Sarcopterygii
Sarcopterygii (/sɑːrˌkɒptəˈrɪdʒiaɪ/; from Greek σάρξ sarx, flesh, and πτέρυξ pteryx, fin)—sometimes considered synonymous with Crossopterygii ("fringe-finned fish", from Greek κροσσός krossos, fringe)—is a clade (traditionally a class or subclass) of the bony fish whose members are known as lobe-finned fish. The group Tetrapoda, a superclass including amphibians, reptiles (including dinosaurs and therefore birds), and mammals, evolved from certain sarcopterygians; under a cladistic view, tetrapods are themselves considered a group within Sarcopterygii.
| Lobe-finned fishes | |
|---|---|
 | |
| Many sarcopterygians: Neoceratodus forsteri, Guiyu oneiros, Psarolepis romeri and the stegocephalian Tiktaalik roseae. | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Euteleostomi |
| Clade: | Sarcopterygii Romer, 1955 |
| Subgroups | |
| |
The living non-tetrapod sarcopterygians include two species of coelacanths and six species of lungfish.
Characteristics

Early lobe-finned fishes are bony fish with fleshy, lobed, paired fins, which are joined to the body by a single bone.[7] The fins of lobe-finned fishes differ from those of all other fish in that each is borne on a fleshy, lobelike, scaly stalk extending from the body. The scales of sarcopterygians are true scaloids, consisting of lamellar bone surrounded by layers of vascular bone, dentine-like cosmine, and external keratin.[8] The morphology of tetrapodomorphs, fish that are similar-looking to tetrapods, give indications of the transition from water to terrestrial life (Clack 2009). Pectoral and pelvic fins have articulations resembling those of tetrapod limbs. These fins evolved into the legs of the first tetrapod land vertebrates, amphibians. They also possess two dorsal fins with separate bases, as opposed to the single dorsal fin of actinopterygians (ray-finned fish). The braincase of sarcopterygians primitively has a hinge line, but this is lost in tetrapods and lungfish. Many early sarcopterygians have a symmetrical tail. All sarcopterygians possess teeth covered with true enamel.
Most species of lobe-finned fishes are extinct. The largest known lobe-finned fish was Rhizodus hibberti from the Carboniferous period of Scotland which may have exceeded 7 meters in length. Among the two groups of extant (living) species, the coelacanths and the lungfishes, the largest species is the West Indian Ocean coelacanth, reaching 2 m (6 ft 7 in) in length and weighing up 110 kg (240 lb). The largest lungfish is the African lungfish which can reach 2 m (6.6 ft) in length and weigh up to 50 kg (110 lb).[9][10]
Classification
Taxonomists who subscribe to the cladistic approach include the grouping Tetrapoda within this group, which in turn consists of all species of four-limbed vertebrates.[11] The fin-limbs of lobe-finned fishes such as the coelacanths show a strong similarity to the expected ancestral form of tetrapod limbs. The lobe-finned fishes apparently followed two different lines of development and are accordingly separated into two subclasses, the Rhipidistia (including the Dipnoi, the lungfish, and the Tetrapodomorpha which include the Tetrapoda) and the Actinistia (coelacanths).
Taxonomy
The classification below follows Benton 2004, and uses a synthesis of rank-based Linnaean taxonomy and also reflects evolutionary relationships. Benton included the Superclass Tetrapoda in the Subclass Sarcopterygii in order to reflect the direct descent of tetrapods from lobe-finned fish, despite the former being assigned a higher taxonomic rank.[12]
| Actinistia |  |
Actinistia, coelacanths, are a subclass of mostly fossil lobe-finned fishes. This subclass contains the coelacanths, including the two living coelacanths, the West Indian Ocean coelacanth and the Indonesian coelacanth. |
|---|---|---|
| Dipnoi | Dipnoi, lungfish, also known as salamanderfish,[13] are a subclass of freshwater fish. Lungfish are best known for retaining characteristics primitive within the bony fishes, including the ability to breathe air, and structures primitive within the lobe-finned fishes, including the presence of lobed fins with a well-developed internal skeleton. Today, lungfish live only in Africa, South America, and Australia. While vicariance would suggest this represents an ancient distribution limited to the Mesozoic supercontinent Gondwana, the fossil record suggests advanced lungfish had a widespread freshwater distribution and the current distribution of modern lungfish species reflects extinction of many lineages following the breakup of Pangaea, Gondwana, and Laurasia. | |
| Tetrapodomorpha | Tetrapodomorpha, tetrapods and their extinct relatives, are a clade of vertebrates consisting of tetrapods (four-limbed vertebrates) and their closest sarcopterygian relatives that are more closely related to living tetrapods than to living lungfish (Amemiya et al. 2013). Advanced forms transitional between fish and the early labyrinthodonts, like Tiktaalik, have been referred to as "fishapods" by their discoverers, being half-fish, half-tetrapods, in appearance and limb morphology. The Tetrapodomorpha contain the crown group tetrapods (the last common ancestor of living tetrapods and all of its descendants) and several groups of early stem tetrapods, and several groups of related lobe-finned fishes, collectively known as the osteolepiforms. The Tetrapodamorpha minus the crown group Tetrapoda are the stem tetrapoda, a paraphyletic unit encompassing the fish to tetrapod transition. Among the characters defining tetrapodomorphs are modifications to the fins, notably a humerus with convex head articulating with the glenoid fossa (the socket of the shoulder joint). Tetrapodomorph fossils are known from the early Devonian onwards, and include Osteolepis, Panderichthys, Kenichthys, and Tungsenia.[14] |


- Subclass Sarcopterygii
- †Order Onychodontida
- Order Actinistia
- Infraclass Dipnomorpha
- †Order Porolepiformes
- Subclass Dipnoi
- Order Ceratodontiformes
- Order Lepidosireniformes
- Infraclass Tetrapodomorpha
- †Order Rhizodontida
- Superorder Osteolepidida
- †Order Osteolepiformes
- †Family Tristichopteridae
- †Order Panderichthyida
- Superclass Tetrapoda
- †Order Osteolepiformes
Phylogeny
The cladogram presented below is based on studies compiled by Philippe Janvier and others for the Tree of Life Web Project,[15] Mikko's Phylogeny Archive [16] and Swartz 2012.[17]
| Sarcopterygii |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

- Sarcopterygii incertae sedis
- †Guiyu oneiros Zhu et al., 2009
- †Diabolepis speratus (Chang & Yu, 1984)
- †Langdenia campylognatha Janvier & Phuong, 1999
- †Ligulalepis Schultze, 1968
- †Meemannia eos Zhu, Yu, Wang, Zhao & Jia, 2006
- †Psarolepis romeri Yu 1998 sensu Zhu, Yu, Wang, Zhao & Jia, 2006
- †Megamastax ambylodus Choo, Zhu, Zhao, Jia, & Zhu, 2014
- †Sparalepis tingi Choo,Zhu,Qu,Yu,Jia & Zhaoh, 2017[18][19]
- Paraphyletic Osteolepida incertae sedis [taxa not treated by Ahlberg & Johanson, 1998]:
- †Bogdanovia orientalis Obrucheva 1955 [has been treated as Coelacanthinimorph sarcopterygian]
- †Canningius groenlandicus Säve-Söderbergh, 1937
- †Chrysolepis
- †Geiserolepis
- †Latvius
- †L. grewingki (Gross, 1933)
- †L. porosus Jarvik, 1948
- †L. obrutus Vorobyeva, 1977
- †Lohsania utahensis Vaughn, 1962
- †Megadonichthys kurikae Vorobyeva, 1962
- †Platyethmoidia antarctica Young, Long & Ritchie, 1992
- †Shirolepis ananjevi Vorobeva, 1977
- †Sterropterygion brandei Thomson, 1972
- †Thaumatolepis edelsteini Obruchev, 1941
- †Thysanolepis micans Vorobyeva, 1977
- †Vorobjevaia dolonodon Young, Long & Ritchie, 1992
- Paraphyletic Elpistostegalia/Panderichthyida incertae sedis
- †Parapanderichthys stolbovi (Vorobyeva, 1960) Vorobyeva, 1992
- †Howittichthys warrenae Long & Holland, 2008
- †Livoniana multidentata Ahlberg, Luksevic & Mark-Kurik, 2000
- Stegocephalia incertae sedis
- †Antlerpeton clarkii Thomson, Shubin & Poole, 1998
- †Austrobrachyops jenseni Colbert & Cosgriff, 1974
- †Broilisaurus raniceps (Goldenberg, 1873) Kuhn, 1938
- †Densignathus rowei Daeschler, 2000
- †Doragnathus woodi Smithson, 1980
- †Jakubsonia livnensis Lebedev, 2004
- †Limnerpeton dubium Fritsch, 1901 (nomen dubium)
- †Limnosceloides Romer, 1952
- †L. dunkardensis Romer, 1952 (Type)
- †L. brahycoles Langston, 1966
- †Occidens portlocki Clack & Ahlberg, 2004
- †Ossinodus puerorum emend Warren & Turner, 2004
- †Romeriscus periallus Baird & Carroll, 1968
- †Sigournea multidentata Bolt & Lombard, 2006
- †Sinostega pani Zhu et al., 2002
- †Ymeria denticulata Clack et al., 2012
Evolution
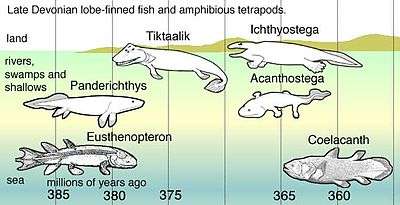
- Panderichthys, suited to muddy shallows;
- Tiktaalik with limb-like fins that could take it onto land;
- Early tetrapods in weed-filled swamps, such as:
- Acanthostega, which had feet with eight digits,
- Ichthyostega with limbs.

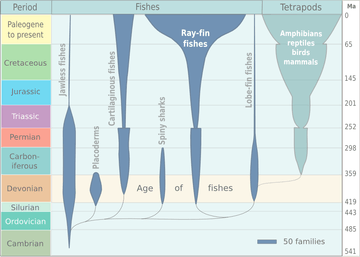
Lobe-finned fishes (sarcopterygians) and their relatives the ray-finned fishes (actinopterygians) comprise the superclass of bony fishes (Osteichthyes) characterized by their bony skeleton rather than cartilage. There are otherwise vast differences in fin, respiratory, and circulatory structures between the Sarcopterygii and the Actinopterygii, such as the presence of cosmoid layers in the scales of sarcopterygians. The earliest fossils of sarcopterygians, found in the uppermost Silurian (ca 418 Ma), closely resembled the acanthodians (the "spiny fish", a taxon that became extinct at the end of the Paleozoic). In the early–middle Devonian (416–385 Ma), while the predatory placoderms dominated the seas, some sarcopterygians came into freshwater habitats.
In the Early Devonian (416–397 Mya), the sarcopterygians split into two main lineages: the coelacanths and the rhipidistians. Coelacanths never left the oceans and their heyday was the late Devonian and Carboniferous, from 385 to 299 Ma, as they were more common during those periods than in any other period in the Phanerozoic; coelacanths (genus Latimeria) still live today in the open (pelagic) oceans.
The Rhipidistians, whose ancestors probably lived in the oceans near the river mouths (estuaries), left the ocean world and migrated into freshwater habitats. In turn, they split into two major groups: lungfish and the tetrapodomorphs. Lungfish radiated into their greatest diversity during the Triassic period; today fewer than a dozen genera remain. They evolved the first proto-lungs and proto-limbs, adapting to living outside a submerged water environment by the middle Devonian (397–385 Ma).
There are three major hypotheses as to how lungfish evolved their stubby fins (proto-limbs). The traditional explanation is the "shrinking waterhole hypothesis", or "desert hypothesis", posited by the American paleontologist Alfred Romer, who believed that limbs and lungs may have evolved from the necessity of having to find new bodies of water as old waterholes dried up.[21]
A second, the "inter-tidal hypothesis", was published by a team of Polish paleontologists—Grzegorz Niedźwiedzki, Piotr Szrek, Katarzyna Narkiewicz, Marek Narkiewicz, and Per Ahlberg—in 2010. They argued that sarcopterygians may have first emerged unto land from intertidal zones rather than inland bodies of water. Their hypothesis is based on the discovery of the 395 million-year-old Zachełmie tracks in Zachełmie, Świętokrzyskie Voivodeship, Poland, the oldest-ever-discovered fossil evidence of tetrapods.[22][23]
The third hypothesis is dubbed the "woodland hypothesis" and was proposed by the American paleontologist Greg Retallack in 2011. He argues that limbs may have developed in shallow bodies of water in woodlands as a means of navigating in environments filled with roots and vegetation. He based his conclusions on the evidence that transitional tetrapod fossils are consistently found in habitats that were formerly humid and wooded floodplains.[21][24]
A fourth, minority hypothesis posits that advancing onto land achieved more safety from predators, less competition for prey, and certain environmental advantages not found in water—such as oxygen concentration (Carroll et al. 2005, as cited by (Hohn-Schulte et al. 2013)), and temperature control (Clack 2007, as cited by (Hohn-Schulte et al. 2013))—implying that organisms developing limbs were also adapting to spending some of their time out of water. However, studies have found that sarcopterygians developed tetrapod-like limbs suitable for walking well before venturing onto land (King 2011, as cited by (Pierce et al. 2012)); this suggests they adapted to walking on the ground-bed under water before they advanced onto dry land.
The first tetrapodomorphs, which included the gigantic rhizodonts, had the same general anatomy as the lungfish, who were their closest kin, but they appear not to have left their water habitat until the late Devonian epoch (385–359 Ma), with the appearance of tetrapods (four-legged vertebrates). Tetrapods are the only tetrapodomorphs which survived after the Devonian.
Non-tetrapod sarcopterygians continued until towards the end of Paleozoic era, suffering heavy losses during the Permian–Triassic extinction event (251 Ma).
References
Citations
- Brazeau, Martin D.; Friedman, Matt (April 23, 2015). "The origin and early phylogenetic history of jawed vertebrates". Nature. 520 (7548): 490–7. Bibcode:2015Natur.520..490B. doi:10.1038/nature14438. PMC 4648279. PMID 25903631.
- "PALAEOBLOG".
- "Descubrimiento de fósil de pez óseo en China aporta nuevos conocimientos clave sobre origen de los vertebrados_Spanish.china.org.cn".
- Zhu, M; Zhao, W; Jia, L; Lu, J; Qiao, T; Qu, Q (2009). "The oldest articulated osteichthyan reveals mosaic gnathostome characters". Nature. 458 (7237): 469–474. Bibcode:2009Natur.458..469Z. doi:10.1038/nature07855. PMID 19325627.
- Coates, M.I. (2009). "Palaeontology: Beyond the Age of Fishes". Nature. 458 (7237): 413–414. Bibcode:2009Natur.458..413C. doi:10.1038/458413a. PMID 19325614.
- Pharyngula Archived 2012-03-09 at the Wayback MachineScience blogs, 1 April 2009.
- Clack, J. A. (2002) Gaining Ground. Indiana University
- Kardong, Kenneth V. (1998). Vertebrates: Comparative Anatomy, Function, Evolution, second edition, USA: McGraw-Hill, 747 pp.. ISBN 0-07-115356-X/0-697-28654-1.
- Froese, Rainer, and Daniel Pauly, eds. (2009). "Lepidosirenidae" in FishBase. January 2009 version.
- Protopterus aethiopicus Archived 2011-08-03 at the Wayback Machine. Fishing-worldrecords.com
- Nelson 2006.
- Benton, M.J. (2004). Vertebrate Paleontology. 3rd ed. Blackwell Science Ltd
- Ernst Heinrich Philipp August Haeckel, Edwin Ray Lankester, L. Dora Schmitz (1892). The History of Creation, Or, The Development of the Earth and Its Inhabitants by the Action of Natural Causes: A Popular Exposition of the Doctrine of Evolution in General, and of that of Darwin, Goethe, and Lamarck in Particular : from the 8. German Ed. of Ernst Haeckel. D. Appleton. p. 422.CS1 maint: multiple names: authors list (link) page 289
- Jing Lu, Min Zhu, John A. Long, Wenjin Zhao, Tim J. Senden, Liantao Jia and Tuo Qiao (2012). "The earliest known stem-tetrapod from the Lower Devonian of China". Nature Communications. 3: 1160. Bibcode:2012NatCo...3.1160L. doi:10.1038/ncomms2170. PMID 23093197.CS1 maint: multiple names: authors list (link)
- Janvier, Philippe. 1997. Vertebrata. Animals with backbones. Version 01 January 1997 (under construction). http://tolweb.org/Vertebrata/14829/1997.01.01 in The Tree of Life Web Project, http://tolweb.org/
- Haaramo, Mikko (2003). "Sarcopterygii". In Mikko's Phylogeny Archive. Retrieved November 4, 2013.
- Swartz, B. (2012). "A marine stem-tetrapod from the Devonian of Western North America". PLoS ONE. 7 (3): e33683. Bibcode:2012PLoSO...733683S. doi:10.1371/journal.pone.0033683. PMC 3308997. PMID 22448265.
- Choo, Brian; Zhu, Min; Qu, Qingming; Yu, Xiaobo; Jia, Liantao; Zhao, Wenjin (2017-03-08). "A new osteichthyan from the late Silurian of Yunnan, China". PLOS ONE. 12 (3): e0170929. Bibcode:2017PLoSO..1270929C. doi:10.1371/journal.pone.0170929. ISSN 1932-6203. PMC 5342173. PMID 28273081.
- PLOS. "Ancient southern China fish may have evolved prior to the 'Age of Fish". www.sciencedaily.com. Archived from the original on 2017-03-08. Retrieved 2017-03-11.
- Benton 2005.
- "Fish-Tetrapod Transition Got A New Hypothesis In 2011". Science 2.0. December 27, 2011. Retrieved January 2, 2012.
- Grzegorz Niedźwiedzki, Piotr Szrek, Katarzyna Narkiewicz, Marek Narkiewicz & Per E. Ahlberg (2010). "Tetrapod trackways from the early Middle Devonian period of Poland". Nature. Nature Publishing Group. 463 (7277): 43–48. Bibcode:2010Natur.463...43N. doi:10.1038/nature08623. PMID 20054388.CS1 maint: multiple names: authors list (link)
- Shanta Barley (January 6, 2010). "Oldest footprints of a four-legged vertebrate discovered". New Scientist. Retrieved January 3, 2010.
- Retallack, Gregory (May 2011). "Woodland Hypothesis for Devonian Tetrapod Evolution". Journal of Geology. University of Chicago Press. 119 (3): 235–258. Bibcode:2011JG....119..235R. doi:10.1086/659144.
Carroll RL, Irwin J, Green DM. 2005. Thermal physiology and the origin of terrestriality in vertebrates. Zoological Journey of the Linnean Society. 143:345–358.
Hohn-Schulte, B., H. Preuschoft, U. Witzel, and C. Distler-Hoffmann. 2013. Biomechanics and functional preconditions for terrestrial lifestyle in basal tetrapods, with special consideration of Tiktaalik roseae. Historical Biology 25:167-181.
Clack JA. 2007. Devonian climate change, breathing, and the origin of the tetrapod stem group. Integrative and Comparative Biology. p. 1–14.
King, H.M., Shubin, N.H., Coates, M.I. & Hale, M.E. Behavioural evidence for the evolution of walking and bounding before terrestriality in sarcopterygian fishes. Proceedings of the National Academy of Sciences. USA 108: 21146–21151 (2011).
Pierce, S. E., J. A. Clack, and J. R. Hutchinson. 2012. Three-dimensional limb joint mobility in the early tetrapod Ichthyostega. Nature 486:523-U123.
Clack, J. A. 2009. The fin to limb transition: New data, interpretations, and hypotheses from paleontology and developmental biology. Annual Review of Earth and Planetary Sciences 37:163-179.
Amemiya, C. T., J. Alfoldi, A. P. Lee, S. H. Fan, H. Philippe, I. MacCallum, I. Braasch et al. 2013. The African coelacanth genome provides insights into tetrapod evolution. Nature 496:311-316.
Bibliography
- Nelson, Joseph S. (2006). Fishes of the World. John Wiley & Sons. ISBN 978-0-471-25031-9.
| Forerunners |  | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jawless fish |
| ||||||||||||
| Jawed fish |
| ||||||||||||
| Lists | |||||||||||||
| Related |
| ||||||||||||
† extinct | |||||||||||||
