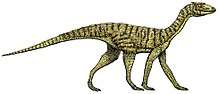
Phytosaur
Phytosaurs are an extinct group of large, mostly semiaquatic Late Triassic archosauriform reptiles. Phytosaurs belong to the family Phytosauridae and the order Phytosauria. Phytosauria and Phytosauridae are often considered to be equivalent groupings containing the same species, but some studies have identified non-phytosaurid phytosaurians. Phytosaurs were long-snouted and heavily armoured, bearing a remarkable resemblance to modern crocodilians in size, appearance, and lifestyle, as an example of convergence or parallel evolution. The name "phytosaur" means "plant reptile", as the first fossils of phytosaurs were mistakenly thought to belong to plant eaters. The name is misleading because the sharp teeth in phytosaur jaws clearly show that they were predators.
| Phytosaurs | |
|---|---|
 | |
| Skull of Machaeroprosopus mccauleyi | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Archosauriformes |
| Clade: | Crurotarsi von Meyer, 1861 |
| Order: | †Phytosauria Jaeger, 1828 |
| Genera | |
| |
| Synonyms | |
|
Parasuchia Huxley, 1875 | |
For many years, phytosaurs were considered to be the most basal group of Pseudosuchia (crocodile-line archosaurs), meaning that they were thought to be more closely related to the crocodilians than to birds (the other living group of archosaurs). Some recent studies of the evolutionary relationships of early archosauriforms suggest that phytosaurs evolved before the split between crocodile- and bird-line archosaurs and are the sister taxon of Archosauria. Others retain the older classification of phytosaurs as pseudosuchians.
Phytosaurs had a nearly global distribution during the Triassic. Fossils have been recovered from Europe, North America, India, Morocco, Thailand, Brazil, Greenland[1] and Madagascar. Fossils attributed to phytosaurs have been found in Early Jurassic rocks, possibly extending their temporal range beyond the Triassic-Jurassic boundary.
Description
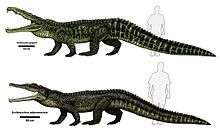
Phytosaurs are known from many different morphologies, specifically with vastly different skull forms. These changes relate to the feeding and habits of the animals, not completely evolutionary modifications. Dolichorostral ("long snouted") phytosaurs have a long, slender snout with many conical teeth that are homodont (all the same). These taxa were most likely piscivores that were well adapted to capture fast aquatic prey, but not terrestrial animals. Paleorhinus, Rutiodon and Mystriosuchus are dolichorostral phytosaurs, but do not form a distinct group of taxa (named Mystriosuchinae of Friedrich von Huene) as other morphotypes such as Pseudopalatus are more closely related to Mystriosuchus than it is to the other long-snouted taxa. Brachyrostral ("short snouted") forms are the opposite, having a massive, broad snout, and very strong skulls and jaws. They are heterodont, as the front teeth are prominent fangs, and the rear teeth are blade-like for slicing food into chunks that can easily be swallowed. Taxa like this, such as Nicrosaurus and Smilosuchus, were powerful taxa that fed on stronger prey, such as terrestrial animals that came to the water to drink. Altirostral ("high snouted") animals are intermediate between the two distinct types. They had heterodont dentition but not as extremely developed as the brachyrostral type. Pseudopalatus is an altirostral phytosaur, and was most likely a generalist feeder. Modern crocodilians exhibit a similar morphological diversity, for example the broad snouted altirostral alligator and the long snouted dolichorostral gavial.[2]
Various phytosaurs have crests and similar ornamentions in their snouts. Nicrosaurus has a ridge along the snout that would have supported a keratinous crest in life, while Mystriosuchus westphali has several bony crests.[2]
Differences from crocodiles
Despite their great similarities in appearance and lifestyle, there are still a number of minor differences that distinguish phytosaurs from true crocodiles. For one thing, the phytosaur ankle structure is much more primitive than that of any crocodile. Also, phytosaurs lack the bony secondary palate that enables crocodiles to breathe even when the mouth is full of water. However, it is possible that phytosaurs had a fleshy palate, as many Mesozoic crocodiles are presumed to have had. Phytosaurs were even better armoured than crocodiles, protected by heavy bony scutes (often found as fossils), and the belly reinforced with a dense arrangement of gastralia (abdominal ribs). Finally, and most noticeably, phytosaurs had nostrils placed near or above the level of the eyes, in contrast to crocodiles where the nostrils are near the end of the snout. This adaptation may have developed to allow them to breathe while the rest of the body was submerged.
Teeth
Unlike most crocodilians, phytosaurs have tooth serrations.[3]
In a 2001 study of the biomechanics of the dinosaur Albertosaurus's teeth, William L. Abler also examined a phytosaur's teeth, finding that it had had serrations so fine that they resembled a crack in the tooth.[4] Albertosaurus had similarly crack-like serrations, but, at the base of each serration Abler discovered a round void, which would have functioned to distribute force over a larger surface area.[4] This void, termed an ampulla, would hinder the ability of the "crack" formed by the serration to propagate through the tooth.[4] The phytosaur was found to lack adaptations for preventing its dental "cracks" from propagating.[4] Abler examined another sort of prehistoric predator, Dimetrodon, and found that it also lacked adaptations for guarding against crack propagation.[4]
Locomotion and terrestriality
Phytosaurs have been traditionally held as rather "primitive" animals in regards to terrestrial locomotion, particularly in regards to archosaurs such as crocodilians, lacking the erect gait seen in these, other pseudosuchians, dinosaurs and pterosaurs. However, the Apatopus ichnofossil shows that the animals did in fact have an erect gait like their archosaur relatives.[5][6]
Most phytosaurs are thought to be aquatic animals, and indeed most do show adaptations for such a lifestyle; swim tracks attributed to phytosaurs, for example, are known.[3] However, at least Nicrosaurus seems to have evolved towards a secondarily terrestrial lifestyle, developing longer limb bones, straighter femora and a deeper pelvis, and indeed occurs in terrestrial or marginal lacustrine settings. Combined with its deep upper jaw, it probably led a similar lifestyle to terrestrial predatory crocodylomorphs like sebecians.[7]
Inversely, some dolichorostral forms like Mystriosuchus have become further specialised to life in the water, and occurred in marine environments.[8] A skeleton of Mystriosuchus planirostris, found in a marine setting and with evidence of little post-mortem transportation - indicating that it died either at sea or in a freshwater environment nearby - shows that this animal had paddle-like limbs, less adapted for terrestrial locomotion than in most other phytosaurs.[9] Furthermore, the tail of Mystriosuchus was laterally compressed and could have been used in propulsion.[10]
Endocast Studies
Scans on various phytosaur braincases suggest that these animals generally had long olfactory tracts, weakly demarcated cerebral regions, dorsoventrally short endosseous labyrinths and various sinuses, including large antorbital and dural venous ones; the general bauplan is vaguely similar to that of crocodilians, but differs significantly in the presence of multiple sinuses, smaller cerebral hemispheres and smaller endosseous labyrinths. The similarities are considered to be plesiomorphic in relation to the ancestral archosauriform design, lacking many features seen in avemetatarsalians, though convergence in terms of lifestyle might also play a role.[11]
Reproduction
No phytosaur eggs have been found so far. There are pits associated with footprints in the Chinle Formation, but these "nests" are apparently the result of sandstone weathering.[12] A recent study suggests they might have had parental care.[13]
History

When the first phytosaur fossils were found, it was not immediately obvious what kind of animal/species they were. The first phytosaur species known to science was named Phytosaurus cylindricodon - "plant lizard with cylindrical teeth" - by G. Jaeger in 1828 because he mistakenly believed that petrified mud fillings in the jaw were herbivore teeth. The specimen is too poor to be diagnostic, and this species name is no longer valid. The name of the group - Phytosauria - was coined by the German paleontologist Hermann von Meyer in 1861, on the basis of this first species.
The next species to be described was Belodon plieningeri by von Meyer in von Meyer and Plieninger 1844. The altogether more appropriate name Parasuchia ("alongside the crocodiles", as they resembled crocodiles to a great degree) was coined by Thomas Huxley in 1875 along with his discovery and naming of the Indian species Parasuchus hislopi (Chatterjee, 1978), on the basis of a partial snout. The specimen also is usually considered non-diagnostic, and the name Parasuchus replaced by Paleorhinus. Although the names Parasuchidae and Phytosauridae are variously still used by different specialists, "phytosaur" is the standard generic name for these animals, despite the fact that these animals have been clearly shown to be carnivorous.
Evolutionary history

Phytosaurs first appeared during the Carnian age, evolving from an unknown crurotarsan ancestor. There are no clear intermediate forms, as even the earliest known phytosaurs are highly specialized aquatic animals, unlike most contemporary archosauriforms that were terrestrial. However, a recent study has suggested that Diandongosuchus is a basal phytosaur. If this is the case, this taxon offers more of a bridge between phytosaurs and earlier Archosauriformes.[14]
The earliest phytosaurs are traditionally classified in the genus Paleorhinus, now thought to be polyphyletic. Parasuchus and related basal species were widely distributed, meaning that phytosaurs dispersed across Pangea early on and there were probably few geographical barriers for their distribution; only in the southernmost regions are they rare, possibly due to increased aridity.[15]
A somewhat more advanced and larger form, Angistorhinus appears at the same time or soon after. Later in the Carnian, both these animals were replaced by more specialised forms like Rutiodon, Leptosuchus, and the huge Smilosuchus (Lucas 1998). The Carnian-Norian extinction meant that these animals died off, and the Early Norian sees new genera like Nicrosaurus and Pseudopalatus, both of which belong to the most derived clade of phytosaurs, the Pseudopalatinae. Later in the middle Norian the advanced and specialised fish-eater Mystriosuchus appears. Fossil remains of this widespread animal is known from Germany, northern Italy, and Thailand. Finally the large Redondasaurus in south-west North America and the long-snouted (altirostral) Angistorhinopsis ruetimeyeri in Europe continued the group into the Rhaetian. Phytosaur footprints (the ichnotaxon Apatopus) are also known from the latest Rhaetian of the East Coast of USA (the Newark Supergroup) (Olsen et al. 2002). This indicates that phytosaurs continued as successful animals until the very end of the Triassic, when, along with many other large crurotarsan reptiles, they were killed off by the end Triassic extinction event, about 200 Ma ago.
There have been reports of phytosaur remains found in lowermost Jurassic rocks. Several teeth from Early Jurassic deposits in France have been identified as phytosaur teeth, but other studies argue they have either been misidentified or were reworked from Late Triassic into Early Jurassic deposits. In 1951, a partial upper jaw was discovered in the Early Jurassic Lower Lufeng Series in China and described as a new genus of phytosaur, Pachysuchus, but a study in 2012 reinterpreted the fossil as a sauropodomorph dinosaur.[16]
A fragment of a lower jaw from a longirostrine archosaur has been described from early Hettangian strata in the town of Watchet in Somerset, England. While teleosaurid thalattosuchians had similar longirostrine jaws to phytosaurs and were common in the Jurassic, they do not appear in the earliest Jurassic rocks. The mandible is more similar to those of known phytosaurs than to thalattosuchians, and likely belongs to a phytosaur closely related to the genus Mystriosuchus. The presence of phytosaurs in the earliest Jurassic may have prevented thalattosuchians from occupying similar ecological niches at that time.[17] However, more recent work suggests that the jaw fragment came from a pre-Hettangian rock unit, and is therefore Late Triassic in age.[16]
Classification
Genera
| Genus | Status | Age | Location | Unit | Notes | Images |
|---|---|---|---|---|---|---|
| Junior synonym | Junior synonym of Nicrosaurus | |||||
| Valid | Late Carnian |
Argana Formation |
 | |||
| Valid | Middle Carnian |
Argana Formation |
||||
| Junior synonym | Junior synonym of Machaeroprosopus | |||||
| Nomen dubium | Middle Norian |
Stubensandstein |
Many remains have since been attributed to other animals or given their own genera |  | ||
| Valid | Carnian |
"Pre-Tecovas Horizon" (Dockum Group) |
||||
| Nomen dubium | Norian | Named from several teeth in 1856 | ||||
| Valid | Norian | |||||
| Valid | Ladinian |
Falang Formation |
Possible basalmost taxon. |  | ||
| Valid | Late Carnian | |||||
| Nomen dubium | Late Carnian | |||||
| Valid | Carnian |
Tecovas Formation and unknown formation (Dockum Group) |
||||
| Valid | Carnian-Norian |  | ||||
| ? Valid | Triassic, ? early Olenekian |
? Middle Buntsandstein |
||||
| Valid | Middle Norian |
Stubensandstein |
||||
| Nicrosaurus | Valid | Late Norian - Rhaetian |
Arnstadt Formation |
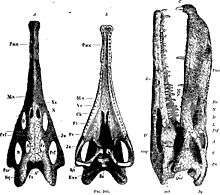 | ||
| Junior synonym | Late Carnian |
Hassberge Formation |
Polyphyletic. | |||
| Valid | Late Carnian - Early Norian |  | ||||
| Valid | Norian | |||||
| Valid | ||||||
| Valid | Norian |  | ||||
| Junior synonym | Junior synonym of Machaeroprosopus | |||||
| Valid | Norian - Rhaetian | |||||
| Valid | Carnian |  | ||||
| Valid | Carnian | |||||
| Valid | Carnian-Norian | |||||
Phylogeny
Phytosaurs are generally regarded as the most basal group of Crurotarsi, a clade of archosaurs that includes crocodilians and their extinct relatives.[18][19][20] Phytosaurs are often excluded from a clade called Suchia, which usually encompasses all other crurotarsans, including aetosaurs, rauisuchians, and crocodylomorphs.[20] Some studies have found polytomies between phytosaurs and other groups, like Ornithosuchidae and Suchia. In these cases, it is unclear whether phytosaurs are the most basal crurotarsans.[21] In one of the earliest studies of crurotarsan phylogeny, Sereno and Arcucci (1990) found Crurotarsi to be a monophyletic grouping consisting of phytosaurs, ornithosuchids, and the more derived suchians, but produced a trichotomy between the three groups in their tree.[22] In resolving this trichotomy, Parrish (1993) placed ornithosuchids, not phytosaurs, as the most basal crurotarsans. However, most other studies, such as Sereno (1991) and Benton et al. (2010), recover phytosaurs in a basalmost position among crurotarsans.[23] Below is a cladogram modified from Benton et al. (2010) showing the widely accepted phylogenetic relationships of phytosaurs:[20]
| Archosauria |
| ||||||||||||||||||||||||||||||
A phylogenetic analysis of early archosaurs by paleontologist Sterling Nesbitt (2011) found strong support for a sister taxon relationship between phytosaurs and Archosauria.[24] If this is the case, phytosaurs would be placed outside Pseudosuchia in a more basal position among archosauriforms. Phytosaurs would be considered closely related to the ancestors of both crocodilians and dinosaurs. Furthermore, the definition of the clade Crurotarsi would change, as it is often defined by the inclusion of phytosaurs. Thus, Crurotarsi would include phytosaurs and all other archosaurs —including dinosaurs— under this phylogeny.[25] Below is a cladogram showing the placement of phytosaurs from Nesbitt (2011):[24]
| Archosauriformes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The phylogenetic analysis of Stocker (2010) placed Paleorhinus outside Phytosauridae as a basal phytosaur. Under this phylogeny, Phytosauridae and Phytosauria are not synonymous. Stocker also erected the clade Leptosuchomorpha for derived phytosaurs, including Leptosuchus and Smilosuchus.[26]
Ezcurra (2016) updated Nesbitt's analysis and found that Phytosauria was once again a group of basal pseudosuchian archosaurs. His study analyzed the ten phylogenetic traits which Nesbitt claimed were lacking in phytosaurs but not archosaurs, thus excluding phytosaurs from Pseudosuchia. Four of the traits (well-developed palatal processes of the maxilla which meet at the midline, an elongated cochlear recess, a tuber on the lateral side of the ulna, and a particular orientation of the calcaneal tuber) were confirmed to support Nesbitt's placement of Phytosauria. However, one of the ten traits was found in Euparkeria (an abducens nerve exit foramen only present in the prootic) and another was found in proterochampsians (a swollen biceps tubercule), so their lack in phytosaurs may be reversals rather than basal traits. Another one of the traits (an antorbital fossa contacting the horizontal process of the maxilla) was found in the basal phytosaur Parasuchus. One trait (short metacarpals compared to metatarsals) was difficult to analyze in any crurotarsan, and another (a medial tuber on the femur) was found in both proterochampsids and Parasuchus. One trait (a divided tibial facet of the astragalus) was also lacking in Marasuchus and Nundasuchus, and therefore had a variable existence in Archosauria. This reanalysis, along with the observance of many traits linking Phytosauria with pseudosuchians, concluded that it was more likely that phytosaurs were pseudosuchians than non-archosaur archosauriforms. The following cladogram is a simplified version the fourth strict reduced consensus tree of Ezcurra's third phylogenetic analyses within his study. This cladogram only shows taxa from the group Eucrocopoda.[27]
| Eucrocopoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Footnotes
- Mateus, O., Clemmensen L., Klein N., Wings O., Frobøse N., Milàn J., Adolfssen J., & Estrup E. (2014). The Late Triassic of Jameson Land revisited: new vertebrate findings and the first phytosaur from Greenland. Journal of Vertebrate Paleontology. Program and Abstracts, 2014, 182.
- Hungerbühler, A (2002). "The Late Triassic phytosaur Mystriosuchus westphali, with a revision of the genus". Palaeontology. 45 (2): 377–418. doi:10.1111/1475-4983.00242.
- Stocker, M. R. & Butler, R. J. 2013. Phytosauria. Geological Society, London, Special Publications 379, 91-117.
- "Abstract," Abler (2001). Page 84.
- Padian, K., Li, C., & Pchelnikova, J. 2010. The trackmaker of (Late Triassic, North America): implications for the evolution of archosaur stance and gait. Palaeontology 53, 175-189.
- Klein, H. & Lucas, S. G. 2013. The Late Triassic tetrapod ichnotaxon Apatopus lineatus (Bock, 1952) and its distribution. Bulletin of the New Mexico Museum of Natural History 61, 313-324.
- Kimmig, J. 2013. Possible secondarily terrestrial lifestyle in the European phytosaur Nicrosaurus kapfii (Late Triassic, Norian): a preliminary study. Bulletin of the New Mexico Museum of Natural History and Science 61, 306-312.
- https://academic.oup.com/zoolinnean/article-abstract/187/1/198/5487160?redirectedFrom=fulltext
- Gozzi, E.; Renesto, S.A. (2003). "Complete specimen of Mystriosuchus (Reptilia, Phytosauria) from the Norian (Late Triassic) of Lombardy (Northern Italy)". Rivista Italiana di Paleontologia e Stratigrafia. 109 (3): 475–498.
- Renesto, S.A.; Lombardo, C. (1999). "Structure of the tail of a phytosaur (Reptilia, Archosauria) from the Norian (Late Triassic) of Lombardy (Northern Italy)". Nota Breve, Rivista Italiana di Paleontologia e Stratigrafia. 105 (1): 135–144.
- Neural and endocranial anatomy of Triassic phytosaurian reptiles and convergence with fossil and modern crocodylians Article in PeerJ 4(7):e2251 · July 2016 DOI: 10.7717/peerj.2251
- Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and Their Kin Capa Sterling J. Nesbitt, Julia Brenda Desojo, Randall B. Irmis Geological Society of London, 2013 - 608 páginas
- https://www.tandfonline.com/doi/full/10.1080/02724634.2019.1726361
- Michelle R. Stoker; Sterling J. Nesbitt; Li-Jun Zhao; Xiao-Chun Wu; Chun Li (2016). "Mosaic evolution in Phytosauria: the origin of long-snouted morphologies based on a complete skeleton of a phytosaur from the Middle Triassic of China". Society of Vertebrate Paleontology 76th Annual Meeting Program & Abstracts: 232.
- "Relationships of the Indian phytosaur Parasuchus hislopi Lydekker, 1885", Article · July 2015 DOI: 10.1002/spp2.1022
- Paul M. Barrett and Xu Xing (2012). "The enigmatic reptile Pachysuchus imperfectus Young, 1951 from the Lower Lufeng Formation (Lower Jurassic) of Yunnan, China" (PDF). Vertebrata PalAsiatica. 50 (2): 151–159.
- Maisch and Kapitzke (2010).
- Benton, M.J. (1999). "Scleromochlus taylori and the origin of dinosaurs and pterosaurs" (PDF). Philosophical Transactions of the Royal Society B: Biological Sciences. 354 (1388): 1423–1446. doi:10.1098/rstb.1999.0489. PMC 1692658.
- Nesbitt, S.J. (2007). "The anatomy of Effigia okeeffeae (Archosauria, Suchia), theropod-like convergence, and the distribution of related taxa" (PDF). Bulletin of the American Museum of Natural History. 302: 1–84. doi:10.1206/0003-0090(2007)302[1:TAOEOA]2.0.CO;2. hdl:2246/5840.
- Brusatte, S.L.; Benton, M.J.; Desojo, J.B.; Langer, M.C. (2010). "The higher-level phylogeny of Archosauria (Tetrapoda: Diapsida)" (PDF). Journal of Systematic Palaeontology. 8 (1): 3–47. doi:10.1080/14772010903537732.
- Gower, D.J.; Wilkinson, M. (1996). "Is there any consensus on basal archosaur phylogeny?". Proceedings of the Royal Society B: Biological Sciences. 263 (1375): 1399–1406. doi:10.1098/rspb.1996.0205.
- Sereno, P.C.; Arcucci, A.B. (1990). "The monophyly of crurotarsal archosaurs and the origin of bird and crocodile ankle joints". Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen. 180: 21–52.
- Sereno, P.C. (1991). "Basal archosaurs: phylogenetic relationships and functional implications". Journal of Vertebrate Paleontology. 11 (4, Supplement): 1–53. doi:10.1080/02724634.1991.10011426.
- Nesbitt, S.J. (2011). "The early evolution of archosaurs: relationships and the origin of major clades" (PDF). Bulletin of the American Museum of Natural History. 352: 1–292. doi:10.1206/352.1. hdl:2246/6112.
- Parker, Bill (27 April 2011). "Dinosaurs Are Crurotarsans". Chinleana. Blogger. Retrieved 27 April 2011.
- Michelle R. Stocker (2010). "A new taxon of phytosaur (Archosauria: Pseudosuchia) from the Late Triassic (Norian) Sonsela Member (Chinle Formation) in Arizona, and a critical reevaluation of Leptosuchus Case, 1922". Palaeontology. 53 (5): 997–1022. doi:10.1111/j.1475-4983.2010.00983.x.CS1 maint: uses authors parameter (link)
- Ezcurra, Martín D. (2016-04-28). "The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms". PeerJ. 4: e1778. doi:10.7717/peerj.1778. ISSN 2167-8359. PMC 4860341. PMID 27162705.
Sources
- Abler, W.L. 2001. A kerf-and-drill model of tyrannosaur tooth serrations. p. 84-89. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
- Carroll, R.L. (1988). Vertebrate Paleontology and Evolution, WH Freeman & Co.
- Chatterjee, S. (1978). A primitive parasuchid (phytosaur) reptile from the Upper Triassic Maleri Formation of India, Palaeontology 21: 83-127
- Hungerbühler, A. (2002). The Late Triassic phytosaur Mystriosuchus Westphali, with a revision of the genus. Palaeontology 45 (2): 377-418
- Jaeger, G.F. 1828. Über die fossilen Reptilien, welche in Würtemberg aufgefunden worden sind. Metzler, Stuttgart.
- Kimmig, J. & Arp, G. (2010) Phytosaur remains from the Norian Arnstadt Formation (Leine Valley, Germany), with reference to European phytosaur habitats. Palaeodiversity 3: 215-224
- Lucas, S.G. (1998). Global Triassic tetrapod biostratigraphy and biochronology. Paleogeog. Palaeoclimatol., Palaeoecol. 143: 347-384.
- Maisch, M.W.; Kapitzke, M. (2010). "A presumably marine phytosaur (Reptilia: Archosauria) from the pre-planorbis beds (Hettangian) of England". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 257 (3): 373–379. doi:10.1127/0077-7749/2010/0076.
- Olsen, P.E., Kent, D.V., H.-D.Sues, Koeberl, C., Huber, H., Montanari, E.C.Rainforth, A., Fowell, S.J., Szajna, M.J., and Hartline, B.W., (2002). Ascent of dinosaurs linked to an iridium anomaly at the Triassic-Jurassic boundary. Science 296: 1305-1307.
- Stocker, Michelle R. (2010). "A new taxon of phytosaur (Archosauria: Pseudosuchia) from the Late Triassic (Norian) Sonsela Member (Chinle Formation) in Arizona, and a critical reevaluation of Leptosuchus, Case, 1922". Palaeontology. 53 (5): 997–1022. doi:10.1111/j.1475-4983.2010.00983.x.CS1 maint: uses authors parameter (link)
- Ballew, K.L. (1989). A phylogenetic analysis of Phytosauria from the Late Triassic of the Western United States. Dawn of the age of dinosaurs in the American Southwest: pp. 309–339.
- Gregory, J.T. (1962). Genera of phytosaurs. American Journal of Science, 260: 652-690.
- Long, R.A. & Murry, P.A. (1995). Late Triassic (Carnian and Norian) tetrapods from the southwestern United States. New Mexico Museum of Natural History and Science Bulletin, 4: 1-254.
External links
- Translation and Pronunciation Guide
- A Preliminary Biomechanical Analysis of Phytosaur Life Habits
- Phytosauria - Palaeos
- Mikko's Phylogeny - cladistic tree
- Great Triassic Assemblages Pt 1 - The Chinle and Newark - some material on phytosaurs
| Wikimedia Commons has media related to Phytosauria. |