Timeline of mosasaur research
This timeline of mosasaur research is a chronologically ordered list of important fossil discoveries, controversies of interpretation, and taxonomic revisions of mosasaurs, a group of giant marine lizards that lived during the Late Cretaceous Epoch. Although mosasaurs went extinct millions of years before humans evolved, humans have coexisted with mosasaur fossils for millennia. Before the development of paleontology as a formal science, these remains would have been interpreted through a mythological lens. Myths about warfare between serpentine water monsters and aerial thunderbirds told by the Native Americans of the modern western United States may have been influenced by observations of mosasaur fossils and their co-occurrence with creatures like Pteranodon and Hesperornis.[1]

The scientific study of mosasaurs began in the late 18th century with the serendipitous discovery of a large fossilized skeleton in a limestone mine near Maastricht in the Netherlands.[2] The fossils were studied by local scholar Adriaan Gilles Camper, who noted a resemblance to modern monitor lizards in correspondence with renowned French anatomist Georges Cuvier.[3] Nevertheless, the animal was not scientifically described until the English Reverend William Daniel Conybeare named it Mosasaurus, after the river Meuse located near the site of its discovery.[4]
By this time the first mosasaur fossils from the United States were discovered by the Lewis and Clark expedition, and the first remains in the country to be scientifically described were reported slightly later from New Jersey.[5] This was followed by an avalanche of discoveries by the feuding Bone War paleontologists Edward Drinker Cope and Othniel Charles Marsh in the Smoky Hill Chalk of Kansas.[6] By the end of the century a specimen of Tylosaurus would be found that preserved its scaley skin.[7] Later Samuel Wendell Williston mistook fossilized tracheal rings for the remains of a fringe of skin running down the animal's back, which subsequently became a common inaccuracy in artistic restorations.[8]
The 20th century soon saw the discovery in Alabama of a strange mosasaur called Globidens, with rounded teeth suited to crushing shells.[9] Mosasaur remains were also discovered in Africa and California.[10] In 1967 Dale Russell published a scientific monograph dedicated to mosasaurs.[11] Embryonic remains in the 1990s confirmed that mosasaurs gave live birth like in ichthyosaurs.[12] The 1990s also saw a revival and escalation of a debate regarding whether or not some supposed mosasaur toothmarks in ammonoid shells were actually made by limpets.[13] By the end of the century, the evolutionary relationship between mosasaurs and snakes as well as the possible involvement of mosasaurs in the extinction of the aforementioned ichthyosaurs became hot button controversies.[14]
The debates regarding snakes, toothmarks, and ichthyosaurs spilled over into the early 21st century. These discussions were also accompanied by the discovery of many new taxa, including new species of Globidens, Mosasaurus, and Tylosaurus as well as entirely new genera like Yaguarasaurus and Tethysaurus.[15] In 2013, Lindgren, Kaddumi, and Polcyn reported the discovery of a Prognathodon specimen from Jordan that preserved the soft tissues of its scaley skin, flippers and tail. Significantly, the tail resembled those of modern carcharinid sharks, although the bottom lobe of the tail fin was longest in the mosasaur whereas shark tails have longer upper lobes.[16]
Prescientific

- One legend told among the Delaware Indians describes the discovery of a fragment of bone left by a monster that had once been killing people near modern Philadelphia in either Pennsylvania or eastern New Jersey. When the hunting party brought the piece of bone back to the village, a wise man encouraged them to set out and find more of the monsters remains. He said that smoking fragments of the bone with tobacco in a small clay spoon could grant wishes like good health for one's children, longevity, or successful hunting.[17] Fossils of the mosasaur Tylosaurus have been found in the area, but dinosaurs and Cretaceous crocodiles are also possible sources.[18] This legend likely predates European contact and may have originated prior to 1500. The bones being burnt in the "clay spoon" is a reference to a primitive kind of clay pipe that the local people had abandoned in favor of a design with a deeper bowl by the 17th century. The general lack of any sign of influence by European culture on the tale is also suggestive of its great antiquity.[19]
- The Cheyenne people of Nebraska believed in mythical thunderbirds and water monsters that were in endless conflict with each other. The thunderbirds were said to resemble giant eagles and killed both people and animals with arrows made of lightning.[20] The fossils of the Niobrara Chalk may have been influential on these stories. The pterosaur Pteranodon and marine reptiles like mosasaurs are preserved in Niobrara Chalk deposits and associated remains may have been interpreted as evidence for antagonism between immense flying animals and serpentine aquatic reptiles. Observations of similar fossils in the past may have been seen as further evidence for thunderbird-water monster conflict.[21]
The Cheyenne believe that there were many different kinds of water monsters that lived not only in lakes, rivers, and springs but also high bluffs and hills. The locations given as water monster habitat are similar to the locations where local marine fossils can be found as fossils often erode out of hillsides or stream banks. The Cheyennes feared the water monsters, because they could be dangerous predators or capsize their canoes. Even in modern times, tradition-minded Cheyenne sometimes take pains to avoid sleeping too close to springs due to fears of water monsters.[21]
- The Crow people also believed in water monsters. These were said to be alligator-like creatures living in the Little Bighorn, Rosebud, and Tongue Rivers. These myths may have been based on discoveries of marine reptile fossils such as mosasaurs and crocodilians eroding out of the river banks.[22] The Crow also believe there were similar alligator-like monsters in the Missouri River. These were called Bulukse'e or Buruksam Wurukce, names meaning "large meat-eater" but commonly translated as "giant lizard or alligator".[23]
- When the Sioux people lived around the Great Lakes, they imagined their mythical Water Monster Unktehi as a large aquatic buffalo-like mammal. This image was likely derived from early observations of large Pleistocene mammal fossils such as mammoths and mastodons eroding out of the banks of local lakes and rivers.[24] As some groups of Sioux began moving west into the regions that includes Montana their depictions of Unktehi tended to converge on the characteristics of local fossils. Although Unktehi continued to be described as horned, it gradually became imagined as reptilian rather than the mammalian portrayals of the Sioux in the Great Lakes region, like the dinosaurs and mosasaurs of the region's Mesozoic rock. Unktehi was described as a snakelike monster equipped with feet, like the elongate sinuous mosasaurs who had four short limbs. Its back was described as ridged and saw-like, a configuration similar to the appearance of a fossil vertebral column eroding from rock.[25]
18th century

1760s
1764
- The first documented mosasaurid skull is found in the chalk quarries of the St Pietersberg near Maastricht, the Netherlands.[26]
1766
- The skull was acquired by lieutenant Jean Baptiste Drouin.[26]
1770s
1772
- Around 1772, limestone miners discovered a second skull of a bizarre animal deep underground near Maastricht. The owner of the land containing the mine, canon Theodorus Joannes Godding, displayed it in his country house. Army surgeon John Leonard Hoffman observed the fossil, corresponding with various naturalists about it.[27]
1780s
- Drouin sold the first St Pietersberg mosasaur skull to the Teylers Museum at Haarlem where it is still curated today under the catalogue number TM 7424.[26]
1790s

1795
- The French revolutionary army took over both Maastricht and Goddin's astonishing fossil. The French misplaced the fossil for a period, before rediscovering it and sending it to Paris for further study.[2]
1798
- Adriaan Gilles Camper recognized that the unusual animal resembled a giant monitor lizard, and wrote to Georges Cuvier in Paris stating such.[3]
1799
- Camper continued his correspondence with Cuvier and reiterated his opinion that the remains from Maastricht belong to a giant monitor lizard.[28]
19th century

1800s
1804
September 10th
- The Lewis and Clark expedition reportedly found the skeleton of a gigantic snake in an island on the Missouri River in territory now belonging to the state of South Dakota. These remains actually probably belong to a mosasaur, possibly Mosasaurus missouriensis. Some of the bones were sent back to Washington, D.C..[29]
1810s
1818
- Mitchell reported the discovery of mosasaur fossils in the Navesink Formation of New Jersey. These were the first North American mosasaur remains to be documented in the scientific literature.[30]
1820s
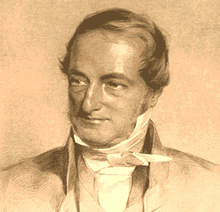
- Samuel von Sömmerring described the new species Lacerta gigantea for fossils discovered in a Bavarian iron mine, mistakenly believing them to be additional remains of the same kind of animal that was discovered in Maastricht. These remains belonged to an animal that was actually more closely related to modern crocodiles and is now known as Geosaurus.[31]
- Reverend William Daniel Conybeare formally named the animal from Maastricht Mosasaurus after a river located near the discovery site.[4]
1829
- Major Benjamin O'Fallon discovered a mosasaur in what is now South Dakota, near the Great Bend of the Missouri River. He brought it home to St. Louis and later sold it to Prince Maximilian of Wied.[11]
- Gideon Mantell applied the species epithet "hoffmanni" to the Mosasaurus type species.[30]
1830s
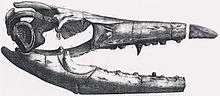
- Richard Harlan came into possession of the snout of O'Fallon's mosasaur, but mistook it for an ichthyosaur and described it at Ichthyosaurus missouriensis.[11]
- "Ichthyosaurus" missouriensis was "officially" reclassified as Mosasaurus missouriensis.[11]
1840s
- August Goldfuss described the new species Mosasaurus maximiliani for the mosasaur, naming it after the Prince.[11]
1850s

- Robert Gibbes reported the presence of teeth and vertebrae now recognized as mosasaurian in origin from a limestone deposit in Alabama.[32]
1853
- Paul Gervaius creates the family Mosasauridae. Originally included genera are Mosasaurus, Liodon, Onchosaurus (later recognised as a batoid fish), Oplosaurus (later recognised as a sauropod dinosaur), Macrosaurus (a historical wastebasket taxon) and Geosaurus (later recognised as a thalattosuchian).[33]
1860s

- Edward Drinker Cope described a variety of new mosasaur taxa from Kansas, including species in the genera Clidastes, Liodon, Mosasaurus, and Platecarpus.[34]
- Cope described the species Macrosaurus proriger.[35] This was the first mosasaur to be reported from Kansas.[36]
- Cope erected the new order Pythonomorpha in which to classify the mosasaurs because of shared traits with modern snakes.[34] He also described the new species Clidastes propython from Alabama.[32]
- Othniel Charles Marsh described the species Halisaurus platyspondylus.[37]
1870s

- Marsh led an expedition into Kansas on behalf of Yale University.[36]
November
- Marsh's crew discovered the first mosasaur specimen to preserve its hindlimbs in Kansas.[36]
- Marsh published a preliminary report about his fossil discoveries in Kansas.[36]
- Betts published the first mention of a nearly complete Clidastes specimen discovered by Marsh's crew in Kansas.[36]
- Marsh described his Kansan mosasaur discoveries in more detail, including the aforementioned Clidastes.[36]
- Marsh erroneously reported the presence of "dermal scutes" in mosasaurs.[38]
- Marsh renamed Macrosaurus proriger Rhinosaurus proriger.[35]
- Marsh renamed Rhinosaurus proriger Tylosaurus proriger.[35]

- Cope described the new species Clidastes planifrons and Tylosaurus nepaeolicus.[39]
- Francis H. Snow reported the first skin impressions of any mosasaur preserved with the Kansan Tylosaurus specimen KUVP 1075. The impressions revealed the presence and shape of its keeled, diamond-shaped scales.[7]
1880s

- Louis Dollo described the new species Hainosaurus bernardi.[40]
- Dollo described the genus Phosphorosaurus.[41]
1890s

- Samuel Wendell Williston found that the supposed dermal scutes reported in mosasaurs by Marsh were actually sclerotic ring bones.[38]
- Georg Baur published a paper on mosasaurs.[42]
- John C. Merriam described the new species Platecarpus clidastoides.[43]
- Between 1880 and this date as many as 52 partial-to-complete Mosasaurus skeletons were found in the Ciply Phosphatic Chalk near Ciply, Belgium.[44]


- Williston published notable early observations on mosasaur biostratigraphy.[45] He reported the first known mosasaur fossils from the Pteranodon beds and the apparent absence of Clidastes from the Rudistes beds of the lower chalk.[46]
- Williston published a review of paleontology's understanding of mosasaurs.[47] He bemoaned the taxonomic confusion caused by Cope and Marsh because their Bone War race for the honor of naming as many species as possible produced a large number of spurious taxa. Williston considered as many as four out of five named mosasaur species named at the time to be completely worthless.[48] He noted that some of the differences used by Cope and Marsh to distinguish their putative mosasaur species resulted from distortion by geologic forces rather than biological variation.[42] One notable taxonomic revision Williston made in this publication was his referral of Cope's 1874 species "Clidastes" planifrons to the genus Platecarpus.[49]
Williston also discussed mosasaur life appearance and behavior. He compared mosasaur scales to those of monitor lizards and misinterpreted some fossilized cartilage as evidence for a fringe of soft tissue down mosasaurs' backs.[50] He hypothesized that mosasaurs left the safety of the water to lay eggs on land.[51] The paleobiology of individual mosasaur taxa also received Williston's attention. He thought the genus Clidastes as an inhabitant of near-surface waters.[52] In contrast, Williston interpreted Platecarpus as a "deep diver" and thought Tylosaurus fed primarily on other marine reptiles.[53]
He also elaborated on his earlier research into mosasaur biostratigraphy. He observed that Tylosaurus ranged throughout the Smoky Hill Chalk but went extinct around the time the Fort Pierre Shale was deposited. Platecarpus was also only known from the Niobrara Formation. Clidastes was only known from the upper Niobrara.[54]
- Henry Fairfield Osborn recognized fossilized cartilage in a Tylosaurus specimen as the remains of tracheal rings.[38]
- Williston first reported the mosasaur specimen now catalogued as KUVP 1001 and classified in the species Platecarpus tympaniticus to the scientific literature.[38]
- Osborn reported the first evidence for a bend in the mosasaur tail in Tylosaurus proriger.[55]
20th century


1900s
- Williston publicly acknowledged that the supposed fringe running down the back of a mosasaur he had reported was actually misidentified tracheal rings.[12]
- Dollo reported the presence of the remains of a large turtle preserved in the stomach of Hainosaurus.[56]
- Fossil hunters Dennis Halvorson and Mike Hanson discovered the largest known specimen of Plioplatecarpus in the Pierre Shale of North Dakota. The skeleton was about 70% complete.[57]
- The excavation of the North Dakota Plioplatecarpus specimen concluded.[57]
1910s
_(14586414078).jpg)

- The "Bunker" Tylosaurus specimen was discovered in Kansas. At almost 45 feet in length it was one of the largest specimens ever classified in the genus and is the largest mosasaur skeleton in the United States to have been mounted for a museum exhibit, which is on display at the University of Kansas museum.[45]
- Charles Whitney Gilmore described the new genus and species Globidens alabamaensis.[32] This unusual mosasaur had rounded teeth specialized for cracking shell and probably fed on the abundant ammonites of the Cretaceous seas.[58]
- Robert Broom described a new Tylosaurus species, T. capensis from Pondoland, South Africa.[59]
- Williston interpreted the mosasaur Clidastes as inhabiting the waters near the surface of its marine home.[60]
- Charles Sternberg discovered a skeleton left behind by a 25 foot long Tylosaurus in Kansas with preserved skin. He observed that the skin was "covered with small, overlapping scales".[61]
- Charles and Levi Sternberg excavated a Platecarpus and a Clidastes sternbergii specimen in Kansas, which were sold to the Paleontological Museum in Uppsala, Sweden.[48]
- A Tylosaurus specimen was discovered in Logan County, Kansas.[62]
1920s
- Carl Wiman described the new species Clidastes sternbergii.[49] He noticed evidence for tail bends in the species Halisaurus sternbergi and Platecarpus tympaniticus. He compared them to the tail bend in ichthyosaurs that support their tail flukes.[63] In one figure, he restored the mosasaur's tail with an upper lobe like ichthyosaurs have.[63] This prescient observation would be confirmed in the early 21st century.[64]
- Sternberg reported the remains of a young polycotylid plesiosaur preserved in the remains of the 1918 Logan County Tylosaurus specimen.[62]
1930s
- Otto Zdansky reported that eight species of mosasaurs had been discovered in Egypt and ten species were known from Africa overall.[59]
- A high school student named Allan Bennison discovered the future type specimen of Plotosaurus bennisoni in Moreno Formation rocks of the San Joaquin Valley in California.[65]
- Another major mosasaur discovery occurred in California when a rancher named Frank Paiva and a Fresno State College professor named William M. Tucker discovered the future type specimen of Plotosaurus tuckeri.[66]
- Robert Leard discovered the future type specimen of Plesiotylosaurus crassidens in Moreno Formation rocks of the Panoche Hills in California.[67]
1940s

- Charles Camp described the new genera Kolposaurus and Plesiotylosaurus, although the genus Kolposaurus had already been used for a different kind of animal and Camp would later rename the mosasaur Plotosaurus. Along with describing the genus itself, Camp named two "Kolposaurus", "K." bennisoni and "K." tuckeri.[68] At least one specimen of these species was found to have fish preserved in its stomach.[62] Camp observed that Plotosaurus was a very evolutionarily advanced mosasaur with an especially stiff torso and powerful tail.[69] Plesiotylosaurus Camp found to be very similar to the original Tylosaurus.[70]
- George Gaylord Simpson reported that the possible mosasaur fossils discovered by the Lewis and Clark expedition had been lost.[71]
- Rainer Zangerl of the Chicago Field Museum led an expedition to collect fossils from the Selma Formation of Alabama and acquired several mosasaur specimens.[32]
1950s
1960s

- Erle Kauffman and Robert Kesling reported a fossil ammonite of the genus Placenticeras exhibiting damage they interpreted as tooth marks from a mosasaur, probably a platecarpine.[72]
- George Sternberg discovered a mostly complete mosasaur specimen with skin impressions near WaKeeney, Kansas. The specimen is now recognized as the best preserved specimen of the genus Ectenosaurus.[43]
Dale Russell published a monograph on American mosasaurs.[11] This monograph contained significant taxonomic revisions for the family. Among these were his conclusion that Platecarpus coryphaeus and Platecarpus ictericus were likely junior synonyms of P. tympaniticus while P. planfirons was too poorly preserved to be classified confidently at all.[73] Russell also dismissed the species Tylosaurus dyspelor described by Cope as a dubious probable-synonym of Tylosaurus proriger.[74]
Russell also published additional information and speculation on mosasaur biostratigraphy. He reported the presence of mosasaur vertebrae up to 90 million years old among the Turonian fossils curated by the University of Kansas Museum of Natural History.[75] Another of this work's significant contributions to mosasaur biostratigraphy concerned the mosasaurs of the Smoky Hill Chalk. Russell devised two biostratigraphic zones based on the Chalk's mosasaur fossils. The lower zone was characterized by Clidastes liodontus, Platecarpus coryphaeus, and Tylosaurus nepaeolicus. The upper was home to Clidastes, Platecarpus ictericus, and Tylosaurus proriger.[54] He regarded Platecarpus and Tylosaurus as deep water animals but concluded that the biostratigraphic evidence from the Smoky Hill mosasaurs suggested that the Chalk's depositional environment was becoming shallower and nearer to the ancient coastline over time.[76] Russell also argued that by the end of the Late Cretaceous, mosasaurs were converging on the body plan that characterized the first ichthyosaurs during the Triassic period and gradually replacing these older marine reptiles.[77]
- A University of Florence team embarked on a fossil hunting expedition to Nigeria. This expedition would discover a large quantity of mosasaur fossils in the Dukamaje Formation.[78]
1970s
- Russell published a paper on the mosasaurs of the Selma Formation, including those collected by the Zangerl expedition in 1945.[32] The paper contained several taxonomic advances including the description of the new species Tylosaurus zangerli and the reclassification of "Clidastes" sternbergii in the genus Halisaurus.[79] Russell considered the Mosasaurus species M. lemonnieri and M. conodon to be synonyms.[80] Overall he concluded that the known mosasaurs of the Selma Formation were Clidastes propython, Halisaurus sternbergi, Globidens alabamaensis, Platecarpus, Prognathodon, and T. zangerli.[32] He noted that the biostratigraphic distribution of mosasaurs in the Selma Formation were different than the mosasaurs of the Smoky Hill Chalk.[54]
- The University of Florence team continued searching for mosasaur fossils in Nigeria.[78]
- Azzaroli and others erected the new genus Goronyosaurus for the species "Mosasaurus" nigeriensis.[81]
- Russell claimed that unlike all other mosasaurs, Globidens dakotensis didn't have teeth on its pterygoid, although this claim has been subsequently dismissed.[82]
- Martin and Stewart argued that some fossils discovered in Cenomanian rocks in Kansas represented the oldest known mosasaur fossils.[75]
1980s
- Joan Wiffen described the new genus Moanasaurus.[40]
- Sankar Chatterjee and Zinsmeister reported the discovery of mosasaur fossils from islands off the coast of Antarctica.[83]
- Suzuki described the new species Mosasaurus hobetsuensis.[84]
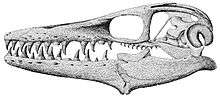
- Bell and SHeldon reported the discovery of subadult mosasaur fossils.[85]
- Judy Massare studied the anatomy of mosasaur teeth and its implication for their diet.[85] She argued that the "slender", recurved teeth of Platecarpus were adapted to feeding on relatively small, soft prey like squid.[86]
- James Martin and Phillip Bork reported the fossilized stomach contents of a Tylosaurus proriger skeleton discovered in South Dakota: a bony fish in the genus Bananogmius, Hesperornis, a lamnid shark, and Clidastes.[87]
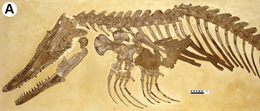
- Rothschild and Martin looked for evidence that mosasaurs suffered from the bends as a result of their diving and ascent activities. Avascular necrosis resulting from the bends was completely absent in fossils of the genus Clidastes, suggesting that it preferred to stay near the surface.[52] In contrast, Rothschild and Martin observed avascular necrosis in nearly every specimen of Platecarpus and Tylosaurus that they surveyed. The presence and ubiquity of the avascular necrosis is consistent with the idea that these were deep diving animals.[52]
- Russell described the new genus Ectenosaurus for the species "Platecarpus" clidastoides.[43]

- Kenneth Wright and Samuel Shannon described the new genus and species Selmasaurus russelli from the Mooreville Chalk near Selma, Alabama.[88]
- Lingham-Soliar reexamined Goronyosaurus from Nigeria.[89]
- Betsy Nicholls described a new species of Hainosaurus, H. pembinensis, from a Pembina Mining Company mine in the Pierre Shale of Manitoba, Canada.[90]
- Massare concluded that mosasaurs were slow swimmers that had to hunt by ambushing prey rather than chasing it down directly.[91]
- Stewart disputed Russell's 1967 mosasaur-based biostragraphic scheme for the Smoky Hill Chalk because he had observed fossils preserved outside of their purported "zones".[54]
- Martin and Rothschild interpreted Clidastes as an inhabitant of shallow waters, while Tylosaurus may have been the Cretaceous paleoecological equivalent to a sperm whale, diving deep in the Western Interior Seaway to hunt the giant squid that lived there.[92]
- Russell suggested that mosasaurs may have reproduced by laying eggs on remote island beaches.[93]
1990s

- Wiffen described the new genus Rikkisaurus.[40]
- Michael Everhart discovered two Platecarpus skulls that had been "co-mingled and partially digested".[94]
- Stewart used a recently proposed biostratigraphic scheme for the Smoky Hill Chalk to propose stratigraphic ranges for its mosasaur taxa.[54]

- Lingham-Soliar rejectd the idea that "Halisaurus" sternbergi belonged to the genus. However, he thought that the genus Phosphorosaurus should be sunk in that genus.[41] He also dismissed the supposed Mosasaurus species M. nigeriensis as a chimera of multiple mosasaur species.[59]
- Lingham-Soliar attempted to reconstruct how mosasaurs swam by studying their vertebral column. He found that the front part of column was stiffened and only the rear portion flexed during swimming. This more similar to the way a modern Alligator swims rather than snakes, which many previous researchers had put forth as mosasaur analogs.[95]
- Robert Carroll and Michael DeBraga published a discussion about aigialosaurs and their evolutionary affinities. They noted that aigialosaur skulls are virtually indistinguishable from those of primitive mosasaurs.[77] However they found the vertebrae in their back and their limb bones to more closely resemble those belonging to the precursors of living monitor lizards.[96]
- Lingham-Soliar regarded the question of how the Hainosaurus reported in 1904 by Dollo to contain a turtle in its stomach could have swallowed a prey item of that size as mysterious.[56]
- Lingham-Soliar hypothesized that Plioplatecarpus marshi "flew" underwater primarily using its powerful fore-flippers rather than using its tail like other mosasaurs due to P. marshi's well-developed shoulder region.[97] He argued that this unusual style of swimming increased its maneuverability, allowing this species to flourish in shallow habitats.[98]
- Nathalie Bardet published a discussion of the possible causes for the extinction of the ichthyosaurs. She expressed doubt that ichthyosaurs were replaced by mosasaurs because they would not be in direct competition with each other. Instead she posited a connection to an extinction event that affected cephalopods at the boundary between the Cenomanian and Turonian ages. She proposed that the disappearance of these many cephalopod species may have deprived ichthyosaurs of their food source and caused their extinction.[99]

- According to Russell, by this time at least 1,823 mosasaur specimens had been recovered from the Niobrara Chalk of Kansas alone.[74]
- Phosphate diggers in the Negev Desert of Israel discovered the remains of a 40-foot mosasaur that was sent to Copenhagen, Denmark for preparation. This mosasaur was nicknamed "Oronosaurus" after the phosphate field in which it was discovered, but would be formally named as a new species of Prognathodon, P. currii, in 2002.[100]
- Lingham-Soliar re-examined the mosasaur genus Leidon.[101]
- Schumacher published additional research on mosasaur biostratigraphy based on Stewart's 1990 paper.[83]

- Lingham-Soliar published a review of Turonian mosasaur fossils that were discovered in Angola.[102] Commenting on the evolutionary origins of the mosasaurs, he remarked that while aigialosaurs had the correct anatomy to be the ancestors of the mosasaurs they were only known from rocks of the same age as the mosasaurs themselves.[77] Until the presence of aigialosaurs earlier in the Cretaceous could be confirmed, Lingham-Soliar noted that there would be room to doubt this hypothesized connection even if it seemed plausible.[103]
- Stewart and Bell argued that the supposed oldest known mosasaur fossils reported by Martin and Stewart from Kansas were not even the remains of mosasaurs. Instead, they attributed the fossils to several kinds of ichthyodectiform fishes.[75]
- Judy Massare observed that mosasaurs exhibited adaptations for rapid acceleration and probably employed this capability as ambush hunters.[104]
- Lingham-Soliar reported the presence of abundant mosasaur remains in Angola and Zaire and observed that Africa was a hotspot of mosasaur discoveries on the same scale as western North America and the Netherlands-Belgium region. He identified four species from Zaire: Halisaurus, Mosasaurus, Plioplatecarpus, and Prognathodon. He also synonymized Angolasaurus with Platecarpus. He considered this genus to be the most widespread of any mosasaur. He interpreted Plioplatecarpus as a snake-like ratchet feeder that "walked" its jaws across large prey to swallow it. He also hypothesized that it swam using its flippers rather than relying on its tail fin.[105]
- Nicholls and Godfrey rejected Lingham-Soliar's reinterpretation of Plioplatecarpus marshi locomotion as underwater flight because there was no reason to believe that its tail was a less developed or effective means of propulsion than those of other mosasaurs. They suggested that its powerful shoulder region was actually an adaptation for shaking prey, like some modern shark species do.[106]
- Massare argued that mosasaurs were cold blooded, like their relatives the monitor lizards.[107]
January
- Patrick Antuzzi discovered a mosasaur skull in the Chico Formation of California. The specimen is now catalogued as SC-VR59 and may have belonged to a species of Clidastes resembling members of this genus previously from New Zealand.[108]
- Theagarten Lingham-Soliar published the first truly thorough description of Mosasaurus hoffmani.[4] He interpreted its preferred habitat as biodiverse coastal waters of depths from 40–50 m.[109] In this paper he also argued that mosasaurs went extinct rapidly.[110]
- Gordon Bell and Martin reported a Mosasaurus conodon skull fossil bearing bite marks apparently inflicted by another M. conodon.[94]
- Michael Everhart discovered a series of five large mosasaur vertebrae with two Cretoxyrhina teeth embedded in it. He found evidence the bone material had been partially dissolved where it had not been exposed to the elements. This implies that these vertebrae had been partially digested before fossilization by the shark that fed upon it.[94] The shark probably only scavenged the specimen, but Everhart could not rule out predation.[111]
- Pete Bussen discovered a specimen of Globidens in the Pierre Shale of Kansas. This was the first specimen of the genus to be documented in Kansas.[112]

- Gordon Bell reported the discovery of a specimen of Plioplatecarpus primaevus accompanied by two embryos of the same species. Both the remains of mother and young were accompanied more than two thousand of teeth shed by shark of the genus Squalicorax. These sharks probably only scavenged the mosasaurs and were likely responsible for the disarticulated state of the skeletons.[113] This discovery proved that mosasaurs gave live birth.[12]
- Sheldon published additional research on mosasaur biostratigraphy based on Stewart's 1990 paper.[83] One notable find was a P. planifrons fossil in the upper Smoky Hill Chalk, the uppermost known remains of this species.[83]
- Cowen disagreed with Lingham-Soliar's claim that Plioplatecarpus marshi was an underwater flyer and suggested that P. marshi's unusual traits were actually specializations for swimming near the surface of the water column by overcoming Carrier's constraint.[106]
- A man named Steve Johnson discovered a complete Platecarpus planifrons skull.[49]
- Schumacher and Varner observed more evidence for tail bends in the genera Clidastes, Platecarpus, and Tylosaurus.[63]
September
- Michael and Pam Everhart excavated the skull and neck of a mosasaur in Kansas.[114]


- Gordon Bell favorably assessed the mosasaur fossil record, finding its documentation of their evolutionary history nearly as complete as the fossil record of horses.[115]
- Bell published a detailed review of mosasaur biostratigraphy.[102]
- In contrast to Williston and Martin and Rothschild, Amy Sheldon interpreted Clidastes as a deep water diver because its bones were less dense than those of other mosasaurs, lending it neutral buoyancy during descents.[116] Meanwhile, she argued that Platecarpus preferred shallower waters.[117]
- Michael Lee argued that mosasaurs and snakes were sister groups.[118]
- Caldwell and Lee argued that the new snake genus and species Pachyrhachis problematicus was more evidence for a close evolutionary affinity between mosasaurs and snakes.[119]
- Tom Caggiano discovered a partial jaw once belonging to a young mosasaur in Kansas. This specimen had been partially digested, possibly by the shark Cretoxyrhina mantelli.[94]
- Shimada reported evidence that mosasaurs were fed upon by sharks, although this may have been scavenging rather than active predation.[120]
- Richard P. Hilton and Antuzzi reported the possible Clidastes skull SC-VR59 from California to the scientific literature.[108]


- Fossils of a mosasaur related to Tylosaurus were discovered on Vega Island, Antarctica.[121]
- Bardet and others reported the discovery of a partial Plioplatecarpus marshi near Liege, Belgium with a tail vertebra bearing grooves apparently inflicted by the shark Centrophorodes appendiculatus, which scavenged the mosasaurs remains before burial.[85]
- Ruud Dortangs discovered the skeleton of a new species of Prognathodon, P. saturator, in a Maastricht cement quarry.[80]
- Lingham-Soliar reported evidence of damage to the skull and braincase of an immature Mosasaurus hoffmanni. He observed that the area of trauma was similar in size and shape to the bony prow at the tip of the snout of a Hainosaurus. Lingham-Soliar believed that the larger Hainosaurus rammed the young Moasaurus to death with this snout prow.[122]
- Peter Ward argued that the supposed toothmarks on a Placenticeras shell described in 1960 by Kauffman and Kesling were actually limpet boreholes.[123] He described how a graduate student named Erica Roux was unable replicate tooth-shaped punctures in the shells of modern Nautilus with a mechanical "jaw" because the shells were too brittle.[124]
- Kase and others also attributed the rounded indentations and puncture marks to limpets rather than mosasaurs.[125]
- Seilacher provided a third voice in favor of the limpet interpretation of punctured ammonite shells.[125]
- Bell and VonLoh reported the discovery of mosasauroids from the Boquillas Formation of Texas and the Greenhorn Formation of South Dakota.[102]
- Lingham-Soliar reported the discovery of a mosasaur called Pluridens walkeri from Africa with unusually long, toothy jaws.[82]
August 8th
- An additional nearly complete skeleton of Mosasaurus hoffmani was discovered in the St. Pietersburg quarry of Masstricht and excavated by crews under the auspices of the Dutch Geological Society and Natural History Museum of Maastricht.[4]

- Holmes Caldwell and Cumbaa rejected the idea that Plioplatecarpus swam using its flippers to "fly" underwater. They noted however, that a specimen of the genus from Scabby Butte, Alberta, Canada seems to have been preserved in a relatively inland and freshwater environmental context, possibly an estuary.[126]
- Lingham-Soliar regarded Goronyosaurus as the most derived mosasaurid.[89] He also speculated on the ecological conditions that allowed the mosasaurs to achieve dominance of the ichthyosaurs and plesiosaurs. Lingham-Soliar observed that the evolution of swifter swimming fishes in the mid-Cretaceous would have challenged predators that actively chased them while yielding a competitive advantage to sneakier ambush hunters.[89] He thought these conditions gave an advantage to mosasaurs and plesiosaurs over the ichthyosaurs and led to the latter group's extinction.[127]
- Lingham Soliar reconstructed the heterodont dentition of Carinodens. Teeth near the tip of the jaw were pointed, teeth near the rear were roughly rectangular and triangular teeth could be found between them.[56]
- Everhart reported physical evidence of shark bite in the mosasaur genera Clidastes, Platecarpus, and Tylosaurus.[57]
- Lingham-Soliar argued that the disappearance of the mosasaurs at their peak of size and diversity was evidence that the mass extinction at the end of the Cretaceous was a rapid event.[110]
- Lee and others argued that snakes were the descendants of mosasaurs.[128]
- Lee, Bell, and Caldwell argued that mosasaurs were the closest relatives of snakes among the traditional lizards.[129]
- Polcyn, Tchernov, and Jacobs described the new genus and species Haasia gittelmani.[130]
- Sakurai and Shibuya described the new species Mosasaurus prismaticus.[131]
- Kass described the species Prognathodon stadtmani.[132]
21st century

2000s
- Holmes and Sues argued that Phosphorosaurus was distinct from Halisaurus after all.[41]
- Lingham-Soliar disagreed with Russell's 1970 synonymization of Mosasaurus lemmonnieri and M. conodon.[80]
- Tchernov and others dismissed the snakes Haasiophis and Pachyrhachis, which had been previously put forward as evidence that snakes were close relatives of mosasaurs,[119] as red herrings that were irrelevant to understanding the origin of snakes or any potential evolutionary ties linking them to mosasaurs.[133]
- Case and others published more research on mosasaurs from islands off the coast of Antarctica.[83]
- Páramo-Fonseca described the new genus and species Yaguarasaurus columbianus.[134]
_(16142481744).jpg)
- Caldwell and Lee reported the discovery of a pregnant aigialosaur specimen of the genus Carsosaurus containing four large embryos inside her.[93] The embryos were oriented ready to be born tail-first, an adaptation seen in modern whales and in other extinct marine reptiles that helps prevent drowning during birth.[135]
- Tsujita and Westermann defended the idea of punctured ammonite shells as mosasaur prey in a contrarian position to the several 1998 papers that doubted this hypothesis.[136] Based on a survey of Placenticeras shells from the Bearpaw Formation of Alberta, they concluded that the arrangements of the holes were too similar to the placement of teeth in a mosasaur's jaw to be the chance result of limpet activity. Their doubts were bolstered by the disparity of abundance in the relevant rocks between the scarce limpet shells and the frequent occurrence of the holes themselves. According to Tsujita and Westermann, the previous attempts to test the effect of a mosasaur bite on a shell experimentally with a simulated jaw assumed an overly simplistic interpretation of mosasaur feeding behavior, so its failure to replicate the shell punctures still did not discredit their putative mosasaurian origins.[137]
- Bardet and Superbiola restudied the Halisaurus remains first discovered in Kansas in 1918 and classified as Halisaurus by Russell in 1970.[138] They found it to be both the oldest specimen referred to that genus and the most primitive member of the entire Mosasauridae.[43]
- Everhart serendipitously rediscovered Sternberg's report of a Tylosaurus with polycotylid stomach contents.[62]

- Michael Everhart observed that young mosasaur fossils are actually fairly common in the Niobrara Chalk.[85]
- Caitlin Kiernan observed that different kinds of mosasaurs were found at different stratigraphic levels within the Eutaw Formation and Selma Group of Alabama. She described three biostratigraphic zones within this interval based on their mosasaur content: the Tylosaurus acme zone, the Clidastes zone, and the Mosasaurus acme zone. The Tylosaurus zone was deposited in shallow water, while Clidastes seems to have inhabited greater depths.[109]
- Bardet and Tonoglu reported the discovery of Mosasaurus hoffmanni jaw fossils in Kastamonu, Turkey.[109]
- Novas and others described the new species Lakumasaurus antarcticus.[45]
- Dortangs and others described the new species Prognathodon saturator.[139]
- Nicholls and Meckert described the new genus and species Kourisodon puntledgensis.[140]
- Thegarten Lingham-Soliar argued against Nathalie Bardet's attribution of ichthyosaur extinction to the loss of their preferred food sources in the Cenomanian-Turonian extinction event expanded on his own 1999 attribution of ichthyosaur extinction to biotic factors. During the Cretaceous the evolution of many fish groups trended towards faster swimming body types, making them harder to hunt for adult ichthyosaurs and harder to escape from for newborn ichthyosaurs. This ecological scene favored ambush predators like plesiosaurs and the newly evolved mosasaurs over the ichthyosaurs, who succumbed to the competition.[127]
September
- Christansen and Bonde formally named the Negev desert mosasaur Prognathodon currii.[100]
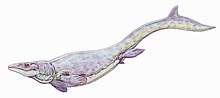
- Caldwell and others published a study on the tooth replacement patterns of mosasaurs. They found that mosasaurs replaced their teeth continuously throughout their lives, with each replacing being slightly larger than its predecessor.[62]
- Polcyn, Tchernov, and Jacobs erected the new genus Haasiasaurus to house the species "Haasia" gittelmani because its generic name had been previously used for a different kind of animal and needed replacing.[141]
- Bardet, Suberbiola, and Jalil described the new genus and species Tethysaurus nopscai.[142]

- Lindgren and Siverson argued that mosasaurs almost went extinct during the middle of the Campanian age, but subsequently recovered their lost biodivsersity.[143]
- Martin and Fox reported the discovery of crushed bivalve shells in the stomach of a Globidens specimen from the Pierre Shale of South Dakota.[62]
May
- Gordon Bell and Mike Caldwell rediscovered the mosasaur snout fragment discovered by Harlan. Using photographs of the Goldfuss mosasaur skull by Takehito Ikejiri they were able to verify that the specimens did in fact come from the same individual.[144]
- Polcyn and Bell described the new genus and species Russellosaurus coheni.[145]
- Bardet and others described the new species Halisaurus arambourgi.[146]
- Everhart described the new species Tylosaurus kansasensis.[147]
- Bell and Polcyn described the new genus and species Dallasaurus turneri.[148]
- Martin described the new species Globidens schurmanni.[149]
- Schulp and others described the new species Prognathodon kianda.[150]
- Polcyn and Everhart described the species Selmasaurus johnsoni.[151]
- Caldwell and others described the new species Taniwhasaurus mikasaensis.[152]
- Smith and Buchy described the new genus and species Vallecillosaurus donrobertoi.[153]
2010s
- Konishi and Caldwell the new genus and species Latoplatecarpus willistoni and referred the species "Plioplatecarpus" nichollsae to the same genus. They also erected a second new genus, Plesioplatecarpus for the species "Clidastes" planifrons.[156]
- Leblanc, Caldwell and Bardet described the new genus and species Eremiasaurus heterodontus.[157]
- Makádi, Caldwell and Ősi described the new genus and species Pannoniasaurus inexpectatus.[158]
- Lindgren, Kaddumi, and Polcyn reported the discovery of a Prognathodon specimen from Jordan that preserved the soft tissues of its scaley skin, flippers and tail. Significantly, the tail resembled those of modern carcharinid sharks, although the bottom lobe of the tail fin was longest in the mosasaur whereas shark tails have longer upper lobes.[16]
- Páramo described the new species Eonatator coellensis.[159]
- Palci, Caldwell and Papazzoni described the new genus Romeosaurus and the new species R. fumanensis and R. sorbinii.[160]
- Konishi and others described the new species Phosphorosaurus ponpetelegans.[161]
- Cuthbertson and Holmes described the new species Plioplatecarpus peckensis.[162]
- Longrich described the species Pluridens calabaria.[163]
- A study on mosasaur tooth implantation and its phylogenetic implications is published by Liu et al. (2016).[164]
- A redescription of the mosasaur Hainosaurus bernardi Dollo (1885) is published by Jimenez-Huidobro & Caldwell (2016), who transfer this species to the genus Tylosaurus and synonymize genera Tylosaurus and Hainosaurus.[165]
- A revision of the species assigned to the mosasaur genus Tylosaurus is published by Jiménez-Huidobro, Simões & Caldwell (2016);[166] their conclusion that T. kansasensis is a junior synonym of T. nepaeolicus is subsequently rejected by Stewart & Mallon (2018).[167]
- Otero and others described the new genus and species Kaikaifilu hervei.[168]
- A study on the phylogenetic relationships of members of Mosasauroidea is published by Simões et al. (2017).[169]
- A study on the robustness of the hypotheses about mosasauroid phylogenetic relationships and a reevaluation of the dataset from the study of Simões et al. (2017) is published by Madzia & Cau (2017).[170]
- A revision of mosasauroids from the Upper Cretaceous marine sediments associated with Gondwanan landmasses is published by Jiménez-Huidobro, Simões & Caldwell (2017).[171]
- A redescription of Mosasaurus hoffmannii based on examination of many specimens is published by Street & Caldwell (2017), who also provide emended diagnoses for both the genus Mosasaurus and its type species M. hoffmannii.[172]
- A study on the presence of ligamentous tooth attachment in mosasaurs and in fossil and modern snakes is published by LeBlanc, Lamoureux & Caldwell (2017).[173]
- Jiménez-Huidobro and others described the new species Tylosaurus saskatchewanensis.[174]
- A basal mosasauroid specimen including a rib and a vertebra, representing a larger individual than the holotype of Phosphorosaurus ponpetelegans and predating P. ponpetelegans by approximately 10 million years, is reported from the Upper Cretaceous (lower Campanian) of Hokkaido (Japan) by Sato et al. (2018).[175]
- Description of a Campanian mosasaur assemblage from the Hannover region of northern Germany is published by Hornung, Reich & Frerichs (2018), who confirm the presence of the genus Clidastes in northern central Europe, as well as the early Transatlantic distribution of a basal member of the genus Prognathodon during the lower Campanian.[176]
- Description of two skulls of subadult specimens of Tylosaurus proriger from the Niobrara Formation (Kansas, United States), and a study on the allometric changes undergone by T. proriger through life, is published by Stewart & Mallon (2018),[167] who reject the hypothesis presented by Jiménez-Huidobro, Simões & Caldwell (2016) that Tylosaurus kansasensis is a junior synonym of Tylosaurus nepaeolicus.[177]
- The smallest-known, neonate-sized specimen of Tylosaurus is described from the Santonian portion of the Niobrara Chalk (Kansas, United States) by Konishi, Jiménez-Huidobro & Caldwell (2018).[178]
- A study evaluating the fossil record of mosasaurs in terms of fossil completeness as a measure of fossil quality is published by Driscoll et al. (2019).[179]
- A study on the morphology of the marginal teeth of Mosasaurus lemonnieri, and on their implications for the distinguishability of this species on the basis of fossil teeth, will be published by Madzia (2019).[180]
- A study on the anatomy of the inner ear of Platecarpus will be published by Yi & Norell (2019).[181]
- An isolated tooth of a tylosaurine mosasaur is described from the Turonian of the Apennine Carbonate Platform by Romano et al. (2019), representing the first tylosaurine from Italy and the southernmost occurrence of a tylosaurine in the northern margin of the Mediterranean Tethys.[182]
- A study on the phylogenetic relationships of tylosaurine mosasaurs is published by Jiménez-Huidobro & Caldwell (2019).[183]
- A review of the taxonomic history of Clidastes liodontus and "Clidastes moorevillensis" is published by Lively (2019).[184]
- A juvenile mosasaur specimen affected by infectious arthritis and spondyloarthropathy is described from the upper Maastrichtian of Antarctica by Talevi et al. (2019), representing the first report of a skeletal pathology of a mosasaur from the Southern Hemisphere.[185]
- Leblanc, Mohr & Caldwell described the new species Globidens simplex.[186]
- Campbell Mekarski, Japundžić, Krizmanić & Caldwell describe a new genus from a specimen recovered in 2008 from Dugi Otok.[187]
Footnotes
- Mayor (2005); "Cheyenne Fossil Knowledge", pages 209–211.
- Ellis (2003); "The Mosasaurs", page 195.
- Ellis (2003); "The Mosasaurs", page 196.
- Ellis (2003); "The Mosasaurs", page 197.
- For the Lewis and Clark mosasaur discovery, see Everhart (2005a); "Enter the Mosasaurs", pages 172-173 and Ellis (2003); "The Mosasaurs", page 216. For the New Jersey find, see only Everhart (2005a); "Enter the Mosasaurs", pages 173.
- Everhart (2005a); "Enter the Mosasaurs", in passim.
- Everhart (2005a); "Enter the Mosasaurs", page 178.
- Everhart (2005a); "Enter the Mosasaurs", pages 178–180.
- For the date of discovery, see Ellis (2003); "The Mosasaurs", page 219. For remarks about its diet, see pages 228–229.
- For African mosasaurs, see Ellis (2003); "The Mosasaurs", page 222 and page 225. For Californian mosasaurs, see Hilton (2003); "Mosasaurs", pages 107–111.
- Ellis (2003); "The Mosasaurs", page 198.
- Everhart (2005a); "Enter the Mosasaurs", page 180.
- Ellis (2003); "The Mosasaurs", pages 235–238.
- For the controversy regarding the relationship between mosasaurs and snakes, see Ellis (2003); "The Mosasaurs", pages 245–250. For the possible role mosasaurs played in the extinction of the ichthyosaurs, see ibid.; "The Ichthyosaurs," pages 114–116 and "The Mosasaurs", page 223.
- For a recently described Globidens species, G. schurmanni see Martin (2007); in passim. For Mosasaurus prismaticus, see Sakurai and Shibuya (1999); in passim. For Tylosaurus kansasensis, see Everhart (2005b); in passim. For the recently described genus Yaguarasaurus, see Páramo-Fonseca (2000); in passim. For Tethysaurus, see Bardet, Superbiola, and Jalil (2003); in passim.
- Lindgren, Kaddumi, and Polcyn (2013); in passim.
- Mayor (2005); "Smoking the Monster's Bone: An Ancient Delaware Fossil Legend", page 68.
- Mayor (2005); "Smoking the Monster's Bone: An Ancient Delaware Fossil Legend", page 69.
- Mayor (2005); "Smoking the Monster's Bone: An Ancient Delaware Fossil Legend", page 70.
- Mayor (2005); "Cheyenne Fossil Knowledge", pages 209–210.
- Mayor (2005); "Cheyenne Fossil Knowledge", page 211.
- Mayor (2005); "Water Monsters and Thunder Birds on the Prairie and in the Badlands", page 275.
- Mayor (2005); "Water Monsters and Thunder Birds on the Prairie and in the Badlands", pages 275–276.
- Mayor (2005); "Water Monsters and Thunder Birds on the Prairie and in the Badlands", page 235.
- Mayor (2005); "Water Monsters and Thunder Birds on the Prairie and in the Badlands", page 237.
- Mulder, E.W.A., 2004, "Maastricht Cretaceous finds and Dutch pioneers in vertebrate palaeontology". In: Touret, J.L.R. & Visser, R.P.W. (eds). Dutch pioneers of the earth sciences, pp. 165-176. Royal Netherlands Academy of Arts and Sciences (KNAW), Amsterdam.
- Rompen, P. 1995. Mosasaurus hoffmanni: De lotgevallen van een type-exemplaar. Thesis, University of Maastricht
- Camper, A.G. (1800). "Lettre de A.G. Camper à G. Cuvier sur les ossemens fossiles de la montagne de St. Pierre, à Maëstricht". Journal de Physique. 51: 278–291.
- For date see Everhart (2005a); "Enter the Mosasaurs", page 172. For other details, see Ellis (2003); "The Mosasaurs", page 216.
- Everhart (2005a); "Enter the Mosasaurs", page 172.
- Ellis (2003); "The Mosasaurs", pages 196–197.
- Ellis (2003); "The Mosasaurs", page 219.
- Russell, Dale. A. (6 November 1967). "Systematics and Morphology of American Mosasaurs" (PDF). Bulletin of the Peabody Museum of Natural History (Yale University).
- Ellis (2003); "The Mosasaurs", page 210.
- Ellis (2003); "The Mosasaurs", page 208.
- Everhart (2005a); "Enter the Mosasaurs", page 175.
- Ellis (2003); "The Mosasaurs", pages 213–214.
- Everhart (2005a); "Enter the Mosasaurs", page 179.
- For C. planifrons, see Everhart (2005a); "Enter the Mosasaurs", pages 164–165. For Tylosaurus nepaeolicus see Ellis (2003); "The Mosasaurs", page 209.
- Ellis (2003); "The Mosasaurs", page 227.
- Ellis (2003); "The Mosasaurs", page 214.
- Everhart (2005a); "Enter the Mosasaurs", page 176.
- Everhart (2005a); "Enter the Mosasaurs", page 167.
- Ellis (2003); "The Mosasaurs", pages 220–221.
- Everhart (2005a); "Enter the Mosasaurs", page 169.
- Everhart (2005a); "Enter the Mosasaurs", pages 169–170.
- Ellis (2003); "The Mosasaurs", page 211.
- Ellis (2003); "The Mosasaurs", page 213.
- Everhart (2005a); "Enter the Mosasaurs", page 165.
- Everhart (2005a); "Enter the Mosasaurs", pages 178–179.
- Everhart (2005a); "Enter the Mosasaurs", page 181.
- Ellis (2003); "The Mosasaurs", page 239.
- Ellis (2003); "The Mosasaurs", pages 239–240.
- Everhart (2005a); "Enter the Mosasaurs", page 170.
- Everhart (2005a); "Enter the Mosasaurs", pages 176–177.
- Ellis (2003); "The Mosasaurs", page 229.
- Ellis (2003); "The Mosasaurs", page 242.
- Ellis (2003); "The Mosasaurs", pages 228–229.
- Ellis (2003); "The Mosasaurs", page 222.
- Ellis (2003); "The Mosasaurs", page 205.
- Ellis (2003); "The Mosasaurs", page 206.
- Everhart (2005a); "Enter the Mosasaurs", page 182.
- Everhart (2005a); "Enter the Mosasaurs", page 177.
- Lindgren, Kaddumi, and Polcyn (2013); "Introduction", page 2.
- Hilton (2003); "Mosasaurs", page 107.
- Hilton (2003); "Mosasaurs", page 108.
- Hilton (2003); "Mosasaurs", page 110.
- Hilton (2003); "Mosasaurs", pages 107–111.
- Hilton (2003); "Mosasaurs", pages 109–110.
- Hilton (2003); "Mosasaurs", pages 110–111.
- Everhart (2005a); "Enter the Mosasaurs", page 173.
- Ellis (2003); "The Mosasaurs", page 235.
- For P. coryphaeus and P. ictericus as synonyms of P. tympaniticus, see Everhart (2005); "Enter the Mosasaurs", page 171. For the dubious nature of P. planifrons, see page 165.
- Ellis (2003); "The Mosasaurs", page 207.
- Ellis (2003); "The Mosasaurs", page 203.
- For Russell's interpretation of Platecarpus and Tylosaurus as deep water animals, see Ellis (2003); "The Mosasaurs", page 239. For the decreasing depth of the Western Interior Seaway as implied by mosasaur fossils, see Everhart (2005a); "Enter the Mosasaurs", page 170.
- Ellis (2003); "The Mosasaurs", page 200.
- Ellis (2003); "The Mosasaurs", pages 222–223.
- For the description of Tylosaurus zangerli, see Ellis (2003); "The Mosasaurs", page 219. For the reclassification of "Clidastes" sternbergii to Halisaurus, see Everhart (2005a); "Enter the Mosasaurs", page 165.
- Ellis (2003); "The Mosasaurs", page 221.
- Azzaroli, de Guili, and Torre (1972); in passim.
- Everhart (2005a); "Enter the Mosasaurs", page 184.
- Everhart (2005a); "Enter the Mosasaurs", page 171.
- Suzuki (1985); in passim.
- Ellis (2003); "The Mosasaurs", page 218.
- Everhart (2005a); "Enter the Mosasaurs", page 168.
- Ellis (2003); "The Mosasaurs", page 228. For the identity of the bony fish as Bananogmius, see Everhart (2005a); "Enter the Mosasaurs", page 182.
- Ellis (2003); "The Mosasaurs", pages 216–217.
- Ellis (2003); "The Mosasaurs", page 223.
- Ellis (2003); "The Mosasaurs", page 230.
- Ellis (2003); "The Mosasaurs", page 232. Everhart (2005a); "Enter the Mosasaurs", page 176.
- For Clidastes as a shallow water animal, see Ellis (2003); "The Mosasaurs", page 205. For Tylosaurus as a sperm whale analogue, see page 240.
- Ellis (2003); "The Mosasaurs", page 217.
- Everhart (2005a); "Enter the Mosasaurs", page 159.
- Ellis (2003); "The Mosasaurs", pages 231–232.
- Ellis (2003); "The Mosasaurs", page 201.
- Ellis (2003); "The Mosasaurs", page 232.
- Ellis (2003); "The Mosasaurs", pages 232–233.
- Ellis (2003); "The Ichthyosaurs," page 114.
- Ellis (2003); "The Mosasaurs", pages 225–226.
- Ellis (2003); "The Mosasaurs", pages 234–235.
- Everhart (2005a); "Enter the Mosasaurs", page 162.
- Ellis (2003); "The Mosasaurs", pages 200–201.
- Ellis (2003); "The Mosasaurs", page 204.
- Ellis (2003); "The Mosasaurs", page 225.
- Ellis (2003); "The Mosasaurs", page 233.
- Ellis (2003); "The Mosasaurs", pages 238–239.
- Hilton (2003); "Mosasaurs", page 111.
- Ellis (2003); "The Mosasaurs", page 220.
- Ellis (2003); "The Mosasaurs", page 244.
- Everhart (2005a); "Enter the Mosasaurs", pages 159–160.
- Everhart (2005a); "Figure 9.10", page 184.
- Ellis (2003); "The Mosasaurs", page 217. Note that the number of embryos given by Ellis may not be correct, see Everhart (2005a); "Enter the Mosasaurs", page 159.
- Ellis (2003); "The Mosasaurs", pages 218–219.
- Ellis (2003); "The Mosasaurs", page 202.
- Ellis (2003); "The Mosasaurs", pages 205–206.
- Ellis (2003); "The Mosasaurs", page 240.
- Ellis (2003); "The Mosasaurs", page 245.
- Ellis (2003); "The Mosasaurs", pages 245–246.
- Everhart (2005a); "Enter the Mosasaurs", pages 185–186.
- Ellis (2003); "The Mosasaurs", page 215.
- Ellis (2003); "The Mosasaurs", pages 230–231.
- Ellis (2003); "The Mosasaurs", page 236.
- Ellis (2003); "The Mosasaurs", pages 236–237.
- Ellis (2003); "The Mosasaurs", page 237.
- Ellis (2003); "The Mosasaurs", page 216.
- Ellis (2003); "The Ichthyosaurs," page 115.
- Ellis (2003); "The Mosasaurs", page 248.
- Ellis (2003); "The Mosasaurs", pages 248–249.
- Polcyn, Tchernov, and Jacobs (1999); in passim.
- Sakurai and Shibuya (1999); in passim.
- Kass (1999); in passim.
- Ellis (2003); "The Mosasaurs", page 247.
- Páramo-Fonseca (2000); in passim.
- Ellis (2003); "The Mosasaurs", pages 217–218.
- Ellis (2003); "The Mosasaurs", pages 237–238.
- Ellis (2003); "The Mosasaurs", page 238.
- Everhart (2005a); "Enter the Mosasaurs", pages 165–167.
- Dortangs et al. (2002); in passim.
- Nicholls and Meckert (2002); in passim.
- Polcyn, Tchernov, and Jacobs (2003); in passim.
- Bardet, Superbiola, and Jalil (2003); in passim.
- Everhart (2005a); "Enter the Mosasaurs", page 163.
- Everhart (2005a); "Enter the Mosasaurs", page 174.
- Polcyn and Bell (2005); in passim.
- Bardet et al. (2005); in passim.
- Everhart (2005b); in passim.
- Bell and Polcyn (2005); in passim.
- Martin (2007); in passim.
- Schulp et al. (2008); in passim.
- Polcyn and Everhart (2008); in passim.
- Caldwell, M.W.; et al. (2008). "A New Species Of Taniwhasaurus (Mosasauridae, Tylosaurinae) From The Upper Santonian-Lower Campanian (Upper Cretaceous) Of Hokkaido, Japan". Journal of Vertebrate Paleontology. 28 (2): 339–348. doi:10.1671/0272-4634(2008)28[339:ANSOTM]2.0.CO;2. ISSN 0272-4634.
- Krister T. Smith; Marie-Céline Buchy (2008). "A new aigialosaur (Squamata: Anguimorpha) with soft tissue remains from the Upper Cretaceous of Nuevo León, Mexico". Journal of Vertebrate Paleontology. 28 (1): 85–94. doi:10.1671/0272-4634(2008)28[85:ANASAW]2.0.CO;2.
- A.S. Schulp; N. Bardet; B. Bouya (2009). "A new species of the durophagous mosasaur Carinodens (Squamata, Mosasauridae) and additional material of Carinodens belgicus from the Maastrichtian phosphates of Morocco". Netherlands Journal of Geosciences. 88 (3): 161–167. doi:10.1017/S0016774600000871.
- Nathalie Bardet (2012). "Maastrichtian marine reptiles of the Mediterranean Tethys: a palaeobiogeographical approach". Bulletin de la Société Géologique de France. 183 (6): 573–586. doi:10.2113/gssgfbull.183.6.573.
- Konishi, Takuya; Michael W. Caldwell (2011). "Two new plioplatecarpine (Squamata, Mosasauridae) genera from the Upper Cretaceous of North America, and a global phylogenetic analysis of plioplatecarpines". Journal of Vertebrate Paleontology. 31 (4): 754–783. doi:10.1080/02724634.2011.579023.
- Leblanc, A.R.H.; Caldwell, M.W.; Bardet, N. (2012). "A new mosasaurine from the Maastrichtian (Upper Cretaceous) phosphates of Morocco and its implications for mosasaurine systematics". Journal of Vertebrate Paleontology. 32 (1): 82–104. doi:10.1080/02724634.2012.624145.
- László Makádi; Michael W. Caldwell; Attila Ősi (2012). "The First Freshwater Mosasauroid (Upper Cretaceous, Hungary) and a New Clade of Basal Mosasauroids". PLoS ONE. 7 (12): e51781. Bibcode:2012PLoSO...751781M. doi:10.1371/journal.pone.0051781. PMC 3526648. PMID 23284766.
- Páramo-Fonseca, M. E. (2013). "Eonatator coellensis nov. sp. (Squamata: Mosasauridae), nueva especie del Cretácico Superior de Colombia" [Eonatator coellensis nov. sp. (Squamata: Mosasauridae), a new species from the Upper Cretaceous of Colombia]. Revista de la Academia Colombiana de Ciencias (in Spanish). 37 (145): 499–518. doi:10.18257/raccefyn.31. ISSN 0370-3908.
- Alessandro Palci; Michael W. Caldwell; Cesare A. Papazzoni (2013). "A new genus and subfamily of mosasaurs from the Upper Cretaceous of northern Italy". Journal of Vertebrate Paleontology. 33 (3): 599–612. doi:10.1080/02724634.2013.731024.
- Takuya Konishi; Michael W. Caldwell; Tomohiro Nishimura; Kazuhiko Sakurai; Kyo Tanoue (2015). "A new halisaurine mosasaur (Squamata: Halisaurinae) from Japan: the first record in the western Pacific realm and the first documented insights into binocular vision in mosasaurs". Journal of Systematic Palaeontology. 14 (10): 809–839. doi:10.1080/14772019.2015.1113447.
- Robin S. Cuthbertson; Robert B. Holmes (2015). "A new species of Plioplatecarpus (Mosasauridae, Plioplatecarpinae) from the Bearpaw Formation (Campanian, Upper Cretaceous) of Montana, U.S.A.". Journal of Vertebrate Paleontology. 35 (3): e922980. doi:10.1080/02724634.2014.922980.
- Nicholas R. Longrich (2016). "A new species of Pluridens (Mosasauridae: Halisaurinae) from the upper Campanian of Southern Nigeria". Cretaceous Research. 64: 36–44. doi:10.1016/j.cretres.2016.03.013.
- Min Liu; David A. Reed; Giancarlo M. Cecchini; Xuanyu Lu; Karan Ganjawalla; Carol S. Gonzales; Richard Monahan; Xianghong Luan; Thomas G. H. Diekwisch (2016). "Varanoid tooth eruption and implantation modes in a Late Cretaceous mosasaur". Frontiers in Physiology. 7: Article 145. doi:10.3389/fphys.2016.00145. PMC 4869606. PMID 27242535.
- Paulina Jimenez-Huidobro; Michael W. Caldwell (2016). "Reassessment and reassignment of the early Maastrichtian mosasaur Hainosaurus bernardi Dollo, 1885, to Tylosaurus Marsh, 1872". Journal of Vertebrate Paleontology. 36 (3): e1096275. doi:10.1080/02724634.2016.1096275.
- Paulina Jiménez-Huidobro; Tiago R. Simões; Michael W. Caldwell (2016). "Re-characterization of Tylosaurus nepaeolicus (Cope, 1874) and Tylosaurus kansasensis Everhart, 2005: Ontogeny or sympatry?". Cretaceous Research. 65: 68–81. doi:10.1016/j.cretres.2016.04.008.
- Robert F. Stewart; Jordan C. Mallon (2018). "Allometric growth in the skull of Tylosaurus proriger (Squamata: Mosasauridae) and its taxonomic implications". Vertebrate Anatomy Morphology Palaeontology. 6: 75–90. doi:10.18435/vamp29339.
- Rodrigo A. Otero; Sergio Soto-Acuña; David Rubilar-Rogers; Carolina S. Gutstein (2017). "Kaikaifilu hervei gen. et sp. nov., a new large mosasaur (Squamata, Mosasauridae) from the upper Maastrichtian of Antarctica". Cretaceous Research. 70: 209–225. doi:10.1016/j.cretres.2016.11.002.
- Tiago R. Simões; Oksana Vernygora; Ilaria Paparella; Paulina Jimenez-Huidobro; Michael W. Caldwell (2017). "Mosasauroid phylogeny under multiple phylogenetic methods provides new insights on the evolution of aquatic adaptations in the group". PLoS ONE. 12 (5): e0176773. Bibcode:2017PLoSO..1276773S. doi:10.1371/journal.pone.0176773. PMC 5415187. PMID 28467456.
- Daniel Madzia; Andrea Cau (2017). "Inferring 'weak spots' in phylogenetic trees: application to mosasauroid nomenclature". PeerJ. 5: e3782. doi:10.7717/peerj.3782. PMC 5602675. PMID 28929018.
- Paulina Jiménez-Huidobro; Tiago R. Simões; Michael W. Caldwell (2017). "Mosasauroids from Gondwanan continents". Journal of Herpetology. 51 (3): 355–364. doi:10.1670/16-017.
- Hallie P. Street & Michael W. Caldwell (2017). "Rediagnosis and redescription of Mosasaurus hoffmannii (Squamata: Mosasauridae) and an assessment of species assigned to the genus Mosasaurus". Geological Magazine. 154 (3): 521–557. Bibcode:2017GeoM..154..521S. doi:10.1017/S0016756816000236.
- Aaron R. H. LeBlanc; Denis O. Lamoureux; Michael W. Caldwell (2017). "Mosasaurs and snakes have a periodontal ligament: timing and extent of calcification, not tissue complexity, determines tooth attachment mode in reptiles". Journal of Anatomy. 231 (6): 869–885. doi:10.1111/joa.12686. PMC 5696141. PMID 28901023.
- Paulina Jiménez-Huidobro; Michael W. Caldwell; Ilaria Paparella; Timon S. Bullard (2018). "A new species of tylosaurine mosasaur from the upper Campanian Bearpaw Formation of Saskatchewan, Canada". Journal of Systematic Palaeontology. 17 (10): 849–864. doi:10.1080/14772019.2018.1471744.
- Tamaki Sato; Takuya Konishi; Tomohiro Nishimura; Takeru Yoshimura (2018). "A basal mosasauroid from the Campanian (Upper Cretaceous) of Hokkaido, northern Japan". Paleontological Research. 22 (2): 156–166. doi:10.2517/2017PR018.
- Jahn J. Hornung; Mike Reich; Udo Frerichs (2018). "A mosasaur fauna (Squamata: Mosasauridae) from the Campanian (Upper Cretaceous) of Hannover, northern Germany". Alcheringa: An Australasian Journal of Palaeontology. 42 (4): 543–559. doi:10.1080/03115518.2018.1434899.
- Paulina Jiménez-Huidobro; Tiago R. Simões; Michael W. Caldwell (2016). "Re-characterization of Tylosaurus nepaeolicus (Cope, 1874) and Tylosaurus kansasensis Everhart, 2005: Ontogeny or sympatry?". Cretaceous Research. 65: 68–81. doi:10.1016/j.cretres.2016.04.008.
- Takuya Konishi; Paulina Jiménez-Huidobro; Michael W. Caldwell (2018). "The smallest-known neonate individual of Tylosaurus (Mosasauridae, Tylosaurinae) sheds new light on the tylosaurine rostrum and heterochrony". Journal of Vertebrate Paleontology. in press (5): e1510835. doi:10.1080/02724634.2018.1510835.
- Daniel A. Driscoll; Alexander M. Dunhill; Thomas L. Stubbs; Michael J. Benton (2019). "The mosasaur fossil record through the lens of fossil completeness" (PDF). Palaeontology. 62 (1): 51–75. doi:10.1111/pala.12381.
- Daniel Madzia (2019). "Dental variability and distinguishability in Mosasaurus lemonnieri (Mosasauridae) from the Campanian and Maastrichtian of Belgium, and implications for taxonomic assessments of mosasaurid dentitions". Historical Biology: An International Journal of Paleobiology. in press: 1–15. doi:10.1080/08912963.2019.1588892.
- Hongyu Yi; Mark Norell (2019). "The bony labyrinth of Platecarpus (Squamata: Mosasauria) and aquatic adaptations in squamate reptiles". Palaeoworld. in press (4): 550–561. doi:10.1016/j.palwor.2018.12.001.
- Marco Romano; Riccardo Manni; Enrico Venditti; Umberto Nicosia; Angelo Cipriani (2019). "First occurrence of a Tylosaurinae mosasaur from the Turonian of the Central Apennines, Italy". Cretaceous Research. 96: 196–209. doi:10.1016/j.cretres.2019.01.001.
- Paulina Jiménez-Huidobro; Michael W. Caldwell (2019). "A new hypothesis of the phylogenetic relationships of the Tylosaurinae (Squamata: Mosasauroidea)". Frontiers in Earth Science. 7: Article 47. Bibcode:2019FrEaS...7...47J. doi:10.3389/feart.2019.00047.
- Joshua R. Lively (2019). "Taxonomy and historical inertia: Clidastes (Squamata: Mosasauridae) as a case study of problematic paleobiological taxonomy". Alcheringa: An Australasian Journal of Palaeontology. in press (4): 516–527. doi:10.1080/03115518.2018.1549685.
- Marianella Talevi; Bruce Rothschild; Marta Fernández; Marcelo Reguero; Matías Mitidieri (2019). "A pathological scapula in a mosasaur from the upper Maastrichtian of Antarctica: evidence of infectious arthritis and spondyloarthropathy". Cretaceous Research. 100: 1–4. doi:10.1016/j.cretres.2019.03.024.
- Aaron R. H. Leblanc; Sydney R. Mohr; Michael W. Caldwell (2019). "Insights into the anatomy and functional morphology of durophagous mosasaurines (Squamata: Mosasauridae) from a new species of Globidens from Morocco". Zoological Journal of the Linnean Society. Online edition (4): 1026–1052. doi:10.1093/zoolinnean/zlz008.
- Michelle Campbell Mekarski; Dražen Japundžić; Katarina Krizmanić; Michael W. Caldwell (2019). "Description of a new basal mosasauroid from the Late Cretaceous of Croatia, with comments on the evolution of the mosasauroid forelimb". Journal of Vertebrate Paleontology. 39: e1577872. doi:10.1080/02724634.2019.1577872.
References
- Azzaroli, A.; De Guili, C.; Torre, D. (1972). "An aberrant mosasaur from the Upper Cretaceous of north western Nigeria". Accademia Nazionale dei Lincei. Rendiconti. Classe di Scienze Fisiche Matematiche e Naturali. 52: 53–56.
- Bardet, N.; Suberbiola, X. P.; Jalil, N-E (2003). "A new mosasauroid (Squamata) from the Late Cretaceous (Turonian) of Morocco". Comptes Rendus Palevol. 2 (8): 607–616. doi:10.1016/j.crpv.2003.09.006.
- Bardet, N.; Suberbiola, X. P.; Iarochene, M.; Bouyahyaoui, F.; Bouya, B.; Amaghzaz, M. (2005). "A new species of Halisaurus from the Late Cretaceous phosphates of Morocco, and the phylogenetical relationships of the Halisaurinae (Squamata: Mosasauridae)". Zoological Journal of the Linnean Society. 143 (3): 447–472. doi:10.1111/j.1096-3642.2005.00152.x.
- Bell, G. L.; Polcyn, M. J. (2005). "Dallasaurus turneri, a new primitive mosasauroid from the Middle Turonian of Texas and comments on the phylogeny of the Mosasauridae (Squamata)". Netherlands Journal of Geoscience (Geologie en Mijnbouw). 84 (3): 177–194. CiteSeerX 10.1.1.169.9534. doi:10.1017/s0016774600020965.
- Dortangs, R. W.; Schulp, A. S.; Mulder, E. W. A.; Jagt, J. W. M.; Peeters, H. H. G.; Graaf, D. T. (2002). "A large new mosasaur from the Upper Cretaceous of the Netherlands". Netherlands Journal of Geosciences/Geologie en Mijnbouw. 81 (1): 1–8. doi:10.1017/s0016774600020515.
- Ellis, Richard (2003). Sea Dragons – Predators of the Prehistoric Oceans. University Press of Kansas. ISBN 0-7006-1269-6.
- Everhart, Michael J. (2005). Oceans Of Kansas: A Natural History Of The Western Interior Sea. Life of the Past. Bloomington: Indiana University Press. p. 322.
- Everhart, M. J. (2005). "Tylosaurus kansasensis, a new species of tylosaurine (Squamata, Mosasauridae) from the Niobrara Chalk of western Kansas, USA". Netherlands Journal of Geosciences / Geologie en Mijnbouw. 84 (3): 231–240. doi:10.1017/s0016774600021016.
- Hilton, Richard P. (2003). Dinosaurs and Other Mesozoic Reptiles of California. Berkeley: University of California Press. pp. 318. ISBN 9780520233157.
- Kass, M. S. (1999). "Prognathodon stadtmani (Mosasauridae): A new species from the Mancos Shale (Lower Campanian) of western Colorado". In Gillette, D. D. (ed.). Vertebrate Paleontology in Utah. Miscellaneous Publication. 99. Utah Geological Survey. pp. 275–294.
- Lindgren, J.; Kaddumi, H. F.; Polcyn, M. J. (2013). "Soft tissue preservation in a fossil marine lizard with a bilobed tail fin". Nature Communications. 4: 2423. Bibcode:2013NatCo...4.2423L. doi:10.1038/ncomms3423. PMID 24022259.
- Martin, J. E. (2007). "A new species of the durophagous mosasaur, Globidens (Squamata: Mosasauridae) from the Late Cretaceous Pierre Shale Group of central South Dakota, USA.". In Martin, J. E.; Parris, D. C. (eds.). The Geology and Paleontology of the Late Cretaceous Marine Deposits of the Dakotas. Special Paper. 427. Geological Society of America. pp. 177–198. doi:10.1130/2007.2427(13). ISBN 978-0-8137-2427-0.
- Mayor, Adrienne (2005). Fossil Legends of the First Americans. Princeton University Press. ISBN 0-691-11345-9.
- Nicholls, E. L.; Meckert, D. (2002). "Marine reptiles from the Nanaimo Group (Upper Cretaceous) of Vancouver Island". Canadian Journal of Earth Sciences. 39 (11): 1591–1603. Bibcode:2002CaJES..39.1591N. doi:10.1139/e02-075.
- Páramo-Fonseca, M. E. (2000). "Yaguarasaurus columbianus (Reptilia, Mosasauridae), a primitive mosasaur from the Turonian (Upper Cretaceous) of Columbia". Historical Biology. 14 (1–2): 121–131. doi:10.1080/10292380009380560.
- Polcyn, M. J.; Tchernov, E.; Jacobs, L. L. (1999). "The Cretaceous biogeography of the eastern Mediterranean with a description of a new basal mosasauroid from Ein Yabrud, Israel". Proceedings 2nd Gondwanan Dinosaur Symposium. 15: 259–290.
- Polcyn, M. J.; Tchernov, E.; Jacobs, L. L. (2003). "Haasiasaurus gen. nov., a new generic name for the basal mosasauroid Haasia Polcyn et al., 1999". Journal of Vertebrate Paleontology. 23 (2): 476. doi:10.1671/0272-4634(2003)023[0476:HGNANG]2.0.CO;2.
- Polcyn, M. J.; Bell, G. L. (2005). "A new mosasauroid, Russellosaurus coheni nov. gen. et. sp., from the early Middle Turonian of Texas". Netherlands Journal of Geosciences / Geologie en Mijnbouw. 84 (3): 321–333. doi:10.1017/s0016774600021107.
- Polcyn, M. J.; Everhart, M. J. (2008). "Description and phylogenetic analysis of a new species of Selmasaurus (Mosasauridae: Plioplatecarpinae) from the Niobrara Chalk of western Kansas". Proceedings of the Second Mosasaur Meeting, Fort Hays Studies. Hays, Kansas: Fort Hays State University (3): 13–28.
- Sakurai, K.; Chitoku, T.; Shibuya, N. (1999). "A new species of Mosasaurus (Reptilia, Mosasauridae) from Hobetsu, Hokkaido, Japan". The Bulletin of the Hobetsu Museum. 15: 53–66.
- Schulp, A. S.; Polcyn, M. J.; Mateus, O.; Jacobs, L. L.; Morais, M. L. (2008). "A new species of Prognathodon (Squamata, Mosasauridae) from the Maastrichtian of Angola, and the affinities of the mosasaur genus Liodon". Proceedings of the Second Mosasaur Meeting, Fort Hays Studies. Hays, Kansas: Fort Hays State University (3): 1–12.
- Suzuki, S. (1985). "A new species of Mosasaurus (Reptilia; Squamata) from the Upper Cretaceous Hakobuchi Group in central Hokkaido, Japan.". In Goto; et al. (eds.). Evolution and Adaptation of Marine Vertebrates. Monograph. 30. Association for Geological Collaboration in Japan.
External links

| Mosasaurinae |
| | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Halisaurinae | |||||||||||
| Russellosaurina |
| ||||||||||
| Related groups and genera | |||||||||||
| Related articles | |||||||||||