Transposase
Transposase is an enzyme that binds to the end of a transposon and catalyses its movement to another part of the genome by a cut and paste mechanism or a replicative transposition mechanism. The word "transposase" was first coined by the individuals who cloned the enzyme required for transposition of the Tn3 transposon.[1] The existence of transposons was postulated in the late 1940s by Barbara McClintock, who was studying the inheritance of maize, but the actual molecular basis for transposition was described by later groups. McClintock discovered that pieces of the chromosomes changed their position, jumping from one chromosome to another. The repositioning of these transposons (which coded for color) allowed other genes for pigment to be expressed.[2] Transposition in maize causes changes in color; however, in other organisms, such as bacteria, it can cause antibiotic resistance.[2] Transposition is also important in creating genetic diversity within species and adaptability to changing living conditions.[3] During the course of human evolution, as much as 40% of the human genome has moved around via methods such as transposition of transposons.[2]
Transposases are classified under EC number EC 2.7.7.
Genes encoding transposases are widespread in the genomes of most organisms and are the most abundant genes known.[4]
Transposase Tn5
| Transposase Tn5 dimerisation domain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
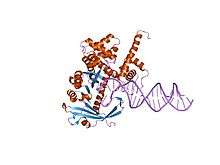 tn5 transposase: 20mer outside end 2 mn complex | |||||||||
| Identifiers | |||||||||
| Symbol | Dimer_Tnp_Tn5 | ||||||||
| Pfam | PF02281 | ||||||||
| InterPro | IPR003201 | ||||||||
| SCOPe | 1b7e / SUPFAM | ||||||||
| |||||||||
Transposase (Tnp) Tn5 is a member of the RNase superfamily of proteins which includes retroviral integrases. Tn5 can be found in Shewanella and Escherichia bacteria.[5] The transposon codes for antibiotic resistance to kanamycin and other aminoglycoside antibiotics.[3][6]
Tn5 and other transposases are notably inactive. Because DNA transposition events are inherently mutagenic, the low activity of transposases is necessary to reduce the risk of causing a fatal mutation in the host, and thus eliminating the transposable element. One of the reasons Tn5 is so unreactive is because the N- and C-termini are located in relatively close proximity to one another and tend to inhibit each other. This was elucidated by the characterization of several mutations which resulted in hyperactive forms of transposases. One such mutation, L372P, is a mutation of amino acid 372 in the Tn5 transposase. This amino acid is generally a leucine residue in the middle of an alpha helix. When this leucine is replaced with a proline residue the alpha helix is broken, introducing a conformational change to the C-Terminal domain, separating it from the N-Terminal domain enough to promote higher activity of the protein.[3] The transposition of a transposon often needs only three pieces: the transposon, the transposase enzyme, and the target DNA for the insertion of the transposon.[3] This is the case with Tn5, which uses a cut-and-paste mechanism for moving around transposons.[3]
Tn5 and most other transposases contain a DDE motif, which is the active site that catalyzes the movement of the transposon. Aspartate-97, Aspartate-188, and Glutamate-326 make up the active site, which is a triad of acidic residues.[7] The DDE motif is said to coordinate divalent metal ions, most often magnesium and manganese, which are important in the catalytic reaction.[7] Because transposase is incredibly inactive, the DDE region is mutated so that the transposase becomes hyperactive and catalyzes the movement of the transposon.[7] The glutamate is transformed into an aspartate and the two aspartates into glutamates.[7] Through this mutation, the study of Tn5 becomes possible, but some steps in the catalytic process are lost as a result.[3]
There are several steps which catalyze the movement of the transposon, including Tnp binding, synapsis (the creation of a synaptic complex), cleavage, target capture, and strand transfer. Transposase then binds to the DNA strand and creates a clamp over the transposon end of the DNA and inserts into the active site. Once the transposase binds to the transposon, it produces a synaptic complex in which two transposases are bound in a cis/trans relationship with the transposon.[3]
In cleavage, the magnesium ions activate oxygen from water molecules and expose them to nucleophilic attack.[6] This allows the water molecules to nick the 3' strands on both ends and create a hairpin formation, which separates the transposon from the donor DNA.[3] Next, the transposase moves the transposon to a suitable location. Not much is known about the target capture, although there is a sequence bias which has not yet been determined.[3] After target capture, the transposase attacks the target DNA nine base pairs apart, resulting in the integration of the transposon into the target DNA.[3]
As mentioned before, due to the mutations of the DDE, some steps of the process are lost—for example, when this experiment is performed in vitro, and SDS heat treatment denatures the transposase. However, it is still uncertain what happens to the transposase in vivo.[3]
The study of transposase Tn5 is of general importance because of its similarities to HIV-1 and other retroviral diseases. By studying Tn5, much can also be discovered about other transposases and their activities.[3]
Tn5 is utilized in genome sequencing by using the Tn5 to append sequencing adaptors and fragment the DNA in a single enzymatic reaction[8], reducing the time and input requirements over traditional Next Generation Sequencing. This means of library preparation is used in the technique called ATAC-seq and also in Illumina dye sequencing.
Sleeping Beauty transposase
The Sleeping Beauty (SB) transposase is the recombinase that drives the Sleeping Beauty transposon system.[9] SB transposase belongs to the DD[E/D] family of transposases, which in turn belong to a large superfamily of polynucleotidyl transferases that includes RNase H, RuvC Holliday resolvase, RAG proteins, and retroviral integrases.[10][11] The SB system is used primarily in vertebrate animals for gene transfer,[12] including gene therapy,[13][14] and gene discovery.[15][16] The engineered SB100X is an enzyme that directs the high levels of transposon integration.[17][18]
Tn7 transposon
The Tn7 transposon is a mobile genetic element found in many prokaryotes such as Escherichia coli (E. coli), and was first discovered as a DNA sequence in bacterial chromosomes and naturally occurring plasmids that encoded resistance to the antibiotics trimethoprim and streptomycin.[19][20] Specifically classified as a transposable element (transposon), the sequence can duplicate and move itself within a genome by utilizing a self-encoded recombinase enzyme called a transposase, resulting in effects such as creating or reversing mutations and changing genome size. The Tn7 transposon has developed two mechanisms to promote its propagation among prokaryotes.[21] Like many other bacterial transposons, Tn7 transposes at low-frequency and inserts into many different sites with little to no site-selectivity. Through this first pathway, Tn7 is preferentially directed into conjugable plasmids, which can be replicated and distributed between bacteria. However, Tn7 is unique in that it also transposes at high-frequency into a single specific site in bacterial chromosomes called attTn7.[22] This specific sequence is an essential and highly conserved gene found in many strains of bacteria. However, the recombination is not deleterious to the host bacterium as Tn7 actually transposes downstream of the gene after recognizing it, resulting in a safe way to propagate the transposon without killing the host. This highly evolved and sophisticated target-site selection pathway suggests this pathway evolved to promote coexistence between the transposon and it host, as well as Tn7's successful transmission into future generations of bacterium.[23]
The Tn7 transposon is 14 kb long and codes for five enzymes.[23] The ends of the DNA sequence consists of two segments that the Tn7 transposase interacts with during recombination. The left segment (Tn7-L) is 150 bp long and the right sequence (Tn7-R) is 90 bp long. Both ends of the transposon contain a series of 22 bp binding sites that the Tn7 transposase recognizes and binds to. Within the transposon are five discrete genes encoding for proteins that make up the transposition machinery. In addition, the transposon contains an integron, a DNA segment containing several cassettes of genes encoding for antibiotic-resistance.[23]
The Tn7 transposon codes for five proteins: TnsA, TnsB, TnsC, TnsD, and TnsE.[23] TnsA and TnsB interact together to form the Tn7 transposase enzyme TnsAB. The enzyme specifically recognizes and binds to the ends of the DNA sequence of the transposon, and excises it by introducing double-stranded DNA breaks to each end. The excised sequence is then inserted to another target DNA site. Much like other characterized transposons, the mechanism for Tn7 transposition involves cleavage of the 3' ends from the donating DNA by the TnsA protein of the TnsAB transposase. However, Tn7 is also uniquely cleaved near the 5' ends, about 5 bp from the 5' end towards the Tn7 transposon, by the TnsB protein of TnsAB. After the insertion of the transposon into the target DNA site, the 3' ends are covalently linked to the target DNA, but the 5 bp gaps are still present at the 5' ends. As a result, repair of these gaps leads to a further 5 bp duplication at the target site. The TnsC protein interacts with the transposase enzyme and the target DNA to promote the excision and insertion processes. The ability of TnsC to activate the transposase depends on its interaction with a target DNA along with its appropriate targeting protein, TnsD or TnsE. The TnsD and TnsE proteins are alternative target selectors that are also DNA binding activators that promote excision and insertion of Tn7. Their ability to interact with a particular target DNA is key to the target-site selection of Tn7. The proteins TnsA, TnsB, and TnsC thus form the core machinery of Tn7: TnsA and TnsB interact together to form the transposase, while TnsC functions as a regulator of the transposase's activity, communicating between the transposase and TnsD and TnsE. When the TnsE protein interacts with the TnsABC core machinery, Tn7 preferentially directs insertions into conjugable plasmids. When the TnsD protein interacts with TnsABC, Tn7 preferentially directs insertions downstream into a single essential and highly conserved site in the bacterial chromosome. This site, attTn7, is specifically recognized by TnsD. [23]
References
- Heffron F, McCarthy BJ, Ohtsubo H, Ohtsubo E (December 1979). "DNA sequence analysis of the transposon Tn3: three genes and three sites involved in transposition of Tn3". Cell. 18 (4): 1153–63. doi:10.1016/0092-8674(79)90228-9. PMID 391406.
- Goodsell D (December 2006). "Transposase". Molecule of the Month. Protein Data Bank.
- Reznikoff WS (March 2003). "Tn5 as a model for understanding DNA transposition". Molecular Microbiology. 47 (5): 1199–206. doi:10.1046/j.1365-2958.2003.03382.x. PMID 12603728.
- Aziz, R.K., M. Breitbart and R.A. Edwards (2010). Transposases are the most abundant, most ubiquitous genes in nature. Nucleic Acids Research 38(13): 4207-4217.Aziz RK, Breitbart M, Edwards RA (July 2010). "Transposases are the most abundant, most ubiquitous genes in nature". Nucleic Acids Research. 38 (13): 4207–17. doi:10.1093/nar/gkq140. PMC 2910039. PMID 20215432.
- McDowall J. "Transposase". InterPro.
- Lovell S, Goryshin IY, Reznikoff WR, Rayment I (April 2002). "Two-metal active site binding of a Tn5 transposase synaptic complex". Nature Structural Biology. 9 (4): 278–81. doi:10.1038/nsb778. PMID 11896402.
- Peterson G, Reznikoff W (January 2003). "Tn5 transposase active site mutations suggest position of donor backbone DNA in synaptic complex". The Journal of Biological Chemistry. 278 (3): 1904–9. doi:10.1074/jbc.M208968200. PMID 12424243.
- Adey, Andrew (December 2010). "Rapid, low-input, low-bias construction of shotgun fragment libraries by high-density in vitro transposition". Genome Biology. 11 (12): R119. doi:10.1186/gb-2010-11-12-r119. PMC 3046479. PMID 21143862.
- Ivics, Z.; Hackett, P.B.; Plasterk, R.A.; Izsvak, Z. (1997). "Molecular reconstruction of Sleeping Beauty: a Tc1-like transposon from fish and its transposition in human cells". Cell. 91 (4): 501–510. doi:10.1016/s0092-8674(00)80436-5. PMID 9390559.
- Craig NL (October 1995). "Unity in transposition reactions". Science. 270 (5234): 253–4. Bibcode:1995Sci...270..253C. doi:10.1126/science.270.5234.253. PMID 7569973.
- Nesmelova IV, Hackett PB (September 2010). "DDE transposases: Structural similarity and diversity". Advanced Drug Delivery Reviews. 62 (12): 1187–95. doi:10.1016/j.addr.2010.06.006. PMC 2991504. PMID 20615441.
- Ivics Z, Izsvák Z (January 2005). "A whole lotta jumpin' goin' on: new transposon tools for vertebrate functional genomics". Trends in Genetics. 21 (1): 8–11. doi:10.1016/j.tig.2004.11.008. PMID 15680506.
- Izsvák Z, Hackett PB, Cooper LJ, Ivics Z (September 2010). "Translating Sleeping Beauty transposition into cellular therapies: victories and challenges". BioEssays. 32 (9): 756–67. doi:10.1002/bies.201000027. PMC 3971908. PMID 20652893.
- Aronovich, E.L., McIvor, R.S., and Hackett, P.B. (2011). The Sleeping Beauty transposon system – A non-viral vector for gene therapy. Hum. Mol. Genet. (in press)Aronovich EL, McIvor RS, Hackett PB (April 2011). "The Sleeping Beauty transposon system: a non-viral vector for gene therapy". Human Molecular Genetics. 20 (R1): R14-20. doi:10.1093/hmg/ddr140. PMC 3095056. PMID 21459777.
- Carlson CM, Largaespada DA (July 2005). "Insertional mutagenesis in mice: new perspectives and tools". Nature Reviews Genetics. 6 (7): 568–80. doi:10.1038/nrg1638. PMID 15995698.
- Copeland NG, Jenkins NA (October 2010). "Harnessing transposons for cancer gene discovery". Nature Reviews. Cancer. 10 (10): 696–706. doi:10.1038/nrc2916. PMID 20844553.
- Mátés L, Chuah MK, Belay E, Jerchow B, Manoj N, Acosta-Sanchez A, et al. (June 2009). "Molecular evolution of a novel hyperactive Sleeping Beauty transposase enables robust stable gene transfer in vertebrates". Nature Genetics. 41 (6): 753–61. doi:10.1038/ng.343. PMID 19412179.
- Grabundzija I, Irgang M, Mátés L, Belay E, Matrai J, Gogol-Döring A, Kawakami K, Chen W, Ruiz P, Chuah MK, VandenDriessche T, Izsvák Z, Ivics Z (June 2010). "Comparative analysis of transposable element vector systems in human cells". Molecular Therapy. 18 (6): 1200–9. doi:10.1038/mt.2010.47. PMC 2889740. PMID 20372108.
- Barth PT, Datta N, Hedges RW, Grinter NJ (March 1976). "Transposition of a DNA sequence encoding trimethoprim and streptomycin resistances from R483 to other replicons". Journal of Bacteriology. 125 (3): 800–810. doi:10.1128/JB.125.3.800-810.1976. PMC 236152. PMID 767328.
- Barth PT, Datta N (Sep 1977). "Two Naturally Occurring Transposons Indistinguishable from Tn7". Journal of General Microbiology. 102 (1): 129–134. doi:10.1099/00221287-102-1-129. PMID 915473.
- Peters J, Craig NL (Nov 2001). "Tn7: smarter than we thought". Nature Reviews Molecular Cell Biology. 2 (11): 806–814. doi:10.1038/35099006. PMID 11715047.
- Gringauz E, Orle KA, Waddell CS, Craig NL (Jun 1988). "Recognition of Escherichia coli attTn7 by transposon Tn7: lack of specific sequence requirements at the point of Tn7 insertion". Journal of Bacteriology. 170 (6): 2832–2840. doi:10.1128/jb.170.6.2832-2840.1988. PMC 211210. PMID 2836374.
- Peters J, Craig NL (Nov 2001). "Tn7: smarter than we thought". Nature Reviews Molecular Cell Biology. 2 (11): 806–814. doi:10.1038/35099006. PMID 11715047.
External links
- Transposases at the US National Library of Medicine Medical Subject Headings (MeSH)
- Overview of all the structural information available in the PDB for UniProt: Q46731 (Transposase for transposon Tn5) at the PDBe-KB.