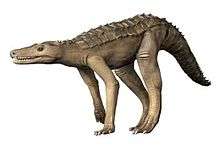
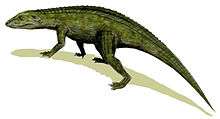
Aetosaur
Aetosaurs (aetosaur /eɪˌɛtoʊˈsɔːr/; order name Aetosauria /eɪˌɛtoʊˈsɔːriə/; from Greek, ἀετός (aetos, "eagle") and σαυρος (sauros, "lizard")) are an extinct order of heavily armoured, medium- to large-sized Late Triassic herbivorous archosaurs. They have small heads, upturned snouts, erect limbs, and a body covered by plate-like scutes. All aetosaurs belong to the family Stagonolepididae. Two distinct subdivisions of aetosaurs are currently recognized, Desmatosuchinae and Aetosaurinae, based primarily on differences in the morphology of the bony scutes of the two groups.[1] Over 20 genera of aetosaurs have been described, and recently there has been controversy regarding the description of some of these genera.
| Aetosaurs | |
|---|---|
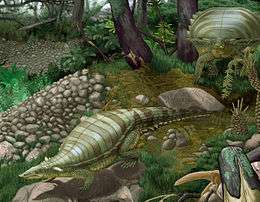 | |
| Two Typothorax in their environment | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Suchia |
| Order: | †Aetosauria Lydekker, 1887 |
| Subgroups | |
| |
Aetosaur fossil remains are known from Europe, North and South America, parts of Africa and India. Since their armoured plates are often preserved and are abundant in certain localities, aetosaurs serve as important Late Triassic tetrapod index fossils. Many aetosaurs had wide geographic ranges, but their stratigraphic ranges were relatively short. Therefore, the presence of particular aetosaurs can accurately date a site that they are found in.[2][3]
Aetosaur remains have been found since the early 19th century, although the very first remains that were described were mistaken for fish scales. Aetosaurs were later recognized as crocodile relatives, with early paleontologists considering them to be semiaquatic scavengers. They are now considered to have been entirely terrestrial animals. Some forms have characteristics that may have been adaptations to digging for food. There is also evidence that some if not all aetosaurs made nests.
Description
The head is small relative to the large body, and quite distinctive in shape, being flat and blunt at the front, like the snout of a pig. Some teeth are chisel-shaped, small and leaf-like, indicating a probable herbivorous diet,[4] although peg-like teeth and a keratinous snout have been considered possible adaptations in some species for feeding on colonial insects.[5][6] A study of the braincase of Stagonolepis robertsoni has shown that there are similarities between it and those of crocodylomorphs, which may indicate a close relationship.[7]
Aetosaurs had a "pillar-erect" erect limb posture similar to that seen in Rauisuchia, a related group of Triassic archosaurs.[2] A pillar-erect limb posture is one where the femur articulates vertically with the acetabulum of the hip, which is angled downward, so that the leg is positioned beneath the body and acts as a pillar bearing weight. While the limb posture is similar to rauisuchians, the feet resemble those of phytosaurs (crocodile-like semiaquatic crurotarsans) in the retention of primitive characteristics.[4] Although the forelimbs are much smaller than the hind limbs, all aetosaurs were quadrupeds.

Aetosaurs were very heavily armored (most certainly as a defense against predators), with large quadrangular, interlocking bony plates, or osteoderms, protecting the back and sides, belly, and tail.[4] Most osteoderms are heavily pitted on their upper surfaces and smooth on their undersides. Their centers are made of cancellous or spongy bone (also called diploë) and their outer portions are made up of compact bone.[8] In life, these plates were probably covered in horn.[9] Dorsal osteoderms, which are found on the backs of aetosaurs, are often ornamented with radial grooves. Dorsal paramedians, those found along the midline of the animal, are often wide and quadrangular with a small boss called a dorsal eminence on the dorsal surface of each plate. In aetosaurs, paramedian plates often have raised or depressed anterior edges where the plates articulate with the ones in front of them. If the anterior edge is raised, the area is called an anterior bar, while if it is depressed, the area is called an anterior lamina. In lateral plates, which are positioned on either side of the paramedian plates, the dorsal eminence is often enlarged into a prominent spike.[10] This spike is especially noticeable in desmatosuchines such as Longosuchus and Desmatosuchus.[11] Osteoderms are useful in diagnosing aetosaur taxa, and aetosaur species can often be identified from individual scutes based on their ornamentation pattern.[12][13]
.jpg)
Primitive genera, like the widespread Norian genus Aetosaurus and the Carnian Coahomasuchus, tended to be small, about a metre (3.2 ft) in length.[14] However, more advanced forms were larger - about 3 metres (9.8 ft) in length - with some taxa, such as Typothorax and Paratypothorax, possessing broad turtle-like bodies, and others, like Desmatosuchus, a narrow-bodied genus at least 4 metres (13 ft) long, equipped with large spines over the shoulders, which added to the animal's defensive armament.[15]
History
Aetosaur material was first described by Swiss paleontologist Louis Agassiz in 1844. He named the genus Stagonolepis from the Lossiemouth Sandstone in Elgin, Scotland, but considered it to be a Devonian fish rather than a Triassic reptile. This may be because he considered the strata to be part of the Old Red Sandstone, and thus Paleozoic in age. Agassiz mistook the osteoderms for large rhomboidal scales, which he thought were arranged in a similar pattern to those of gars. He also thought that these supposed scales were very similar to those of the lobe-finned fish Megalichthys due to their large size.[16]
English biologist Thomas Henry Huxley reconsidered the fish scales described by Agassiz and considered them to belong to a crocodilian. He first proposed this to the Geological Society of London in 1858, and went into more detail in an 1875 paper in the society's quarterly journal. By this time, new material had been uncovered from Elgin that indicated that Stagonolepis was not a fish, but a reptile. However, Stagonolepis was still known primarily by scutes and imprints of scutes, many of which were not well preserved.[17]

More complete aetosaur remains were found from the Lower Stubensandtein of Germany in the 1870s. Among them were complete articulated skeletons of 22 aetosaurs. These specimens were found in a large sandstone outcrop near Stuttgart and were preserved together in an area less than 2 square metres in size. The animals were probably buried under lake sediment soon after they died, with the flow of water repositioning their bodies on the lake bed and putting them in close proximity to one another. In 1877, German paleontologist Oscar Fraas assigned these specimens to the newly erected genus Aetosaurus. Fraas named the genus after the skull's resemblance to the head of an eagle, with a narrow, elongate skull and a pointed snout.[18]
The first aetosaurs from North America were also being described at this time. American paleontologist Edward Drinker Cope named Typothorax in 1875 and Episcoposaurus in 1877, both of which were from New Mexico. However, Cope considered these genera to be phytosaurs, crocodile-like aquatic archosaurs that were also common in the Late Triassic. While new material referable to Typothorax has been found in recent years throughout the American southwest, Episcoposaurus is no longer considered valid. Cope, along with later paleontologists such as Friedrich von Huene, recognized that remains of the type species E. horridus actually belonged to Typothorax. Another species, E. haplocerus, was reassigned to Desmatosuchus in 1953.[19]

A third North American genus called Stegomus was named by Othniel Charles Marsh in 1896.[20] Marsh had a long-time rivalry with Cope that was made famous in the Bone Wars of the late 19th century, in which the two tried to out-compete one another in the field and in scientific literature. Unlike Cope's aetosaurs, Stegomus was found from the eastern United States in Connecticut. Marsh also recognized Stegomus as an aetosaur rather than a phytosaur in his initial description of the genus. Like Cope, many paleontologists tended to consider aetosaur scutes to belong to phytosaurs during this time period.[13]
Aetosauria was first named in 1889 by English naturalist Richard Lydekker and zoologist Henry Alleyne Nicholson.[21] They considered Aetosauria to be one of three suborders of the order Crocodilia, the other two being Parasuchia (a Triassic group) and Eusuchia (a group including all post-Triassic crocodiles). At the time, Cope considered Aetosauria to belong to Rhynchocephalia, an order of reptiles that includes the living tuatara. He also thought that the tightly fitting osteoderms, which were thought to be fused to the ribs, indicated that aetosaurs were transitional between rhynchocephalians and turtles.[21] Nicholson and Lydekker placed a single family within the suborder, Aëtosauridæ. They considered aetosaurs to be similar to living crocodilians apart from their elongated metatarsals, which were compared to those of dinosaurs. In fact, Marsh considered aetosaurs to be closely related to dinosaurs because both had similar metatarsals.[21]
Aetosaur material continued to be described into the early 20th century, with notable paleontologists such as Barnum Brown and Charles Camp collecting specimens.[22] However, aetosaur remains were still being confused with those of phytosaurs. During this time, aetosaurs were usually considered to be members of Pseudosuchia, a now obsolete group consisting of various Triassic archosaurs including phytosaurs. However, similarities were often observed between aetosaurs and Crocodilia, the group to which aetosaurs were initially referred in the 19th century. While the skull had many features in common with pseudosuchians, parts of the postcranial skeleton, in particular the scutes, seemed to be similar to those of crocodilians.[23]
Distribution
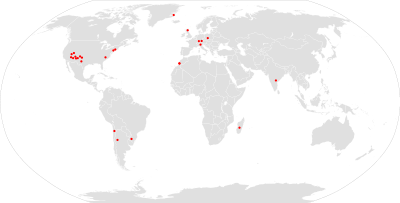
Aetosaur fossils have been found on all continents except Australia and Antarctica, giving them a nearly worldwide distribution during the Late Triassic. Many aetosaur remains have been found from the Chinle Group in the southwestern United States. Most fossils have been found from Arizona, New Mexico, and Texas.[24] Some remains have also been found from Utah, particularly in Canyonlands National Park and Zion National Park.[25] Aetosaurs are also known from the Newark Supergroup along the East Coast of the United States in states such as Connecticut and North Carolina.[26] Two genera are known from Greenland: Aetosaurus and Paratypothorax. They have been found from the Fleming Fjord Formation in Jameson Land.[27] Longosuchus is the only aetosaur known from Africa, with scutes having been found from the late Carnian Timesgadiouine Formation in Morocco.[28] During the Late Triassic, Morocco would have been in close proximity with the Newark Supergroup of North America in the supercontinent of Pangaea. It is also possible that Desmatosuchus was present in Africa, as fossils have been found from the Zarzaitine Series in Algeria that were referred to the genus.[14]
South American aetosaurs have been found from Argentina, Brazil, and Chile. In Argentina, they are known from the Carnian Ischigualasto Formation.[29] Common genera from Ischigualasto include Aetosauroides and Stagonolepis.[2] In Brazil, fossils have been found in the Santa Maria and Caturrita Formations in Rio Grande do Sul (Paleorrota).[14][29] Chilean aetosaurs are represented by one genus, Chilenosuchus, from the Antofagasta Region.[30] Aetosaurs have also been found from India, which, along with South America, was part of Gondwana during the Late Triassic. Early descriptions of aetosaurs included material from the Maleri Formation in south-central India, although it was too inadequate to assign specimens to any particular genus. Based on such descriptions, the Indian aetosaurs most closely resemble Longosuchus and Paratypothorax.[31] Reports of aetosaurs from Madagascar[32] are based on probable crocodylomorph scutes.[33]

Footprints belonging to the ichnogenus Brachychirotherium are often associated with aetosaurs.[34][35] Brachychirotherium has been found from Rio Grande do Sul in Paleorrota, Brazil[29] as well as Italy,[36] Germany,[37] and the eastern United States.[38] They are also common in the southwestern United States, having been found from Canyonlands National Park and Dinosaur National Monument.[25] Many of these tracks have a narrow gauge (meaning left and right prints are placed closely together) and nearly overstep each other. A 2011 functional analysis of the skeleton of Typothorax coccinarum indicated that it had the range of movement necessary to produce the tracks.[39]
Classification
Taxonomy
Aetosaurs belong to Pseudosuchia, a clade of archosaurs that includes living crocodilians and is characterized by the distinctive structure of the ankle bones. Aetosaurs were traditionally referred to a (now obsolete) group called the thecodonts, which included all "primitive" crocodilian relatives that lived in the Triassic. With the rise of phylogenetics, aetosaurs were later placed in a group called Suchia, which included many Triassic crurotarsans as well as later crurotarsans, including crocodilians.[40]
Originally, all aetosaurs were considered members of the family Stagonolepididae. Early phylogenetic analyses split aetosaurs into two subfamilies, Aetosaurinae and Desmatosuchinae. Aetosaurines are characterized by projections called eminences on the dorsal paramedian osteoderms that are close to the midline of the back. Desmatosuchines have a few more distinguishing characteristics, including grooves on the dorsal paramedians that help them lock to the lateral plates in a tight articulation. Many desmatosuchines have long spikes projecting from the lateral plates. These spikes are especially prominent in Desmatosuchus. Aetosaurines, on the other hand, tend to have less spikes. Many aetosaurines, such as Aetosaurus and Neoaetosauroides, have smooth carapaces and lack spikes altogether.[11] More recent studies (see below) have favored a third group, Typothoracisinae, which like Desmatosuchinae has long spikes, but differs in having more sharply angled joints between osteoderms. Moreover, the genus Aetosauroides is now often classified outside Stagonolepididae as a non-stagonolepidid aetosaur, making the names Aetosauria and Stagonolepididae no longer synonymous.[41]
Genera
| Genus | Status | Age | Location | Unit | Notes | Images |
|---|---|---|---|---|---|---|
| Valid | Norian |
Chinle Group |
||||
| Nomen dubium | Late Triassic | 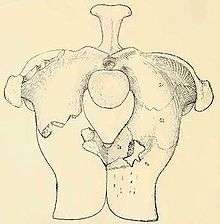 | ||||
| Valid | Carnian | |||||
| Valid | Carnian - Norian |
An aetosaur with aetosaurine-like osteoderms and desmatosuchine-like vertebrae |
||||
| Valid | Carnian - Norian |  | ||||
| Valid |
Lower Stubensandtein |
A small (1.5 m, 5 ft long) aetosaur with a wide geographic distribution in Europe | ||||
| Valid | late Norian - Rhaetian | |||||
| Nomen dubium | Carnian - Norian | |||||
| Valid | Late Triassic | |||||
| Valid | Late Triassic |
Was found in beds that were thought to be Carboniferous or Permian in age rather than Triassic |
||||
| Valid | Carnian | |||||
| Valid | Carnian - Norian | 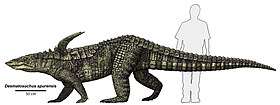 | ||||
| Valid | Late Triassic | Possible desmatosuchine characterized by cervical osteoderms that almost encircle the neck | ||||
| Junior synonym | Junior synonym of Rioarribasuchus | |||||
| Valid | Carnian |
Dockum Group |
 | |||
| Valid | Norian |
Bull Canyon Formation |
||||
| Valid | Norian |  | ||||
| Valid | Carnian - Rhaetian |
Lower Stubensandstein |
 | |||
| Valid | Norian | |||||
| Valid | Carnian | |||||
| Valid | Carnian | |||||
| Valid | Carnian |
Originally assigned to Desmatosuchus |
||||
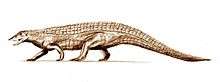
| Valid | Carnian |
Lossiemouth Sandstone |
 | |||
| Junior synonym | Junior synonym of Aetosaurus[26] | |||||
| Valid | Norian |
Originally assigned to Aetosaurus |
||||
|
Valid |
||||||
Phylogeny
|
|
Aetosaur phylogeny was first investigated in 1994 by paleontologist J. Michael Parrish. Aetosauroides, Aetosaurus, Desmatosuchus, Longosuchus, Neoaetosauroides, Stagonolepis, and Typothorax were included in the phylogenetic analysis. Aetosaurs were found to form a clade with rauisuchians, which Parrish termed Rauisuchiformes. Rauisuchiformes also included the superorder Crocodylomorpha, to which living crocodilians belong. Parrish found Aetosauria to be a monophyletic group and thus a true clade consisting of a common aetosaur ancestor and all of its descendants. To phylogenetically define Aetosauria, Parrish identified five synapomorphies, or shared characteristics. The first synapomorphy concerned the jaw, with the premaxilla at its tip being edentulous (toothless), upturned, and wide to form a "shovel". Moreover, the dentary bone in the lower jaw is also toothless, upturned, and broad. The reduced size and simple conical shape of the teeth was considered another synapomorphy. Two more synapomorphies of aetosaurs are shared with crocodylomorphs, but were not considered to be an indication of a close phylogenetic relationship; the body is covered in dorsal and ventral armor to form a complete carapace, and the paramedian osteoderms are much wider than they are long, with distinctive pitting. A final synapomorphy was found in the structure of the limb bones. In all aetosaurs, the limbs are very robust, with large muscle attachments such as the deltopectoral crest of the humerus, the fourth trochanter of the femur, the intracondylar ridge of the tibia, and the iliofibularis trochanter of the fibula.[42]
In Parrish's phylogenetic analysis, Aetosaurus was found to be the most basal member of the clade, the earliest to diverge after the most recent common ancestor. After Aetosaurus, there is a polytomy of three smaller clades in which it is unknown which clade diverged first from the group. Within this polytomy there was Neoaetosauroides, a clade containing Aetosauroides and Stagonolepis, and another polytomy that included Longosuchus, Desmatosuchus, and a clade containing Paratypothorax and Typothorax.
A later study by paleontologists Andrew B. Heckert and Spencer G. Lucas in 1999 expanded the number of synapomorphies that diagnose Aetosauria to 18. New synapomorphies included temporal fenestrae, or holes, that opened on the side of the skull rather than the top, lateral osteoderms articulating with the paramedians, and osteoderms covering the limbs. Aetosaurus was still found to be the most basal member, but the phylogeny of more derived aetosaurs differed in that Typothorax and Paratypothorax were split into two different clades with their sister taxa being Desmatosuchus and Longosuchus, respectively. More importantly, a new aetosaur called Coahomasuchus was included in the analysis. Coahomasuchus was found to be a basal aetosaur closely related to Stagonolepis, and also appeared early in the fossil record of aetosaurs. Previously, basal members were only known from later times, occurring after more advanced aetosaurs.[14]
In 2003, paleontologists Simon R. Harris, David J. Gower, and Mark Wilkinson examined previous phylogenetic studies of aetosaurs and criticized the way in which they used certain characters to produce cladograms. They concluded that only three hypotheses of aetosaur relationships from previous studies were still true: that Aetosaurus is the most basal aetosaur, that Aetosauroides is the sister taxon of Stagonolepis robertsoni, and that Longosuchus and Desmatosuchus are more closely related to each other than either is to Neoaetosauroides. They also went on to correct the trees from all previous analyses.[43]
More recently, a 2007 analysis by paleontologist William G. Parker resulted in a larger tree of aetosaur phylogenetics with the inclusion of Heliocanthus. Based on the tree, Parker defined the clades Typothoracisinae and Paratypothoracisini, both within Aetosaurinae. Parker also gave a revised phylogenetic definition of Aetosauria, mentioning that the previous definition, made by Heckert and Lucas in 2000, was somewhat ambiguous.[11] Heckert & Lucas (2000) defined Aetosauria as a stem-based taxon, claiming that Aetosauria included all crurotarsans that were more closely related to Desmatosuchus than to the immediate sister group of Aetosauria.[44] Because the immediate sister group of Aetosauria was uncertain, Parker offered a new definition with several non-aetosaur crurotarsan genera rather than one sister group. According to Parker, Aetosauria included all taxa more closely related to Aetosaurus and Desmatosuchus than to Leptosuchus, Postosuchus, Prestosuchus, Poposaurus, Sphenosuchus, Alligator, Gracilisuchus, and Revueltosaurus.[11]
A new genus of aetosaur, Aetobarbakinoides, was named in 2012. The phylogenetic analysis in that study found Aetosaurinae to be a paraphyletic grouping. As a paraphyletic group, aetosaurines would share a most recent common ancestor that is also the ancestor of other non-aetosaurine aetosaurs, and thus could not form their own clade.[41] Parker's 2007 analysis accepted this definition. In 2002, Heckert and Lucas defined Aetosaurinae as "a stem-based taxon containing all taxa more closely related to Aetosaurus than to the last common ancestor of Aetosaurus and Desmatosuchus".[11] The 2012 study placed Aetosaurus at the base of the stagonolepidid clade, with traditional aetosaurine taxa placed in successively more derived positions. In the analysis, these taxa are actually more closely related to Desmatosuchus than to Aetosaurus. Thus, under Heckert and Lucas's definition Aetosaurinae might be restricted to only Aetosaurus itself.
Another finding of this study was that Aetosauroides lies outside Stagonolepididae. If this phylogeny is correct, Stagonolepididae and Aetosauria would not be equivalent groupings, and Aetosauroides would be the first non-stagonolepidid aetosaur. The following cladogram simplified after an analysis presented by Julia B. Desojo, Martin D. Ezcurra and Edio E. Kischlat (2012).[41]
| Aetosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
In 2018, Parker conducted a new phylogenetic analysis with the re-description of Calyptosuchus. Below is the cladogram:[1]
| Aetosauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Evolution
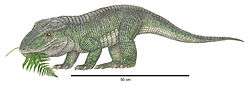
Although aetosaurs are known exclusively from the Late Triassic, their currently accepted position in archosaur phylogeny indicates that they originated from more basal pseudosuchian archosaurs in the Early or Middle Triassic. Given that aetosaurs are highly specialized with many anatomical features not seen in other pseudosuchians, the group's evolutionary origins are poorly understood. The recent discovery of complete specimens of the Late Triassic pseudosuchian Revueltosaurus callenderi indicate that it may have been close to the ancestry of aetosaurs. Several phylogenetic analyses place it as the sister taxon or closest relative of Aetosauria. Like aetosaurs, Revueltosaurus has two rows of paramedian osteoderms along its back and, in the cheek region of the skull, a maxilla that fits into a groove of the jugal bone.[8] One phylogenetic analysis places Turfanosuchus dabanensis, a Middle Triassic pseudosuchian, as the sister taxon of Revueltosaurus and Aetosauria, potentially making it the earliest known "stem aetosaur" ("stem" meaning that it lies on the branch that includes aetosaurs, but is not itself an aetosaur).[45] However, other studies have considered Turfanosuchus part of a different family of pseudosuchians, the gracilisuchids.[46]
In 2012 another "stem aetosaur" was described from the Middle Triassic Manda Beds of Tanzania. It differs from other Middle Triassic pseudosuchians in having a long skull, a small antorbital fenestra that fits into a large antorbital fossa in front of the eye socket, sharp and curved teeth, and osteoderms covering much of its body. Like aetosaurs and Revueltosaurus, it has a maxilla that fits into the jugal. Revueltosaurus, Turfanosuchus, and the unnamed Tanzanian pseudosuchian are all good fits for the hypothesized ancestor of aetosaurs because they both have double rows of leaf-shaped osteoderms along their backs that could potentially have evolved into the tightly fitting paramedian osteoderms of aetosaurs.[45]
Paleobiology
Early interpretations
In 1904, American paleontologist Henry Fairfield Osborn described aetosaurs as carnivorous aquatic animals of the order Parasuchia, mentioning that "[Parasuchia] constitutes an independent order, probably freshwater, littoral, carnivorous, short snouted (Aëtosaurus) or long snouted (Phytosaurus, Mystriosuchus) forms, analogous in their habits to the modern Crocodilia".[47] Early aetosaur remains were often found in clays beside skeletons of aquatic animals such as phytosaurs and terrestrial animals such as dinosaurs and trilophosaurs. This may have led some paleontologists to believe that the animals had died in swampy environments. Because there were a large number of skeletons of animals that would not normally have inhabited swamps in these clays, some paleontologists even suggested that aetosaurs scavenged off the carcasses of animals that became trapped in the swamps and died. Doubts were later raised over this lifestyle, since aetosaur teeth show little indication of carnivory and the weight of the armor suggests that aetosaurs had "a passive mode of life." However, aetosaurs were still regarded as partly aquatic into the mid-20th century.[23]

Feeding
Aetosaurs were herbivores, likely feeding on ferns and seed ferns that were common in the Triassic. The upturned shape of their snout suggests that aetosaurs may have dug up roots and tubers.[48] Aetosaurs have several anatomical features that may have been adaptations to digging, including a short radius relative to the humerus (seen in many other digging tetrapods) and a large deltopectoral crest on the humerus that served as an attachment for muscles. Aetosaurs also have large hind feet, or pes, with large claws that were likely used for skratch-digging. One aetosaur, Typothorax, has an entepicondyle on the humerus, which is the origin of forearm pronator and manual flexor muscles often used in digging. Moreover, it has a large olecranon process on the ulna which projects backward past the elbow, giving a large area for the insertion of the triceps muscle.[35] While many studies have suggested that aetosaurs had a fossorial or burrowing lifestyle,[42] aetosaurs have few of the characteristics that fossorial animals have as adaptations to digging. Therefore, it is likely that aetosaurs were able to dig to some extent, possibly rooting for food, but were unable to burrow.[35]
While features of the limbs indicate that aetosaurs probably dug for food, features of the skull and teeth can indicate what kind of food they were eating. Aetosaurs have many derived features not seen in other crurotarsans, which indicate that they are adapted to a different diet. Unlike the sharp, recurved teeth of other triassic archosaurs, aetosaurs had simple, conical teeth. The tips of the jaws were edentulous, or toothless, and probably supported a beak. The teeth have very little wear, suggesting that aetosaurs did not consume stiff and tough plant material. It is more likely that they consumed non-abrasive vegetation such as soft leaves.[42]
Alternative theories have been proposed for the diet of aetosaurs. In 1947, H J Sawin proposed that the aetosaur Longosuchus was a scavenger based on the close proximity of some specimens to a large number of skeletons that were likely carcasses.[23] A 2009 study of the jaw biomechanics of the South American genus Neoaetosauroides suggested that the animal may have fed on larvae and insects without hard exoskeletons. This is because Neoaetosauroides lacks serrations or wear facets on the teeth and has a jaw leverage that is not designed for strong forces such as crushing and chopping. The study recognized that northern aetosaurs such as Desmatosuchus and Stagonolepis did have jaws that would have supported a strong musculature, and were likely better suited to eating plant material.[6]
Nests
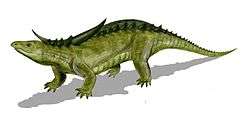
At least some aetosaurs built nests and protected their eggs.[49]
In 1996, geologist Stephen Hasiotis discovered 220‑million-year-old, fossilized, bowl-like pits in Arizona's Petrified Forest, in part of the Chinle Formation, assumed to be aetosaur and phytosaur nests. The "nests" are compacted and appear very similar to the nests of the modern day crocodiles who guard their nests.[48] However, it seems that these "nests" are instead the result of sandstone weathering.[50]
A second possible aetosaur nest site is known from northeastern Italy. The nests are preserved as depressions in carbonate rock that are circular or horseshoe-shaped, with high ridges around the sides. They appear to be unusually complex for nests created by Triassic reptiles. Archosaur footprints were found nearby that resembled aetosaurs, although they were not present in the same layer. Because the tracks were found so close to the nests, it is likely that aetosaurs built them.[49]
Growth
The ages of individual aetosaurs can be determined by examining their osteoderms. Some isolated osteoderms have been claimed to belong to juvenile aetosaurs based on their size and shape but these hypotheses have often been questioned. For example, juvenile osteoderms of Calyptosuchus were later identified as those of the small-bodied pseudosuchian Revueltosaurus (which is not an aetosaur), and juvenile osteoderms of Desmatosuchus have been reinterpreted as those of aetosaur Acaenasuchus, which had a relatively small body size at maturity. Studies of the bone structure of paramedian osteoderms indicate that new bone was deposited along the edges of each plate over the course of an aetosaur's lifetime. This means that lines of arrested growth on the undersides of paramedian osteoderms can be used to determine an individual's age.[8] Comparing the ages of individual specimens with their total body lengths indicates that aetosaurs increased in length at relatively constant rates, but increased in body mass at different rates depending on whether they had wide bodies like Typothorax or narrow bodies like Aetobarbakinoides. Aetosaurs also seem to have grown more slowly than modern crocodilians.[51] Analysis of the limb bones of aetosaurs indicates that they grew quickly when young and more slowly when adults. This pattern of growth is seen in most other pseudosuchians.[8]
Biochronology
Because species of aetosaurs typically have restricted fossil ranges and are abundant in the strata they are found in, they are useful in biochronology. Osteoderms are the most common remains associated with aetosaurs, so a single identifiable scute can accurately date the layer it is found in.
One aetosaur, Typothorax coccinarum, has been used to define the Revueltian land vertebrate faunachron. A land vertebrate faunachron (LVF) is a time interval that is defined by the first appearance datum (FAD), or first occurrence, of a tetrapod index fossil and is commonly used to date Late Triassic and Early Jurassic terrestrial strata.[54][55] Since the FAD of T. coccinarum is at the beginning of the Norian stage, the Revueltian LVF starts at the beginning of the Norian around 216 million years ago. The Revueltian ends with the next FAD, which happens to be that of the phytosaur Redondasaurus and the start of the Apachean LVF.[3][56]
Biochrons for aetosaur genera have been developed for dating strata in the Chinle Group of the southwestern United States. Up to 13 genera of aetosaurs are known from the Chinle Group, with most occurring in multiple localities and over short time spans. In 1996, paleontologists Spencer G. Lucas and Andrew B. Heckert recognized five biochrons based on the presence of aetosaurs throughout the Chinle Group.[57] The number of biochrons grew to 11 in a 2007 study by Heckert and Lucas along with Adrian P. Hunt and Justin A. Spielmann. These biochrons occurred from the Otischalkian LVF to the Apachean LVF and included genera such as Longosuchus, Tecovasuchus, and Typothorax.[53]
Naming controversy
In 2007, paleontologists at the New Mexico Museum of Natural History and Science in Albuquerque, New Mexico were accused of plagiarism in some of their published articles dealing with aetosaurs. In late 2006, the genus Rioarribasuchus was erected as a replacement name for "Desmatosuchus" chamaensis in the museum's bulletin.[58] However, four years earlier paleontologist William Parker reassigned "D." chamaensis to the newly named genus Heliocanthus in an unpublished thesis.[59] Because the name was not published, it was considered a nomen nudum until 2007 when it was described in the Journal of Systematic Palaeontology.[11] The authors of the 2006 paper, Spencer G. Lucas, Adrian P. Hunt, and Justin A. Spielmann, were accused of "intellectual theft" by paleontologists Jeff Martz, Mike Taylor, Matt Wedel, and Darren Naish, who claimed that they knew that Parker was eventually going to redescribe the species and formally erect a new genus. According to Martz, Taylor, Wedel, and Naish, the authors rushed to publish their own name before Parker could publish his.[60]
Another controversy occurred after Spielmann, Hunt, and Lucas published a 2006 paper mentioning that the holotype of Redondasuchus was not a left paramedian but a right one.[61] In 2002, Martz came to the same conclusion in an unpublished thesis.[62] He, along with Taylor, Wedel, and Naish, claimed that this was another form of plagiarism.[60]
These allegations were brought to the attention of the Ethics Education Committee of the Society of Vertebrate Paleontology (SVP) in 2007, and a response was given in 2008. In regard to Redondasuchus, the SVP found no plagiarism involved, while in the case of Heliocanthus and Rioarribasuchus, the SVP did not try to resolve the issue.[63]
The entire controversy came to be known as "Aetogate", in reference to the famous Watergate scandal of the 1970s.[60] It received wide attention from local Albuquerque newspapers and science blogs.[64] It was also the focus of a news article in a 2008 issue of the journal Nature.[65]
References
- Parker, W. G.; Stocker, M. R.; Irmis, R. B. (2008). "A new desmatosuchine aetosaur (Archosauria; Suchia) from the Upper Triassic Tecovas Formation (Dockum Group) of Texas". Journal of Vertebrate Paleontology. 28 (2): 692–701. doi:10.1671/0272-4634(2008)28[692:ANDAAS]2.0.CO;2. ISSN 0272-4634.
- Heckert, A. B.; Lucas, S. G. (2002). "South American occurrences of the Adamanian (Late Triassic: Latest Carnian) index taxon Stagonolepis (Archosauria: Aetosauria) and their biochronological significance". Journal of Paleontology. 76 (5): 852–863. doi:10.1666/0022-3360(2002)076<0852:SAOOTA>2.0.CO;2. ISSN 0022-3360.
- Lucas, S. G. (1998). "Global Triassic tetrapod biostratigraphy and biochronology". Palaeogeography, Palaeoclimatology, Palaeoecology. 143 (4): 347–384. CiteSeerX 10.1.1.572.872. doi:10.1016/S0031-0182(98)00117-5.
- Carroll, Robert L. (1988). Vertebrate Paleontology and Evolution. W.H. Freeman and Company. ISBN 978-0-7167-1822-2.
- Small, B. J. (2002). "Cranial anatomy of Desmatosuchus haplocerus (Reptilia: Archosauria: Stagonolepididae)". Zoological Journal of the Linnean Society. 136 (1): 97–111. doi:10.1046/j.1096-3642.2002.00028.x.
- Desojo, J.B.; Vizcaíno, S.F. (2009). "Jaw biomechanics in the South American aetosaur Neoaetosauroides engaeus". Paläontologische Zeitschrift. 83 (4): 499–510. doi:10.1007/s12542-009-0032-6.
- Gower, D. J.; Walker A. D. (2002). "New data on the braincase of the aetosaurian archosaur (Reptilia: Diapsida) Stagonolepis robertsoni Agassiz". Zoological Journal of the Linnean Society. 136 (1): 7–23. doi:10.1046/j.1096-3642.2002.00023.x.Archosaurian Anatomy and Palaeontology: Essays in Memory of Alick D. Walker, DB Norman & DJ Gower (eds.)
- Desojo, J. B.; Heckert, A. B.; Martz, J. W.; Parker, W. G.; Schoch, R. R.; Small, B. J.; Sulej, T. (2013). "Aetosauria: A clade of armoured pseudosuchians from the Upper Triassic continental beds". Geological Society, London, Special Publications. 379: 203–239. doi:10.1144/SP379.17.
- Colbert, E. H. (1969). Evolution of the Vertebrates (2nd ed.). John Wiley & Sons. ISBN 978-0-471-38461-8.
- Bill Parker (August 12, 2009). "Aetosaurs 101: Osteoderm Nomenclature." Chinleana. Accessed January 8, 2010.
- Parker, W. G. (2007). "Reassessment of the aetosaur Desmatosuchus chamaensis with a reanalysis of the phylogeny of the Aetosauria (Archosauria:Pseudosuchia)". Journal of Systematic Palaeontology. 5: 41–68. doi:10.1017/S1477201906001994.
- Long, R. A.; Ballew K. L. (1985). "Aetosaur dermal armor from the late Triassic of southwestern North America, with special reference to material from the Chinle Formation of Petrified Forest National Park". Museum of Northern Arizona Bulletin. 47: 45–68.
- Parker, Bill (13 February 2009). "The Aetosaur Paper that Changed Everything". Chinleana. Blogspot. Retrieved 8 January 2010.
- Heckert, A. B.; Lucas S. G. (1999). "A new aetosaur (Reptilia: Archosauria) from the Upper Triassic of Texas and the phylogeny of aetosaurs". Journal of Vertebrate Paleontology. 19 (1): 50–68. CiteSeerX 10.1.1.563.9516. doi:10.1080/02724634.1999.10011122.
- Parker, W.G. (2005). "A new species of the Late Triassic aetosaur Desmatosuchus (Archosauria: Pseudosuchia)". Comptes Rendus Palevol. 4 (4): 327–340. doi:10.1016/j.crpv.2005.03.002.
- Agassiz, L. (1844). Monographie des poisons fossils du Vieux Grés Rouge ou Systéme Dévonien (Old Red Sandstone) des Iles Britanniques ed de Russie. Neuchâtel: Jent et Gassman. p. 171.
- Huxley, T.H. (1875). "On Stagonolepis Robertsoni, and on the evolution of the Crocodilia". Quarterly Journal of the Geological Society. 31 (1–4): 423–438. doi:10.1144/GSL.JGS.1875.031.01-04.29. hdl:2027/hvd.32044107172884.
- Fraas, O. (1877). "Aetosaurus ferratus Fr. Die gepanzerte Vogel-Echse aus dem Stubensandstein bei Stuttgar". Festschrift zur Feier des 400jährigen Jubiläums der Eberhard-Karls-Universät zu Tübingen, Wurttembergische Naturwissenschaftliche Jahreshefte. 33 (3): 1–22.
- Gregory, J.T. (1953). "Typothorax and Desmatosuchus" (PDF). Postilla, Peabody Museum of Natural History Bulletin (16): 1–27. Archived from the original (PDF) on 2010-06-27.
- Marsh, O.C. (1896). "A new belodont reptile (Stegomus) from the Connecticut River Sandstone". American Journal of Science. 2 (7): 59–62. doi:10.2475/ajs.s4-2.7.59.
- Nicholson, H.A.; Lydekker, R. (1889). "Order Crocodilia". A manual of palaeontology for the use of students, with a general introduction on the principles of palaeontology. II. Nature. pp. 1180–1196.
- Parker, Bill (9 August 2009). "Who Has Worked on Aetosaurs?". Chinleana. Blogspot. Retrieved 24 July 2010.
- Sawin, H.J. (1947). "The pseudosuchian reptile Typothorax meadei". Journal of Paleontology. 21 (3): 201–238.
- Heckert, A.B.; Spielmann, J.A.; Lucas, S.G.; Hunt, A.P. (2007). "Biostratigraphic utility of the Upper Triassic aetosaur Tecovasuchus (Archosauria:Stagonolepididae), an index taxon of St. Johnsian (Adamanian:Late Carnian) time" (PDF). In Lucas, S.G.; Spielmann, J.A. (eds.). The Global Triassic. New Mexico Museum of Natural History and Science Bulletin. 41. pp. 51–57.
- Santucci, V.L.; Kirkland, J.I. (2010). "An overview of National Park Service paleontological resources from the parks and monuments in Utah" (PDF). In Sprinkel, D.A.; Chidsey, T.C. Jr.; Anderson, P.B. (eds.). Geology of Utah's Parks and Monuments. 2010 Utah Geological Association Publication 28 (3rd ed.). pp. 589–623.
- Lucas, S.G.; Heckert, A.B.; Huber, P. (1998). "Aetosaurus (Archosauromorpha) from the Upper Triassic of the Newark Supergroup, eastern United States, and its biochronological significance" (PDF). Palaeontology. 41 (6): 1215–1230. Archived from the original (PDF) on 2012-03-09.
- Jenkins, F.A.; Shubin, N.H.; Amaral, W.W.; Gatesy, S.M.; Schaff, C.R.; Clemmensen, L.B.; Downs, W.R.; Davidson, A.R.; Bonde, N.; Osbaeck, F.F. (1994). "Late Triassic continental vertebrates and depositional environments of the Fleming Fjord Formation, Jameson Land, East Greenland". Meddelelser om Grønland, Geoscience. 32: 1–25.
- Lucas, S.G. (1998). "The aetosaur Longosuchus from the Triassic of Morocco and its biochronological significance". Comptes Rendus de l'Académie des Sciences, Série IIA. 326 (8): 589–594. doi:10.1016/S1251-8050(98)80211-5.
- Silva, R.C. Da; Carvalho, I. De S.; Schwanke, C. (2007). "Vertebrate dinoturbation from the Caturrita Formation (Late Triassic, Paraná Basin), Rio Grande do Sul State, Brazil" (PDF). Gondwana Research. 11 (2007): 303–310. doi:10.1016/j.gr.2006.05.011. Archived from the original (PDF) on 2011-07-06.
- Desojo, J.B. (2001). "Redescripcion del aetosaurio Chilenosuchus forttae Casamiqueal (Diapsida: Arcosauria): presencia de Triasico continental en el norte de Chile" (PDF). Revista Geológica de Chile. 30 (1): 53–63. doi:10.4067/s0716-02082003000100004.
- Heckert, A.B.; Lucas, S.G.; Hunt, A.P.; Spielmann, J.A. (2007). "Late Triassic aetosaur biochronology revisited" (PDF). In Lucas, S.G.; Spielmann, J.A. (eds.). The Global Triassic. New Mexico Museum of Natural History and Science Bulletin. 41. pp. 49–50.
- Lucas, S.G.; Heckert, A.P. (2002). "The Hyperodapedon biochron, Late Triassic of Pangea" (PDF). Albertiana. 27: 30–38.
- Parker, William G. (2008). "Description of new material of the aetosaur Desmatosuchus spurensis (Archosauria: Suchia) from the Chinle Formation of Arizona and a revision of the genus Desmatosuchus". PaleoBios. 28: 1–40.
- Hunt, A.P.; Lockley, M.G.; Lucas, S.G. (1993). "Vertebrate and invertebrate tracks and trackways from Upper Triassic Strata of the Tucumcari Basin, East-Central New Mexico, U.S.A.". In Lucas, S.G.; Morales, M. (eds.). The Nonmarine Triassic. New Mexico Museum of Natural History and Science Bulletin. 3. pp. 199–201.
- Heckert, A.B.; Lucas, S.G.; Rinehart, L.F.; Celesky, M.D.; Spielmann, J.A.; Hunt, A.P. (2010). "Articulated skeletons of the aetosaur Typothorax coccinarum Cope (Archosauria: Stagonolepididae) from the Upper Triassic Bull Canyon Formation (Revueltian: early-mid Norian), eastern New Mexico, USA" (PDF). Journal of Vertebrate Paleontology. 30 (3): 619–642. doi:10.1080/02724631003763524.
- Avanzini, M. (2000). "Synaptichnium tracks with skin impressions from the Anisian (middle Triassic) of the Southern alps (val di non - Italy)". Ichnos. 7 (4): 243–251. doi:10.1080/10420940009380164.
- Haubold, H. (1983). "Archosaur evidence in the Buntsandstein (Lower Triassic)" (PDF). Acta Palaeontologica Polonica. 28 (1–2): 123–132.
- Olsen, P.E.; Baird, D. (1988). "The ichnogenus Atriepus and its significance for Triassic biostratigraphy". In Padian, K. (ed.). The Beginning of the Age of Dinosaurs: Faunal Change Across the Triassic-Jurassic Boundary. Cambridge: Cambridge University Press. pp. 61–88. ISBN 978-0-521-36779-0.
- Lucas, S.G.; Heckert, A.B. (2011). "Late Triassic aetosaurs as the trackmaker of the tetrapod footprint ichnotaxon Brachychirotherium". Ichnos. 18 (4): 197–208. doi:10.1080/10420940.2011.632456.
- Brusatte, S.L.; Benton, M.J.; Desojo, J.B.; Langer, M.C. (2010). "The higher-level phylogeny of Archosauria (Tetrapoda: Diapsida)" (PDF). Journal of Systematic Palaeontology. 8 (1): 3–47. doi:10.1080/14772010903537732.
- Julia B. Desojo; Martin D. Ezcurra; Edio E. Kischlat (2012). "A new aetosaur genus (Archosauria: Pseudosuchia) from the early Late Triassic of southern Brazil" (PDF). Zootaxa. 3166: 1–33. doi:10.11646/zootaxa.3166.1.1. ISSN 1175-5334.
- Parrish, J.M. (1994). "Cranial osteology of Longosuchus meadei and the phylogeny and distribution of the Aetosauria". Journal of Vertebrate Paleontology. 14 (2): 196–209. doi:10.1080/02724634.1994.10011552.
- Harris, S.R.; Gower, D.J.; Wilkinson, M. (2002). "Intraorganismal homology, character construction, and the phylogeny of aetosaurian archosaurs (Reptilia, Diapsida)" (PDF). Systematic Biology. 52 (2): 239–52. doi:10.1080/10635150309341. PMID 12746149.
- Heckert, A. B.; Lucas S. G. (2000). "Taxonomy, phylogeny, biostratigraphy, biochronology, paleobiogeography, and evolution of the Late Triassic Aetosauria (Archosauria: Crurotarsi)". Zentralblatt für Geologie und Paläontologie, Teil I. 11-12: 1539–1587.
- Nesbitt, S.J.; Sidor, C.A.; Angielczyk, K.D.; Smith, R.M.; Parker, W. (2012). "Derivation of the aetosaur osteoderm carapace: evidence from a new, exceptionally preserved "stem aetosaur" from the Middle Triassic (Anisian) Manda Beds of southwestern Tanzania". Journal of Vertebrate Paleontology. 32 (Supp. 1): 149. doi:10.1080/02724634.2012.10635175.
- Butler, Richard J.; Sullivan, Corwin; Ezcurra, Martín D.; Liu, Jun; Lecuona, Agustina; Sookias, Roland B. (2014-06-10). "New clade of enigmatic early archosaurs yields insights into early pseudosuchian phylogeny and the biogeography of the archosaur radiation". BMC Evolutionary Biology. 14 (1): 128. doi:10.1186/1471-2148-14-128. ISSN 1471-2148. PMC 4061117. PMID 24916124.
- Osborn, H.F. (1904). "Reclassification of the reptilia". The American Naturalist. 38 (446): 93–115. doi:10.1086/278383.
- Ziehn, K.; Sangalang, E.; Fritz-Laylin, L.; Twu, C.; Huynh, A.; Smith, D. (1 May 2000). "Introduction to the Aetosauria". University of California Museum of Paleontology - History of life through time. Regents of the University of California. Archived from the original on 24 September 2010. Retrieved 18 August 2010.
- Avanzini, M.; Dalla vecchia, F.M.; Mietto, P; Piubelli, D; Preto, N; Rigo, M; Roghi, G (2007). "A vertebrate nesting site in northeastern Italy reveals unexpectedly complex behavior for late Carnian reptiles". PALAIOS. 22 (5): 465–475. doi:10.2110/palo.2005.p05-137r.
- Anatomy, Phylogeny and Palaeobiology of Early Archosaurs and Their Kin Capa Sterling J. Nesbitt, Julia Brenda Desojo, Randall B. Irmis Geological Society of London, 2013 - 608 páginas
- Taborda, J. R. A.; Cerda, I. A.; Desojo, J. B. (2013). "Growth curve of Aetosauroides scagliai Casamiquela 1960 (Pseudosuchia: Aetosauria) inferred from osteoderm histology". Geological Society, London, Special Publications. 379: 413–423. doi:10.1144/SP379.19.
- Lucas, S.G.; Heckert, A.B. & Huber, P. (1998). "Aetosaurus (Archosauromorpha) from the Upper Triassic of the Newark Supergroup, eastern United States, and its biochronological significance" (PDF). Palaeontology. 41 (6): 1215–1230.
- Heckert, A.B.; Lucas, S.G.; Hunt, A.P. & Spielmann, J.A. (2007). "Late Triassic aetosaur biochronology revisited" (PDF). In Lucas, S.G. & Spielmann, J.A. (eds.). The Global Triassic. New Mexico Museum of Natural History and Science Bulletin. 41. pp. 49–50.
- Lucas, S.G. (1991). "Sequence stratigraphic correlation of nonmarine and marine Late Triassic biochronologies, western United States". Albertiana. 9: 11–18.
- Lucas, S.G.; Hunt, A.P. (1993). "Tetrapod biochronology of the Chinle Group (Upper Triassic), western United States". In Lucas, S.G.; Morales, M. (eds.). The Nonmarine Triassic. New Mexico Museum of Natural History and Science Bulletin. 3. pp. 327–329.
- Lucas, S.G.; Hunt, A.P.; Heckert, A.B.; Spielmann, J.A. (2007). "Global Triassic tetrapod biostratigraphy and biochronology: 2007 status" (PDF). In Lucas, S.G.; Spielmann, J.A. (eds.). The Global Triassic. New Mexico Museum of Natural History and Science Bulletin. 41. pp. 229–240.
- Lucas, S.G.; Heckert, A.B. (1996). "Late Triassic aetosaur biochronology". Albertiana. 17: 57–64.
- Lucas, S.G.; Hunt, A.P.; Spielmann, J.A. (2006). "Rioarribasuchus, a new name for an aetosaur from the Upper Triassic of north-central New Mexico". In Harris; et al. (eds.). The Triassic-Jurassic Terrestrial Transition. New Mexico Museum of Natural History and Science Bulletin. Albuquerque: New Mexico Museum of Natural History and Science. p. 37.
- Parker, W.G. (2003). Description of a new specimen of Desmatosuchus haplocerus from the Late Triassic of Northern Arizona (M.Sc. thesis). Flagstaff: Northern Arizona University. p. 315.
- Martz, Jeff; Taylor, Mark; Wedel, Matt; Naish, Darren (1 June 2008). "Timeline of publications and letters". Aetogate: asking for answers in New Mexico. Retrieved 15 August 2010.
- Spielmann, J.A.; Hunt, A.P.; Lucas, S.G.; Heckert, A.B. (2006). "Revision of Redondasuchus (Archosauria: Aetosauria) from the Upper Triassic Redonda Formation, New Mexico, with description of a new species" (PDF). In Harris, H.D.; Lucas, S.G.; Spielmann, J.A.; Lockley, M.G.; Milner, A.R.C.; Kirkland, J.L. (eds.). The Triassic-Jurassic Terrestrial Transition. 37. New Mexico Museum of Natural History and Science Bulletin. pp. 583–587.
- Martz, J.W. (2002). The morphology and ontogeny of Typothorax coccinarum (Archosauria Stagonolepididae) from the Upper Triassic of the American Southwest (PDF) (M.Sc. thesis). Texas Tech University. Retrieved 3 July 2010.
- "Statement from the Executive Committee about the allegations of unethical conduct from J. Martz, W. Parker, M. Taylor and M. Wedel against S. Lucas, A. Hunt, A. Heckert, and J. Spielmann" (PDF). Society of Vertebrate Paleontology. 2008. Archived from the original (PDF) on 2010-07-14. Cite journal requires
|journal=(help) - Martz, Jeff; Taylor, Mark; Wedel, Matt; Naish, Darren (17 February 2010). "Press Coverage". Aetogate: asking for answers in New Mexico. Retrieved 15 August 2010.
- Dalton, R. (2008). "Fossil reptiles mired in controversy". Nature. 451 (7178): 510. doi:10.1038/451510a. PMID 18235465.
General references
- Benton, M. J. (2000). Vertebrate Paleontology, 2nd ed. Blackwell Science Ltd.
External links
| Wikimedia Commons has media related to Aetosaur. |
| Wikispecies has information related to Aetosaur |
- Archosauromorpha: Suchia (Aetosauridae) - Palaeos.com
- Aetosauria Translation and Pronunciation Guide - Ben Creisler
- Introduction to the Aetosauria - University of California Museum of Paleontology
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||