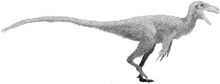
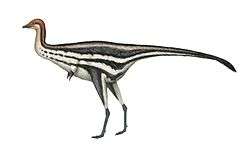
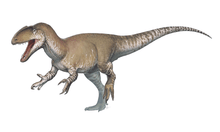Ornithomimosauria
The Ornithomimosauria, ornithomimosaurs ("bird-mimic lizards") or ostrich dinosaurs[7] are theropod dinosaurs which bore a superficial resemblance to the modern-day ostrich. They were fast, omnivorous or herbivorous dinosaurs from the Cretaceous Period of Laurasia (now Asia, Europe and North America), as well as Africa and possibly Australia.[8] The group first appeared in the Early Cretaceous and persisted until the Late Cretaceous. Primitive members of the group include Nqwebasaurus, Pelecanimimus, Shenzhousaurus, Hexing and Deinocheirus, the arms of which reached 2.4 m (8 feet) in length. More advanced species, members of the family Ornithomimidae, include Gallimimus, Struthiomimus, and Ornithomimus. Some paleontologists, like Paul Sereno, consider the enigmatic alvarezsaurids to be close relatives of the ornithomimosaurs and place them together in the superfamily Ornithomimoidea (see classification below).
| Ornithomimosaurs | |
|---|---|
 | |
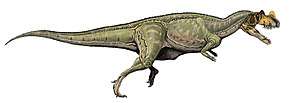
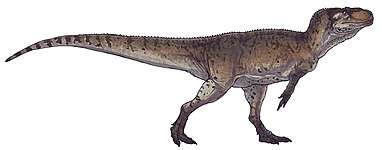
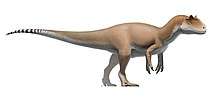
| Collection of four ornithomimosaurs. From top left to right: Gallimimus, Struthiomimus, Ornithomimus and Anserimimus. | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Clade: | Maniraptoriformes |
| Clade: | †Ornithomimosauria Barsbold, 1976 |
| Subgroups[2] | |
| |
| Synonyms | |
| |
Description
The skulls of ornithomimosaurs were small, with large eyes, above relatively long and slender necks. The most basal members of the taxon (such as Pelecanimimus and Harpymimus) had a jaw with small teeth, while the later and more derived species had a toothless beak.[9]

The fore limbs ("arms") were long and slender and bore powerful claws. The hind limbs were long and powerful, with a long foot and short, strong toes terminating in hooflike claws. Ornithomimosaurs were probably among the fastest of all dinosaurs. Like other coelurosaurs, the ornithomimosaurian hide was feathered rather than scaly.
Palaeobiology
Ornithomimosaurs probably acquired most of their calories from plants. Many ornithomimosaurs, including primitive species, have been found with numerous gastroliths in their stomachs, characteristic of herbivores. Henry Fairfield Osborn suggested that the long, sloth-like "arms" of ornithomimosaurs may have been used to pull down branches on which to feed, an idea supported by further study of their strange, hook-like hands.[10] The sheer abundance of ornithomimids — they are the most common small dinosaurs in North America — is consistent with the idea that they were plant eaters, as herbivores usually outnumber carnivores in an ecosystem. However, they may have been omnivores that ate both plants and small animal prey.
Comparisons between the scleral rings of two ornithomimosaur genera (Garudimimus and Ornithomimus) and modern birds and reptiles indicate that they may have been cathemeral, active throughout the day at short intervals.[11]
Feathers
Unambiguous evidence of feathers is known from Ornithomimus edmontonicus, of which there are multiple specimens preserving feather traces.[12] Deinocheirus and Pelecanimimus have been speculated to be feathered as well, the former due to the presence of a pygostyle,[13] and the later due to possible impressions (otherwise taken to be collagen fibers).
There is a debate on whereas ornithomimids possessed the pennaceous feathers seen in Pennaraptora.[14] Otherwise, a very ostrich-like plumage and feather range is seen in known in one specimen.[15]
Classification
Named by O.C. Marsh in 1890, the family Ornithomimidae was originally classified as a group of "megalosaurs" (a "wastebasket taxon" containing any medium to large sized theropod dinosaurs), but as more theropod diversity was uncovered, their true relationships to other theropods started to resolve, and they were moved to the Coelurosauria. Recognizing the distinctiveness of ornithomimids compared to other dinosaurs, Rinchen Barsbold placed ornithomimids within their own infraorder, Ornithomimosauria, in 1976. The contents of Ornithomimidae and Ornithomimosauria varied from author to author as cladistic definitions began to appear for the groups in the 1990s.
In the early 1990s, prominent paleontologists such as Thomas R. Holtz, Jr. proposed a close relationship between theropods with an arctometatarsalian foot; that is, bipedal dinosaurs in which the upper foot bones were 'pinched' together, an adaptation for running. Holtz (1994) defined the clade Arctometatarsalia as "the first theropod to develop the arctometatarsalian pes and all of its descendants." This group included the Troodontidae, Tyrannosauroidea, and Ornithomimosauria. Holtz (1996, 2000) later refined this definition to the branch-based "Ornithomimus and all theropods sharing a more recent common ancestor with Ornithomimus than with birds." Subsequently, the idea that all arctometatarsalian dinosaurs formed a natural group was abandoned by most paleontologists, including Holtz, as studies began to demonstrate that tyrannosaurids and troodontids were more closely related to other groups of coelurosaurs than they were to ornithomimosaurs. Since the strict definition of Arctometatarsalia was based on Ornithomimus, it became redundant with the name Ornithomimosauria under broad definitions of that clade, and the name Arctometatarsalia was mostly abandoned.
The paleontologist Paul Sereno, in 2005, proposed the clade "Ornithomimiformes", defining them as all species closer to Ornithomimus edmontonicus than to Passer domesticus. Because he had redefined Ornithomimosauria in a much narrower sense, a new term was made necessary within his preferred terminology to denote the clade containing the sistergroups Ornithomimosauria and Alvarezsauridae — previously the latter had been contained within the former. However, this concept only appeared on Sereno's Web site and has not yet been officially published as a valid name.[16]
"Ornithomimiformes" was identical in content to Holtz's Arctometatarsalia, as it has a very similar definition. While "Ornithomimiformes" is the newer group, Sereno rejected the idea that Arctometatarsalia should take precedence, because the meaning of the former name has been changed very radically by Holtz.[16]
Phylogeny

Ornithomimosauria has variously been used for the branch-based group of all dinosaurs closer to Ornithomimus than to birds, and in more restrictive senses. The more exclusive sense began to grow in popularity when the possibility arose that alvarezsaurids might fall under Ornithomimosauria if an inclusive definition were adopted. Another clade, Ornithomimiformes, was defined by Sereno (2005) as (Ornithomimus velox > Passer domesticus) and replaces the more inclusive use of Ornithomimosauria when alvarezsaurids or some other group are found to be closer relatives of ornithomimosaurs than maniraptorans, with Ornithomimosauria redefined to include dinosaurs closer to Ornithomimus than to alvarezsaurids. Gregory S. Paul has proposed that Ornithomimosauria might be a group of primitive, flightless birds, more advanced than Deinonychosauria and Oviraptorosauria.[17]
The cladogram below follows an analysis by Yuong-Nam Lee, Rinchen Barsbold, Philip J. Currie, Yoshitsugu Kobayashi, Hang-Jae Lee, Pascal Godefroit, François Escuillié & Tsogtbaatar Chinzorig. The analysis was published in 2014, and includes many ornithomimosaurian taxa.[2]
| Coelurosauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The cladogram below follows an analysis by Jin Liyong, Chen Jun and Pascal Godefroit (2012).[6]
| Ornithomimosauria |
| ||||||||||||||||||||||||||||||
The cladogram presented here follows the one recovered by Turner, Clarke, Ericson and Norell, 2007.[18] Clade names follow definitions provided by Sereno, 2005.[19]
| Ornithomimosauria |
| ||||||||||||||||||||||||||||||||||||||||||
See also
References
- Holtz, Thomas R. Jr. (2012) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix.
- Lee, Y.-N.; Barsbold, R.; Currie, P.J.; Kobayashi, Y.; Lee, H.-J.; Godefroit, P.; Escuillié, F.; Chinzorig, T. (2014). "Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus". Nature. 515 (7526): 1–4. Bibcode:2014Natur.515..257L. doi:10.1038/nature13874. PMID 25337880.
- Brownstein CD. (2016) Redescription of Arundel formation Ornithomimosaur material and a reinterpretation of Nedcolbertia justinhofmanni as an "Ostrich Dinosaur": Biogeographic implications. PeerJ Preprints 4:e2308v1 https://doi.org/10.7287/peerj.preprints.2308v1
- Choiniere, J. N.; Forster, C. A.; De Klerk, W. J. (2012). "New information on Nqwebasaurus thwazi, a coelurosaurian theropod from the Early Cretaceous (Hauteriverian?) Kirkwood Formation in South Africa". Journal of African Earth Sciences. 71–72: 1–17. Bibcode:2012JAfES..71....1C. doi:10.1016/j.jafrearsci.2012.05.005.
- Sereno, P. (2017). "Early Cretaceous ornithomimosaurs (Dinosauria: Coelurosauria) from Africa". Ameghiniana. 54 (5): 576–616. doi:10.5710/AMGH.23.10.2017.3155.
- Jin Liyong, Chen Jun & Pascal Godefroit (2012). "A New Basal Ornithomimosaur (Dinosauria: Theropoda) from the Early Cretaceous Yixian Formation, Northeast China". In Godefroit, P. (ed.). Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems. Indiana University Press. pp. 467–487.
- "Ostrich dinosaurs".
- Choiniere, Jonah N.; Forster, Catherine A.; De Klerk, William J. (2012). "New information on Nqwebasaurus thwazi, a coelurosaurian theropod from the Early Cretaceous Kirkwood Formation in South Africa". Journal of African Earth Sciences. 71–72: 1–17. Bibcode:2012JAfES..71....1C. doi:10.1016/j.jafrearsci.2012.05.005.
- Last of the Dinosaurs: The Cretaceous Period
- Nicholls and Russell (1985).
- Schmitz and Motani (2011)
- van der Reest, A.J.; Wolfe, A.P.; Currie, P.J. (2016). "A densely feathered ornithomimid (Dinosauria: Theropoda) from the Upper Cretaceous Dinosaur Park Formation, Alberta, Canada". Cretaceous Research. 58: 108–117. doi:10.1016/j.cretres.2015.10.004.
- Lee, Yuong-Nam; Barsbold, Rinchen; Currie, Philip J.; Kobayashi, Yoshitsugu; Lee, Hang-Jae; Godefroit, Pascal; Escuillié, François; Chinzorig, Tsogtbaatar (2014). "Resolving the long-standing enigmas of a giant ornithomimosaur Deinocheirus mirificus". Nature. 515 (7526): 257–260. Bibcode:2014Natur.515..257L. doi:10.1038/nature13874. PMID 25337880.
- Foth, Christian; Tischlinger, Helmut; Rauhut, Oliver W. M. (2014). "New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers". Nature. 511 (7507): 79–82. Bibcode:2014Natur.511...79F. doi:10.1038/nature13467. PMID 24990749.
- Van Der Reest, Aaron J.; Wolfe, Alexander P.; Currie, Philip J. (2016). "A densely feathered ornithomimid (Dinosauria: Theropoda) from the Upper Cretaceous Dinosaur Park Formation, Alberta, Canada". Cretaceous Research. 58: 108–117. doi:10.1016/j.cretres.2015.10.004.
- Sereno, P. C. (2005). Stem Archosauria—TaxonSearch Archived 2009-01-15 at the Wayback Machine [version 1.0, 2005 November 7]
- Paul, G.S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press.
- Turner, et al. (2007).
- "Taxon Search: TaxonSearch Archive Page - Stem Archosauria 1.0". Archived from the original on 2009-01-15.
Further reading
- Barrett, P. M. (2005). "The diet of ostrich dinosaurs (Theropoda: Ornithomimosauria)". Palaeontology. 48 (2): 347–358. doi:10.1111/j.1475-4983.2005.00448.x.
- British Museum (Natural History): Ostrich Dinosaurs
- Jacobsen, A.R. 2001. Tooth-marked small theropod bone: An extremely rare trace. p. 58-63. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
- Li Xu; Yoshitsugu Kobayashi; Junchang Lü; Yuong-Nam Lee; Yongqing Liu; Kohei Tanaka; Xingliao Zhang; Songhai Jia; Jiming Zhang (2011). "A new ornithomimid dinosaur with North American affinities from the Late Cretaceous Qiupa Formation in Henan Province of China". Cretaceous Research. 32 (2): 213–222. doi:10.1016/j.cretres.2010.12.004.
- Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 337-363.
- Nicholls, E. L.; Russell, A. P. (1985). "Structure and function of the pectoral girdle and forelimb of Struthiomimus altus (Theropoda: Ornithomimidae)". Palaeontology. 28: 643–677.
- Norell, M. A.; Makovicky, P.; Currie, P. J. (2001). "The beaks of ostrich dinosaurs". Nature. 412 (6850): 873–874. Bibcode:2001Natur.412..873N. doi:10.1038/35091139. PMID 11528466.
- Schmitz, L. & Motani, R. (2011). "Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology". Science. 332 (6030): 705–8. Bibcode:2011Sci...332..705S. doi:10.1126/science.1200043. PMID 21493820.
- Sereno, P. C. 2005. Stem Archosauria—TaxonSearch [version 1.0, 2005 November 7]
- Tanke, D.H. and Brett-Surman, M.K. 2001. Evidence of Hatchling and Nestling-Size Hadrosaurs (Reptilia:Ornithischia) from Dinosaur Provincial Park (Dinosaur Park Formation: Campanian), Alberta, Canada. pp. 206–218. In: Mesozoic Vertebrate Life—New Research Inspired by the Paleontology of Philip J. Currie. Edited by D.H. Tanke and K. Carpenter. Indiana University Press: Bloomington. xviii + 577 pp.
- Turner, A.H.; Pol, D.; Clarke, J.A.; Erickson, G.M.; Norell, M. (2007). "Supporting online material for: A basal dromaeosaurid and size evolution preceding avian flight". Science. 317 (5843): 1378–1381. Bibcode:2007Sci...317.1378T. doi:10.1126/science.1144066. PMID 17823350. (supplement)
External links
| Wikimedia Commons has media related to Ornithomimosauria. |
| Wikispecies has information related to Ornithomimosauria |