Insulin-like growth factor 1
Insulin-like growth factor 1 (IGF-1), also called somatomedin C, is a hormone similar in molecular structure to insulin which plays an important role in childhood growth, and has anabolic effects in adults.
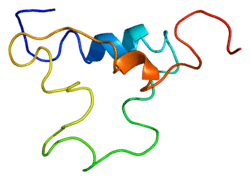
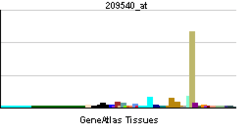
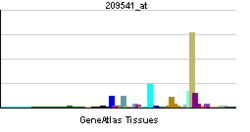
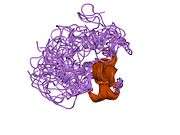
IGF-1 is a protein that in humans is encoded by the IGF1 gene.[5][6] IGF-1 consists of 70 amino acids in a single chain with three intramolecular disulfide bridges. IGF-1 has a molecular weight of 7,649 Daltons.[7]
IGF-1 is produced primarily by the liver. Production is stimulated by growth hormone (GH). Most of IGF-1 is bound to one of 6 binding proteins (IGF-BP). IGFBP-1 is regulated by insulin. IGF-1 is produced throughout life; the highest rates of IGF-1 production occur during the pubertal growth spurt. The lowest levels occur in infancy and old age.
A synthetic analog of IGF-1, mecasermin, is used in children for the treatment of growth failure.[8]
Synthesis and circulation
IGF-1 is produced primarily by the liver as an endocrine hormone as well as in target tissues in a paracrine/autocrine fashion. Production is stimulated by growth hormone (GH) and can be retarded by undernutrition, growth hormone insensitivity, lack of growth hormone receptors, or failures of the downstream signaling pathway post GH receptor including SHP2 and STAT5B. Approximately 98% of IGF-1 is always bound to one of 6 binding proteins (IGF-BP). IGFBP-3, the most abundant protein, accounts for 80% of all IGF binding. IGF-1 binds to IGFBP-3 in a 1:1 molar ratio. IGFBP-1 is regulated by insulin.[9]
IGF-1 is produced throughout life. The highest rates of IGF-1 production occur during the pubertal growth spurt. The lowest levels occur in infancy and old age.
Protein intake increases IGF-1 levels in humans, independent of total calorie consumption.[10] Factors that are known to cause variation in the levels of growth hormone (GH) and IGF-1 in the circulation include: insulin levels, genetic make-up, the time of day, age, sex, exercise status, stress levels, nutrition level and body mass index (BMI), disease state, ethnicity, estrogen status and xenobiotic intake.[11]
Mechanism of action
IGF-1 is a primary mediator of the effects of growth hormone (GH). Growth hormone is made in the anterior pituitary gland, is released into the blood stream, and then stimulates the liver to produce IGF-1. IGF-1 then stimulates systemic body growth, and has growth-promoting effects on almost every cell in the body, especially skeletal muscle, cartilage, bone, liver, kidney, nerve, skin, hematopoietic, and lung cells. In addition to the insulin-like effects, IGF-1 can also regulate cellular DNA synthesis.[12]
IGF-1 binds to at least two cell surface receptor tyrosine kinases: the IGF-1 receptor (IGF1R), and the insulin receptor. Its primary action is mediated by binding to its specific receptor, IGF1R, which is present on the surface of many cell types in many tissues. Binding to the IGF1R initiates intracellular signaling. IGF-1 is one of the most potent natural activators of the AKT signaling pathway, a stimulator of cell growth and proliferation, and a potent inhibitor of programmed cell death .[13][14] The IGF-1 receptor seems to be the "physiologic" receptor because it binds IGF-1 with significantly higher affinity than insulin receptor does. IGF-1 activates the insulin receptor at approximately 0.1 times the potency of insulin. Part of this signaling may be via IGF1R/Insulin Receptor heterodimers (the reason for the confusion is that binding studies show that IGF1 binds the insulin receptor 100-fold less well than insulin, yet that does not correlate with the actual potency of IGF1 in vivo at inducing phosphorylation of the insulin receptor, and hypoglycemia).
IGF-1 binds and activates its own receptor, IGF-1R, through the cell surface expression of Receptor Tyrosine Kinase's (RTK's)[15] and further signal through multiple intracellular transduction cascades. IGF-1R is the critical role-playing inducer in modulating the metabolic effects of IGF-1 for cellular senescence and survival. At a localized target cell, IGF-1R elicits the mediation of paracrine activity. After its activation the initiation of intracellular signaling occurs inducing a magnitude of signaling pathways. An important mechanistic pathway involved in mediating a cascade affect a key pathway regulated by phosphatidylinositol-3 kinase (PI3K) and its downstream partner, mTOR (mammalian Target of Rapamycin)[15]. Rapamycin binds with the enzyme FKBPP12 to inhibit the mTORC1 complex. mTORC2 remains unaffected and responds by up-regulating AKT, driving signals through the inhibited mTORC1. Phosphorylation of Eukaryotic translation initiation factor 4E (EIF4E) by mTOR suppresses the capacity of Eukaryotic translation initiation factor 4E-binding protein 1 (EIF4EBP1) to inhibit EIF4E and slow metabolism.[16] A mutation in the signaling pathway PI3K-AKT-mTOR is a big factor in the formation of tumors found predominantly on skin, internal organs, and secondary lymph nodes (Kaposi sarcoma).[17] IGF-1R allows the activation of these signaling pathways and subsequently regulates the cellular longevity and metabolic re-uptake of biogenic substances. A therapeutic approach targeting towards the reduction of such tumor collections could be induced by ganitumab. Ganitumab is a monoclonal antibody (mAb) directed antagonistically against IGF-1R. Ganitumab binds to IGF-1R, preventing binding of IGF-1 and the subsequent triggering of the PI3K-mTOR signaling pathway; inhibition of this pro-survival pathway may result in the inhibition of tumor cell expansion and the induction of tumor cell apoptosis.
Insulin-like growth factor 1 has been shown to bind and interact with all seven IGF-1 binding proteins (IGFBPs): IGFBP1, IGFBP2, IGFBP3, IGFBP4, IGFBP5, IGFBP6, and IGFBP7. Some IGFBPs are inhibitory. For example, both IGFBP-2 and IGFBP-5 bind IGF-1 at a higher affinity than it binds its receptor. Therefore, increases in serum levels of these two IGFBPs result in a decrease in IGF-1 activity.
Related growth factors
IGF-1 is closely related to a second protein called "IGF-2". IGF-2 also binds the IGF-1 receptor. However, IGF-2 alone binds a receptor called the "IGF-2 receptor" (also called the mannose-6 phosphate receptor). The insulin-like growth factor-II receptor (IGF2R) lacks signal transduction capacity, and its main role is to act as a sink for IGF-2 and make less IGF-2 available for binding with IGF-1R. As the name "insulin-like growth factor 1" implies, IGF-1 is structurally related to insulin, and is even capable of binding the insulin receptor, albeit at lower affinity than insulin.
A splice variant of IGF-1 sharing an identical mature region, but with a different E domain is known as mechano-growth factor (MGF).[18]
Disorders
Laron dwarfism
Rare diseases characterized by inability to make or respond to IGF-1 produce a distinctive type of growth failure. One such disorder, termed Laron dwarfism does not respond at all to growth hormone treatment due to a lack of GH receptors. The FDA has grouped these diseases into a disorder called severe primary IGF deficiency. Patients with severe primary IGFD typically present with normal to high GH levels, height below 3 standard deviations (SD), and IGF-1 levels below 3 SD. Severe primary IGFD includes patients with mutations in the GH receptor, post-receptor mutations or IGF mutations, as previously described. As a result, these patients cannot be expected to respond to GH treatment.
People with Laron syndrome have very low rates of cancer and diabetes.[19] Notably people with untreated Laron syndrome also never develop acne.[20]
Acromegaly
Acromegaly is a syndrome that results when the anterior pituitary gland produces excess growth hormone (GH). A number of disorders may increase the pituitary's GH output, although most commonly it involves a tumor called pituitary adenoma, derived from a distinct type of cell (somatotrophs). It leads to anatomical changes and metabolic dysfunction caused by both an elevated GH and elevated IGF-1 levels.[21] High level of IGF-1 in acromegaly is related to an increased risk of some cancers, particularly colon cancer and thyroid cancer.[22]
Cancer
A mutation in the signaling pathway PI3K-AKT-mTOR is a factor in the formation of tumors found predominantly on skin, internal organs, and secondary lymph nodes (Kaposi sarcoma).[23]
IGF-1R allows the activation of these signaling pathways and subsequently regulates the cellular longevity and metabolic re-uptake of biogenic substances. A therapeutic approach targeting towards the reduction of such tumor collections could be induced by ganitumab. Ganitumab is a monoclonal antibody (mAb) directed antagonistically against IGF-1R. Ganitumab binds to IGF-1R, preventing binding of IGF-1 and the subsequent triggering of the PI3K-mTOR signaling pathway; inhibition of this pro-survival pathway may result in the inhibition of tumor cell expansion and the induction of tumor cell apoptosis.
Use as a diagnostic test
IGF-1 levels can be measured in the blood in 10-1000 ng/ml amounts. As levels do not fluctuate greatly throughout the day for an individual person, IGF-1 is used by physicians as a screening test for growth hormone deficiency and excess in acromegaly and gigantism.
Interpretation of IGF-1 levels is complicated by the wide normal ranges, and marked variations by age, sex, and pubertal stage. Clinically significant conditions and changes may be masked by the wide normal ranges. Sequential measurement over time is often useful for the management of several types of pituitary disease, undernutrition, and growth problems.
Possible causes of elevated IGF-1 levels
- acromegaly (especially when GH is also high)
- high-protein diet [24]
- high glycemic-index diet[25]
- dairy products (except for cheese) consumption[25][26]
- delayed puberty[27]
- pregnancy[28]
- hyperthyroidism[28]
- IGF-1 assay problems[28]
- some rare tumors (i.e. carcinoids) secreting IGF-1[29]
Use as a therapeutic agent
Patients with severe primary insulin-like growth factor-1 deficiency (IGFD), called Laron syndrome, may be treated with either IGF-1 alone or in combination with IGFBP-3.[30] Mecasermin (brand name Increlex) is a synthetic analog of IGF-1 which is approved for the treatment of growth failure.[30] IGF-1 has been manufactured recombinantly on a large scale using both yeast and E. coli.
Clinical trials
Recombinant protein
Several companies have evaluated administering recombinant IGF-1 in clinical trials for type 1 diabetes, type 2 diabetes, amyotrophic lateral sclerosis,[31] severe burn injury and myotonic muscular dystrophy.
Results of clinical trials evaluating the efficacy of IGF-1 in type 1 diabetes and type 2 diabetes showed reduction in hemoglobin A1C levels and daily insulin consumption. However the sponsor discontinued the program due to an exacerbation of diabetic retinopathy,[32] coupled with a shift in corporate focus towards oncology.
Two clinical studies of IGF-1 for ALS were conducted and although one study demonstrated efficacy the second was equivocal, and the product was not submitted for approval to the FDA.
Society and culture
History of name
In the 1950s IGF-1 was called "sulfation factor" because it stimulated sulfation of cartilage in vitro,[33] and in the 1970s due to its effects it was termed "nonsuppressible insulin-like activity" (NSILA).
See also
References
- GRCh38: Ensembl release 89: ENSG00000017427 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000020053 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
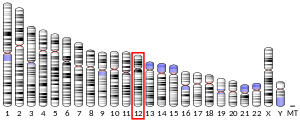
- Höppener JW, de Pagter-Holthuizen P, Geurts van Kessel AH, Jansen M, Kittur SD, Antonarakis SE, Lips CJ, Sussenbach JS (1985). "The human gene encoding insulin-like growth factor I is located on chromosome 12". Hum. Genet. 69 (2): 157–60. doi:10.1007/BF00293288. PMID 2982726.
- Jansen M, van Schaik FM, Ricker AT, Bullock B, Woods DE, Gabbay KH, Nussbaum AL, Sussenbach JS, Van den Brande JL (1983). "Sequence of cDNA encoding human insulin-like growth factor I precursor". Nature. 306 (5943): 609–11. doi:10.1038/306609a0. PMID 6358902.
- Rinderknecht E, Humbel RE (1978). "The amino acid sequence of human insulin-like growth factor I and its structural homology with proinsulin". J Biol Chem. 253 (8): 2769–2776. PMID 632300.
- Keating GM (2008). "Mecasermin". BioDrugs. 22 (3): 177–88. doi:10.2165/00063030-200822030-00004. PMID 18481900.
- Christoffersen CT, Bornfeldt KE, Rotella CM, Gonzales N, Vissing H, Shymko RM, et al. (July 1994). "Negative cooperativity in the insulin-like growth factor-I receptor and a chimeric IGF-I/insulin receptor". Endocrinology. 135 (1): 472–5. doi:10.1210/endo.135.1.8013387. PMID 8013387.
- Levine ME, Suarez JA, Brandhorst S, Balasubramanian P, Cheng CW, Madia F, et al. (March 2014). "Low protein intake is associated with a major reduction in IGF-1, cancer, and overall mortality in the 65 and younger but not older population". Cell Metabolism. 19 (3): 407–17. doi:10.1016/j.cmet.2014.02.006. PMC 3988204. PMID 24606898.
- Scarth JP (2006). "Modulation of the growth hormone-insulin-like growth factor (GH-IGF) axis by pharmaceutical, nutraceutical and environmental xenobiotics: an emerging role for xenobiotic-metabolizing enzymes and the transcription factors regulating their expression. A review". Xenobiotica. 36 (2–3): 119–218. doi:10.1080/00498250600621627. PMID 16702112.
- Yakar S, Rosen CJ, Beamer WG, Ackert-Bicknell CL, Wu Y, Liu JL, et al. (September 2002). "Circulating levels of IGF-1 directly regulate bone growth and density". The Journal of Clinical Investigation. 110 (6): 771–81. doi:10.1172/JCI15463. PMC 151128. PMID 12235108.
- Peruzzi F, Prisco M, Dews M, Salomoni P, Grassilli E, Romano G, et al. (October 1999). "Multiple signaling pathways of the insulin-like growth factor 1 receptor in protection from apoptosis". Molecular and Cellular Biology. 19 (10): 7203–15. doi:10.1128/mcb.19.10.7203. PMC 84713. PMID 10490655.
- Juin P, Hueber AO, Littlewood T, Evan G (June 1999). "c-Myc-induced sensitization to apoptosis is mediated through cytochrome c release". Genes & Development. 13 (11): 1367–81. doi:10.1101/gad.13.11.1367. PMC 316765. PMID 10364155.
- Laron Z (October 2001). "Insulin-like growth factor 1 (IGF-1): a growth hormone". Molecular Pathology. 54 (5): 311–6. doi:10.1136/mp.54.5.311. PMC 1187088. PMID 11577173.
- Martin D, Nguyen Q, Molinolo A, Gutkind JS (May 2014). "Accumulation of dephosphorylated 4EBP after mTOR inhibition with rapamycin is sufficient to disrupt paracrine transformation by the KSHV vGPCR oncogene". Oncogene. 33 (18): 2405–12. doi:10.1038/onc.2013.193. PMID 23708663.
- Wang Z, Feng X, Molinolo AA, Martin D, Vitale-Cross L, Nohata N, et al. (April 2019). "4E-BP1 Is a Tumor Suppressor Protein Reactivated by mTOR Inhibition in Head and Neck Cancer". Cancer Research. 79 (7): 1438–1450. doi:10.1158/0008-5472.CAN-18-1220. PMC 6445709. PMID 30894372.
- Carpenter V, Matthews K, Devlin G, Stuart S, Jensen J, Conaglen J, Jeanplong F, Goldspink P, Yang SY, Goldspink G, Bass J, McMahon C (February 2008). "Mechano-growth factor reduces loss of cardiac function in acute myocardial infarction". Heart Lung Circ. 17 (1): 33–9. doi:10.1016/j.hlc.2007.04.013. PMID 17581790.
- Wade N (17 February 2011). "Ecuadorean Villagers May Hold Secret to Longevity". New York Times.
- Khanna N, Kubba R (28 February 2014). World Clinics: Dermatology - Acne. JP Medical Ltd. ISBN 9789350909768.
- Giustina A, Chanson P, Kleinberg D, Bronstein MD, Clemmons DR, Klibanski A, van der Lely AJ, Strasburger CJ, Lamberts SW, Ho KK, Casanueva FF, Melmed S (2014). "Expert consensus document: A consensus on the medical treatment of acromegaly". Nat Rev Endocrinol. 10 (4): 243–8. doi:10.1038/nrendo.2014.21. PMID 24566817.
- AlDallal S (August 2018). "Acromegaly: a challenging condition to diagnose". review. International Journal of General Medicine. 11: 337–343. doi:10.2147/IJGM.S169611. PMID 30197531.
- Wang Z, Feng X, Molinolo AA, Martin D, Vitale-Cross L, Nohata N, et al. (April 2019). "4E-BP1 Is a Tumor Suppressor Protein Reactivated by mTOR Inhibition in Head and Neck Cancer". Cancer Research. 79 (7): 1438–1450. doi:10.1158/0008-5472.CAN-18-1220. PMC 6445709. PMID 30894372.
- Levine ME, Suarez JA, Brandhorst S, Balasubramanian P, Cheng CW, Madia F, et al. (March 2014). "Low protein intake is associated with a major reduction in IGF-1, cancer, and overall mortality in the 65 and younger but not older population". primary. Cell Metabolism. 19 (3): 407–17. doi:10.1016/j.cmet.2014.02.006. PMID 24606898.
- Sakuma TH, Maibach HI (2012). "Oily skin: an overview". review. Skin Pharmacology and Physiology. 25 (5): 227–35. doi:10.1159/000338978. PMID 22722766.
- Melnik BC, John SM, Schmitz G (June 2011). "Over-stimulation of insulin/IGF-1 signaling by western diet may promote diseases of civilization: lessons learnt from laron syndrome". primary. Nutrition & Metabolism. 8: 41. doi:10.1186/1743-7075-8-41. PMC 3141390. PMID 21699736.
- Imran SA, Pelkey M, Clarke DB, Clayton D, Trainer P, Ezzat S (2010). "Spuriously Elevated Serum IGF-1 in Adult Individuals with Delayed Puberty: A Diagnostic Pitfall". primary. International Journal of Endocrinology. 2010. doi:10.1155/2010/370692. PMC 2939391. PMID 20862389.
- Freda PU (August 2009). "Monitoring of acromegaly: what should be performed when GH and IGF-1 levels are discrepant?". review. Clinical Endocrinology. 71 (2): 166–70. doi:10.1111/j.1365-2265.2009.03556.x. PMC 3654652. PMID 19226264.
- Phillips JD, Yeldandi A, Blum M, de Hoyos A (October 2009). "Bronchial carcinoid secreting insulin-like growth factor-1 with acromegalic features". primary. The Annals of Thoracic Surgery. 88 (4): 1350–2. doi:10.1016/j.athoracsur.2009.02.042. PMID 19766843.
- Rosenbloom AL (2007). "The role of recombinant insulin-like growth factor I in the treatment of the short child". Curr. Opin. Pediatr. 19 (4): 458–64. doi:10.1097/MOP.0b013e3282094126. PMID 17630612.
- Vaught JL, Contreras PC, Glicksman MA, Neff NT (1996). "Potential utility of rhIGF-1 in neuromuscular and/or degenerative disease". Ciba Found. Symp. 196: 18–27, discussion 27–38. PMID 8866126.
- "Genentech Discontinues IGF-I Drug Development Effort in Diabetes" (Press release). Genentech. 5 September 1997. Retrieved 15 March 2013.
- Salmon WD, Daughaday WH (1957). "A hormonally controlled serum factor which stimulates sulfate incorporation by cartilage in vitro". J Lab Clin Med. 49 (6): 825–36. PMID 13429201.
External links
- Insulin-Like+Growth+Factor+I at the US National Library of Medicine Medical Subject Headings (MeSH)
- Overview of all the structural information available in the PDB for UniProt: P05019 (Insulin-like growth factor I) at the PDBe-KB.
PDB gallery | |
|---|---|
|