Nucleotidyltransferase
Nucleotidyltransferases are transferase enzymes of phosphorus-containing groups, e.g., substituents of nucleotidylic acids or simply nucleoside monophosphates. The general reaction of transferring a nucleoside monophosphate moiety from A to B, can be written as:
- A-P-N + B A + B-P-N

For example, in the case of polymerases, A is pyrophosphate and B is the nascent polynucleotide. They are classified under [[Enzyme Commw9iudn owiddmlsi;ja \l- groups
- Adenylyltransferases, which transfer adenylyl- groups
- Guanylyltransferases, which transfer guanylyl- groups
- Cytitidylyltransferases, which transfer cytidylyl- groups
- Thymidylyltransferases, which transfer thymidylyl- groups
Role in metabolism
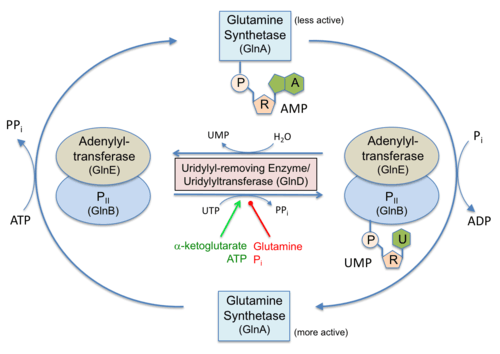
Many metabolic enzymes are modified by nucleotidyltransferases. The attachment of an AMP (adenylylation) or UMP (uridylylation) can activate or inactivate an enzyme or change its specificity (see figure). These modifications can lead to intricate regulatory networks that can finely tune enzymatic activities so that only the needed compounds are made (here: glutamine).[1]
Role in DNA repair mechanisms
Nucleotidyl transferase is a component of the repair pathway for single nucleotide base excision repair. This repair mechanism begins when a single nucleotide is recognized by DNA glycosylase as incorrectly matched or has been mutated in some way (UV light, chemical mutagen, etc.), and is removed. Later, a nucleotidyl tranferase is used to fill in the gap with the correct base, using the template strand as the reference.[2]
References
- Voet D, Voet JG, Pratt CW (2008). Fundamentals of Biochemistry (3rd ed.). John Wiley & Sons, pp. 767
-
Yuan Liu; Rajendra Prasad; William A. Beard; Padmini S. Kedar; Esther W. Hou; David D. Shock; Samuel H. Wilson (May 4, 2007). "Coordination of Steps in Single-nucleotide Base Excision Repair Mediated by Apurinic/Apyrimidinic Endonuclease 1 and DNA Polymerase β". Journal of Biological Chemistry. 282 (18): 13532–13541. doi:10.1074/jbc.M611295200. PMC 2366199. PMID 17355977.
The enzymes and accessory factors involved in the BER subpathways in mammalian cells have received considerable attention. As summarized above, five distinct enzymatic reaction are involved during SN-BER. These are 1) removal of a modified base by a lesion-specific monofunctional DNA N-glycosylase, 2) 5'-incision of the abasic site by a hydrolytic strand incision enzyme, 3) DNA synthesis by a nucleotidyltransferase, 4) removal of the 5'-dRP group by a'-elimination reaction, and 5) nick sealing by DNA ligase (36, 37)
External links
- Nucleotidyltransferases at the US National Library of Medicine Medical Subject Headings (MeSH)