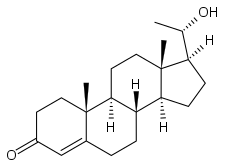
20α-Dihydroprogesterone
20α-Dihydroprogesterone (20α-DHP), also known as 20α-hydroxyprogesterone (20α-OHP), is a naturally occurring, endogenous progestogen.[1][2][3] It is a metabolite of progesterone, formed by the 20α-hydroxysteroid dehydrogenases (20α-HSDs) AKR1C1, AKR1C2, and AKR1C3 and the 17β-hydroxysteroid dehydrogenase (17β-HSD) HSD17B1.[4][5] 20α-DHP can be transformed back into progesterone by 20α-HSDs and by the 17β-HSD HSD17B2.[6] HSD17B2 is expressed in the human endometrium.[7][8][9] In animal studies, 20α-DHP has been found to be selectively taken up into and retained in target tissues such as the uterus, brain, and skeletal muscle.[6]
 | |
 | |
| Names | |
|---|---|
| IUPAC name
(8S,9S,10R,13S,14S,17S)-17-[(1S)-1-hydroxyethyl]-10,13-dimethyl-1,2,6,7,8,9,11,12,14,15,16,17-dodecahydrocyclopenta[a]phenanthren-3-one | |
| Other names
20α-DHP; 20α-Hydroxyprogesterone; 20α-OHP; 20α-Hydroxypregn-4-en-3-one; Pregn-4-en-20α-ol-3-one | |
| Identifiers | |
| |
3D model (JSmol) |
|
| ChemSpider | |
| ECHA InfoCard | 100.005.137 |
| EC Number |
|
| MeSH | 20-alpha-Dihydroprogesterone |
PubChem CID |
|
| UNII | |
| |
| |
| Properties | |
| C21H32O2 | |
| Molar mass | 316.478 g/mol |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa). | |
| Infobox references | |
20α-DHP has very low affinity for the progesterone receptor and is much less potent as a progestogen in comparison to progesterone, with about one-fifth of the relative progestogenic activity.[1][2][3][10][11][6][12] It has also been found to act as an aromatase inhibitor and to inhibit the production of estrogen in breast tissue in vitro.[13]
A single 200-mg oral dose of micronized progesterone has been found to result in peak levels of 20α-DHP of around 1 ng/mL after 2 hours.[14] In another study however, peak levels of 20α-DHP were around 10 ng/mL during therapy with 300 mg/day oral micronized progesterone.[15] 20α-DHP is formed from progesterone in the liver and in target tissues such as the endometrium.[15] It appears to be more slowly eliminated than progesterone.[15]
Levels of 5α-DHP have been quantified.[16]
See also
References
- Beranič N, Gobec S, Rižner TL (2011). "Progestins as inhibitors of the human 20-ketosteroid reductases, AKR1C1 and AKR1C3". Chem. Biol. Interact. 191 (1–3): 227–33. doi:10.1016/j.cbi.2010.12.012. PMID 21182831.
- Tony M. Plant; Anthony J. Zeleznik (15 November 2014). Knobil and Neill's Physiology of Reproduction: Two-Volume Set. Academic Press. pp. 1–. ISBN 978-0-12-397769-4.
- Cynthia L. Darlington (27 April 2009). The Female Brain. CRC Press. pp. 4–. ISBN 978-1-4200-7745-2.
- Marianne J. Legato (29 October 2009). Principles of Gender-Specific Medicine. Academic Press. pp. 617–. ISBN 978-0-08-092150-1.
- Rižner TL, Penning TM (January 2014). "Role of aldo-keto reductase family 1 (AKR1) enzymes in human steroid metabolism". Steroids. 79: 49–63. doi:10.1016/j.steroids.2013.10.012. PMC 3870468. PMID 24189185.
- Nowak FV (April 2002). "Distribution and metabolism of 20 alpha-hydroxylated progestins in the female rat". J. Steroid Biochem. Mol. Biol. 80 (4–5): 469–79. doi:10.1016/S0960-0760(02)00039-0. PMID 11983494.
- Casey ML, MacDonald PC, Andersson S (November 1994). "17 beta-Hydroxysteroid dehydrogenase type 2: chromosomal assignment and progestin regulation of gene expression in human endometrium". J. Clin. Invest. 94 (5): 2135–41. doi:10.1172/JCI117569. PMC 294662. PMID 7962560.
- Lora Hedrick Ellenson (1 December 2016). Molecular Genetics of Endometrial Carcinoma. Springer. pp. 12–. ISBN 978-3-319-43139-0.
- Byrns MC (January 2014). "Regulation of progesterone signaling during pregnancy: implications for the use of progestins for the prevention of preterm birth". J. Steroid Biochem. Mol. Biol. 139: 173–81. doi:10.1016/j.jsbmb.2013.01.015. PMID 23410596.
- Bertram G. Katzung (30 November 2017). Basic and Clinical Pharmacology 14th Edition. McGraw-Hill Education. p. 728. ISBN 978-1-259-64116-9.
In addition to progesterone, 20α- and 20β-hydroxyprogesterone (20α- and 20β-hydroxy-4-pregnene-3-one) also are found. These compounds have about one-fifth the progestational activity of progesterone in humans and other species.
- Ogle TF, Beyer BK (February 1982). "Steroid-binding specificity of the progesterone receptor from rat placenta". J. Steroid Biochem. 16 (2): 147–50. doi:10.1016/0022-4731(82)90160-1. PMID 7078152.
- Zander J, Forbes TR, Von Munstermann AM, Neher R (April 1958). "Delta 4-3-Ketopregnene-20 alpha-ol and delta 4-3-ketopregnene-20 beta-ol, two naturally occurring metabolites of progesterone; isolation, identification, biologic activity and concentration in human tissues". J. Clin. Endocrinol. Metab. 18 (4): 337–53. doi:10.1210/jcem-18-4-337. PMID 13513735.
- Pasqualini JR, Chetrite G (2008). "The anti-aromatase effect of progesterone and of its natural metabolites 20alpha- and 5alpha-dihydroprogesterone in the MCF-7aro breast cancer cell line". Anticancer Res. 28 (4B): 2129–33. PMID 18751385.
- Sitruk-Ware R, Bricaire C, De Lignieres B, Yaneva H, Mauvais-Jarvis P (October 1987). "Oral micronized progesterone. Bioavailability pharmacokinetics, pharmacological and therapeutic implications--a review". Contraception. 36 (4): 373–402. doi:10.1016/0010-7824(87)90088-6. PMID 3327648.
- Padwick ML, Endacott J, Matson C, Whitehead MI (September 1986). "Absorption and metabolism of oral progesterone when administered twice daily". Fertil. Steril. 46 (3): 402–7. doi:10.1016/S0015-0282(16)49576-2. PMID 3743792.
- Trabert B, Falk RT, Stanczyk FZ, McGlynn KA, Brinton LA, Xu X (September 2015). "Reproducibility of an assay to measure serum progesterone metabolites that may be related to breast cancer risk using liquid chromatography-tandem mass spectrometry". Horm Mol Biol Clin Investig. 23 (3): 79–84. doi:10.1515/hmbci-2015-0026. PMC 4966666. PMID 26353176.