatic acid subdomain upstream of the Walker A motif in ABC transporters, is critical for ATP binding". FEBS Lett. 580 (4): 1049–55. doi:10.1016/j.febslet.2005.12.051. PMID 16412422.
- Koonin EV (June 1993). "A common set of conserved motifs in a vast variety of putative nucleic acid-dependent ATPases including MCM proteins involved in the initiation of eukaryotic DNA replication". Nucleic Acids Res. 21 (11): 2541–7. doi:10.1093/nar/21.11.2541. PMC 309579. PMID 8332451.