Ornithocheiridae
Ornithocheiridae (or ornithocheirids, meaning "bird hands") is a group of pterosaurs within the suborder Pterodactyloidea. These pterosaurs were among the last to possess teeth. Members that belong to this group lived from the Early to Late Cretaceous periods (Valanginian to Cenomanian stages), around 140 to 96 million years ago.
| Ornithocheirids | |
|---|---|
 | |
| Restored skeleton of Tropeognathus mesembrinus in the National Museum of Brazil | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Order: | †Pterosauria |
| Suborder: | †Pterodactyloidea |
| Clade: | †Anhangueria |
| Clade: | †Ornithocheirae |
| Family: | †Ornithocheiridae Seeley, 1870 |
| Type species | |
| †Pterodactylus simus Owen, 1861 | |
| Genera | |
| |
| Synonyms | |
| |
Ornithocheirids are generally infamous for having an enormously controversial and very confusing taxonomy. Although agreements that these animals were related, and therefore similar to istiodactylids and pteranodontians, there is still no virtual consensus over the exact content and interrelationships of this group.[1] Ornithocheirids were the most successful pterosaurs during their reign, they were also the largest pterosaurs before the appearance of the azhdarchids such as Quetzalcoatlus. Ornithocheirids were excellent fish hunters, they used various flight techniques to catch their prey, and they are also capable of flying great distances without flapping constantly. The later existence of the crested pteranodontids made paleontologists think that ornithocheirids were their potential ancestors. This is due to their very similar aspects such as the different flying techniques, the long distance flights, and most of all their diet, which consists mainly on fish.
History
Naming
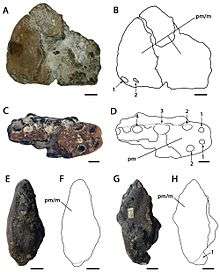
The family Ornithocheiridae is without a doubt, one of the most well-known pterosaur groups, this is mostly due to their very controversial and convoluted taxonomic history. Most of the ornithocheirid fossil record consists of isolated teeth, as well as fragmentary bones, reaching hundreds or even thousands of remains in some localities.[1] The first uncovered ornithocheirid remains were described in 1861 by British paleontologist Sir Richard Owen; he assigned the fossil remains to a new species of Pterodactylus: P. simus.[2] In 1869, British paleontologist Harry Govier Seeley erected the new generic name Ornithocheirus (from Ancient Greek meaning "bird hand"), and assigned P. simus as its type species, therefore creating Ornithocheirus simus. Later, in 1870, Seeley created the name Ornithocheirae to only contain Ornithocheirus.[3] However, in the same year, this was emended to Ornithocheiridae by Seeley himself following the article 11.7.1.3 of the ICZN.[4] In 1874, Owen had proposed two new genera for the Cretaceous British pterosaurs: Coloborhynchus (meaning "maimed beak") and Criorhynchus (meaning "ram beak") based on highly distinctive jaw fragments.[5] Owen reassigned P. simus as the type species of Criorhynchus, creating Criorhynchus simus. He referred three species to Coloborhynchus, including the addition of a new species called C. clavirostris; no type species was designated however.[6] In 1876 however, Seeley pointed out that Criorhynchus was a junior synonym of Ornithocheirus, a concept that was followed by paleontologist Richard Lydekker in 1888.[5] In the latter year, Lydekker acknowlegded that Ornithocheirus simus was the type species of Ornithocheirus, he also distiguished O. simus by its tall rostrum, while other species referred to Ornithocheirus had lanceolate jaw tips. Therefore, to avoid confusion, Lydekker preferred to use the name Criorhynchus for O. simus, and Ornithocheirus for the species with lanceolate jaw tips, this concept was later favored by paleontologist Reginald Walter Hooley in 1914. In his review of Ornithocheirus, he divided the family Ornithocheiridae into two subfamilies: Ornithocheirinae and Criorhynchinae; the former consisted of Ornithocheirus and Lonchodectes, while the latter consisted of Amblydectes and Criorhynchus.[7] In his review, Hooley also considered the species Coloborhynchus clavirostris as a synonym of Criorhynchus simus.[4][8] In 1967, paleontologist Oskar Kuhn placed Criorhynchus within the family Criorhynchidae (which is a term now considered synonymous to Ornithocheiridae), and also recognized Ornithocheirus within the family Ornithocheiridae and subfamily Ornithocheirinae. He also designated the species Coloborhynchus clavirostris as the type species of Coloborhynchus, but agreed with Hooley that it was synonymous with Criorhynchus simus.[6][4] In 1994 however, Yuong-Nam Lee revalidated the genus Coloborhynchus (with C. clavirostris as its type species), and regarded it as distinct from Criorhynchus simus.[6] Later, in 2001, paleontologist David Unwin revised the taxonomic history of the Cambridge Greensand pterosaurs, and divided Ornithocheiridae into three genera: Ornithocheirus, Coloborhynchus and Anhanguera. Unwin also designated Pterodactylus simus as the type and only species of Ornithocheirus.[4]
In 2003, Unwin defined the family Ornithocheiridae as Haopterus gracilis, Ornithocheirus simus, their most recent common ancestor, and all its descendants. He included the genera Anhanguera, Brasileodactylus, Coloborhynchus, Haopterus, Ludodactylus and Ornithocheirus within the family, and also concluded that Araripesaurus, Arthurdactylus and Santanadactylus may belong to this family as well. However, their taxonomic status and precise relationships with other ornithocheirids are uncertain.[9] In 2019, upon the description of the pterosaur Mimodactylus, Haopterus, which was assigned to this family by Unwin,[9] and recovered as a basal eupterodactyloid by Brian Andres and colleagues,[10] was reassigned by Alexander Kellner and colleagues as the sister taxon of the former.[11]
In 2014, Andres and colleagues defined the Ornithocheiridae with a different definition: the most inclusive clade containing Ornithocheirus simus but not Anhanguera blittersdorffi. They placed the genera Coloborhynchus, Ornithocheirus and Tropeognathus within the Ornithocheiridae, while placing Anhanguera within the separate family Anhangueridae. However, back in 2001, Unwin considered the name Anhangueridae a junior synonym of Ornithocheiridae,[5] a concept that was later followed by several paleontologists such as Mark Witton in 2013.[1] Later phylogenetic analyses however, contradict this name synonymy, with Ornithocheiridae and Anhangueridae classified as different families, therefore following the 2014 analysis by Andres and colleagues instead.[12][13][14]
Description

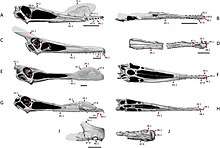
Ornithocheirids were among the most successful and widely distributed pterosaurs, these animals were characterized by long jaws that had spike-like teeth. Wingspans of smaller ornithocheirids such as Ferrodraco measured approximately 4 meters (13 ft),[14][13] while wingspans of giant morphs reached 8 meters (26 ft) or more.[13] Specimen NHMUK R481, a specimen that belongs to the species Coloborhynchus capito, the largest toothed pterosaur, had wingspan that may have reached 7 meters (23 ft).[15] However, in 2013, a specimen referred to the genus Tropeognathus (MN 6594-V) was calculated to have had a normal wingspan of 8.26 meters (27.1 ft), while a calculated maximum winspan reached 8.70 meters (28.5 ft), meaning that this specimen clearly shows that wingspans of toothed pterosaurs could exceed 7 meters (23 ft).[16]
Skull
Ornithocheirids had elongated jaws with rounded sagittal crests on both tips, as well as robust fang-like teeth.[13][17] The sagittal crest of the species Ornithocheirus simus and Tropeognathus mesembrinus extended to the anterior end of the rostrum, a feature that is also seen in Siroccopteryx moroccensis; further synapomorphies between these three species were also found, including the premaxilla having a tall and narrow shape in anterior aspect, the anterolateral margins of the premaxilla being convex in both anterior and lateral view, a feature that resulted in a bluntly rounded outline of the tip of the rostrum. The rostrum in S. moroccensis lacks a constriction that is posterior to the anterior rosette, a feature also shared by O. simus and T. mesembrinus, therefore another synapomorphy supported by these three species. Yet another feature shared by these three species is that the teeth are short, straight, and relatively uniform in size, something that is not present in other ornithocheirans such as Coloborhynchus and Anhanguera.[13] In Coloborhynchus, the teeth were found to have been heterodont, elongated, recurved and caniniform, which is similar to those seen in another ornithocheirid called Caulkicephalus.[18] The genus Caulkicephalus, though having similarities with other ornithocheirids, including the anterior end of the rostrum being transversely expanded, or having a low, bony sagittal crest that includes a smooth dorsal margin on the rostrum, still possesses some unique features. The most distinct characteristic of Caulkicephalus is that it bores a frontoparietal crest, a feature that is only seen in pteranodontians such as Pteranodon, and in Ludodactylus,[19] a pterosaur once assigned to the Ornithocheiridae, but many recent analysis however, have placed it within the more inclusive group Anhangueria, outside the Ornithocheiridae.[12][14]
Postcranial skeleton

According to many studies, the wing-membranes of the ornithocheirids were extensive and broad, resulting in a high aspect ratio, analyses also show that most ornithocheirids had low wing loadings, which led to an easier maneuver during flight.[20] Ornithocheirid wings mostly differed from earlier pterosaur wings because of their impressive size, and the wings weren't only capable of soaring high, but also soaring very long distances without flapping.[21] These pterosaurs also bore proportionally enormous forelimbs; around five times stronger than their legs, and much stronger in biomechanical testing.[22]
Studies of the pelvis of several ornithocheirids show that they were mostly of moderate size compared to the body as a whole, which is similar to the structures in other ornithocheiroids. The pelvic bones, which in total were three, were fused, this is seen in many related genera such as Anhanguera. The ilium was long and low, and its front and rear blades projected horizontally beyond the edges of the lower pelvic bones.[23] Given the structure length, the processes of these rod-like forms indicate that the hindlimb muscles attached to them with limited strength.[24] An ischiopubic blade is seen in most ornithocheirids, where the pubic bone is fused with the broad ischium, resulting in a narrow build.[22] The front of the pubic bones were also articulated with a unique structure, resulting in a pair of prepubic bones within, this formed a cusp covering the rear belly of the pterosaur, and was located between the pelvis and the belly ribs. This element suggests of a vertical mobility, and therefore had a function in breathing, with the chest cavity compensating in a relative rigidity.[25] Normally, the blades of ornithocheirids, most notably in the larger genera, were fused in both sides, closing the pelvis from below and forming the pelvic canal. Ornithocheirids didn't have the hip joint perforated, but directed obliquely upwards, preventing a perfectly vertical position of the leg, and therefore allowed considerable mobility for the hindlimbs.[26][22]
Vertebral columns and vertebrae of ornithocheirids are only known from scarce remains, and it is hard to determine how their posture were. In some genera such as Tropeognathus, the first five dorsal vertebrae are fused into a notarium, with five sacral vertebrae fused into a synsacrum, and the third and fourth sacral vertebrae are keeled within. Ornithocheirids also had a relatively long and straight neck. The more advanced species typically had their neck longer than their torso.[22]
Classification

Ornithocheirids were impressively large compared to other types of pterosaurs from the Early Cretaceous, and they were also one of the most successful of all pterosaurs.[27][15] Most ornithocheirids were also long considered to only have a piscivorous diet, while this is mostly true, some remains uncovered were suggested to belong to an ornithocheirid with a non-piscivorous diet.[28]
Other evolutionary changes of ornithocheirids include their territorial range, they were even present in the southern continent of Oceania, represented by two genera: Ferrodraco and Mythunga.[14][29][30] Most of the species discovered in Europe and South America belonged to Coloborhynchus and Ornithocheirus, with some currently being synonyms of Tropeognathus and Anhanguera.[6][17]
The genus Cimoliopterus, originally Pterodactylus cuvieri, once assigned to Ornithocheirus as O. cuvieri, and formerly classified within this family, was reclassified by Pêgas et al. as a targaryendraconian in 2019. Paleontologists found it to be closer to the newly named Targaryendraco (originally Ornithocheirus wiedenrothi), but as the sister taxon of Aetodactylus and Camposipterus, taxa that were also once classified within the Ornithocheiridae.[13][31] In the same year however, a phylogenetic analysis by Jacobs et al. had recovered both Cimoliopterus and Camposipterus within the Ornithocheiridae again, specifically placed as sister taxa.[13]
Synapomorphies found unique in Coloborhynchus made authors assign it to the family Ornithocheiridae.[13] However, some authors placed Coloborhynchus within the family Anhangueridae instead,[11] and with the later discovery of Uktenadactylus, which shared very similar features with Coloborhynchus, it was suggested that it would be its sister taxon.[6][13]
Phylogeny
The phylogenetic placement of the family Ornithocheiridae was conducted by Andres and Myers in 2013, where they placed the family as the sister taxon of Anhangueridae, and within the more inclusive group Ornithocheirae.[32] In 2019, a later analysis by Pentland et al. had included the subfamily Ornithocheirinae within the family, with the genera Ferrodraco, Mythunga, Ornithocheirus, and Coloborhynchus as the members. In the analysis, they also placed the genus Tropeognathus within the family, but in a basal position.[14]
|
Topology 1: Andres & Myers (2013).[32]
|
Topology 2: Pentland et al. (2019).[14]
|
Paleobiology
Feeding and diet

Unlike the related istiodactylids, ornithocheirids were mainly fish hunters, consisting on a piscivorous diet that varied depending on the species. Paleontologists suggest that the feeding habits of the earlier istiodactylids changed through the process of evolution, resulting in a more aquatic-based lifestyle. The studies and analysis of ornithocheirids made paleontologists conclude how they hunted or catched fish successfully.[33] The first basis is their long snouts, which they used for catching the fish near the surface of the water, paleontologists also suggest that they flew with moderated speed, giving them an overall advantage of a surprise attack during the hunt. Others suggest that ornithocheirids, similar to pteranodontids, just glided down directly to the water when catching prey, this technique can also be seen in modern-day soaring birds such as albatrosses.[34]
Teeth enamels
Some discoveries, found that several ornithocheirids weren't completely fish hunters or aquatic consumers. This is confirmed with a 2017 analysis of carbon isotopes found on teeth enamels of ornithocheirids. In the analysis, they compared the enamel values of ornithocheirids with the aquatic and terrestrial consumers near the fossil site.[28] The ornithocheirid enamels found in the Twin Mountains Formation were shown with lower enamel values compared to the aquatic consumers that lived nearby, suggesting a more terrestrial diet, similar to the carnivorous theropods found in the site, such as Acrocanthosaurus. Enamels found in the Paw Paw Formation however, were described with higher values, in comparison to the nearby terrestrial consumers. These enamel remains were analyzed, and paleontologists then found out that the values seen were more similar to aquatic consumer enamels rather the terrestrial ones.[28] These analyses are considered very useful reference, since there isn't much data on several ornithocheirid genera. The earlier species that partially fed on terrestrial prey, such as Coloborhynchus, were also analyzed, and paleontologists conclude that there can be a specific evolutionary change between the earlier and later formations, due to the age difference.[6] This can be demostrated with the enamel remains found in the Twin Mountains and Paw Paw formations. The ones found in the Twin Mountains Formation dated back to the earlier Aptian stage, while the remains found in the Paw Paw Formation dated back to the later Albian stage.[28]
Locomotion and flight

Similar to modern-day albatrosses, most ornithocheirids used a flight technique called "dynamic soaring", which consists of travelling long distances without flapping using the vertical gradient of wind speed near the ocean surface as an advantage, but the flight speed is moderate. Several studies showed that most ornithocheirids sprawled their limbs to a large degree, similar to crocodiles, while other studies however, conclude that ornithocheirids were generally quadrupedal. Yet other studies concluded that ornithocheirids held their limbs more or less vertically extended, similar to an avian- or mammal-like configuration.[24] Some studies in the later genera, show that ornithocheirids spend lots of their time sea fishing, and as a result, they perhaps influenced the later pteranodontids with the same piscivorous diet, as well as their well developed flight techniques.[35][36] Analyses of limb proportions in the genus Anhanguera however, show that some ornithocheirids were consistent on hopping, but the later genera were suggested that they most likely walked on four limbs, which consists on their wing-fingers as the front limbs, and using their hind limbs to balance.[37]
Paleoecology
Ornithocheirids were a widespreaded type of pterosaurs, with many fossil remains found across the world.[17] The first true ornithocheirid specimens were uncovered in the Cambridge Greensand of England, these belong to the infamous genus Ornithocheirus, and dated back to the Albian stage of the Early Cretaceous.[5] Within the fossil site, several other pterosaurs were also found, these include the ornithocheirids Amblydectes and Coloborhynchus,[6] the targaryendraconian Camposipterus, the lonchodectid Lonchodraco,[4] and the azhdarchoid Ornithostoma.[38] The ornithischians Anoplosaurus, Acanthopholis,[39] and the dubious Eucercosaurus and Trachodon were also found within the formation.[40] Fossil remains of the sauropod Macrurosaurus were also present.[41] The bird Enaliornis,[42] as well as the ichthyosaurs Cetarthrosaurus, Platypterygius and Sisteronia were also found alongside the remains of ornithocheirids.[43]
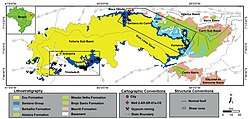
A Lagerstätte called the Santana Group (sometimes known as the Santana Formation) in northeastern Brazil was found to contain a large number of pterosaur genera. The most diverse formation of the group is the Romualdo Formation, known for its wide variety of pterosaur remains.[44] The formation dates back 111 to 108 million years ago, also during the Albian stage of the Early Cretaceous. The Romualdo Formation is found to contain a variety of ornithocheirids, including Tropeognathus,[16] Coloborhynchus[45] and Araripesaurus,[46] the targaryendraconian Barbosania,[47] and the anhanguerids Anhanguera and Maaradactylus were also found alongside.[48] The related Araripedactylus, Brasileodactylus,[49] Cearadactylus,[50] Santanadactylus[51] and Unwindia[52] were also present within the fossil site. Many other pterosaur were found within, these include the tapejarid Tapejara,[53] as well as the thalassodromids Thalassodromeus[54] and Tupuxuara.[55] Other animals such the theropods Irritator,[56] Mirischia[57] and Santanaraptor, as well as the crocodylomorph Araripesuchus were also found.[58] Several turtle remains were found within the formation, with some specimens referring to the genera Santanachelys, Cearachelys and Araripemys,[59][60] and along with these, many fish remains were also found, these were assigned to the genera Brannerion, Rhinobatos, Rhacolepis, Tharrhias and Tribodus.[61]
Ornithocheirids were also partially distributed in North America, and several specimens that were found are thought to belong to the genus Uktenadactylus (originally Coloborhychus wadleighi).[62] This pterosaur was uncovered in the Paw Paw Formation of Texas, United States, which dated back to the Albian and Cenomanian stages. The formation includes several ankylosaurian dinosaurs such as Pawpawsaurus, Texasetes, and an indetermine nodosaurid.[63][64][65] Within the fossil site, several specimens of ammonoids were thought to belong to the genera Turrilites and Scaphites,[66] and along with these, shark remains of Leptostyrax were also found.[67]
See also
References
- Witton 2013, p. 152.
- Martill, D.M. (2010). "The early history of pterosaur discovery in Great Britain". Geological Society of London, Special Publications. 343 (1): 287–311.
- Seeley, H.G. (1870). The Ornithosauria: an Elementary Study of the Bones of Pterodactyles. Cambridge. p. 113.
- Rodrigues, T.; Kellner, A. (2013). "Taxonomic review of the Ornithocheirus complex (Pterosauria) from the Cretaceous of England". ZooKeys (308): 1–112. doi:10.3897/zookeys.308.5559. PMC 3689139. PMID 23794925.
- Unwin, D.M. (2001). "An overview of the pterosaur assemblage from the Cambridge Greensand (Cretaceous) of Eastern England". Mitteilungen aus dem Museum für Naturkunde in Berlin, Geowissenschaftliche Reihe. 4: 189–221.
- Rodrigues, T. and Kellner, A.W.A. (2008). "Review of the pterodactyloid pterosaur Coloborhynchus". Flugsaurier: pterosaur papers in honour of Peter Wellnhofer. Zitteliana B. 28: 219–228.CS1 maint: uses authors parameter (link)
- Kellner AWA, Tomida Y (2000) Description of a new species of Anhangueridae (Pterodactyloidea) with comments on the pterosaur fauna from the Santana Formation (Aptian-Albian), northeastern Brazil. National Science Museum Monographs 17: 1-135.
- Hooley, Reginald Walter (1914). "On the Ornithosaurian genus Ornithocheirus, with a review of the specimens from the Cambridge Greensand in the Sedgwick Museum, Cambridge". Annals and Magazine of Natural History. 13 (78): 529–557. doi:10.1080/00222931408693521. ISSN 0374-5481.
- Unwin, D. M. (2003). "On the phylogeny and evolutionary history of pterosaurs". Geological Society, London, Special Publications. 217 (1): 139–190. Bibcode:2003GSLSP.217..139U. CiteSeerX 10.1.1.924.5957. doi:10.1144/GSL.SP.2003.217.01.11.
- Andres, B.; Clark, J.; Xu, X. (2014). "The Earliest Pterodactyloid and the Origin of the Group". Current Biology. 24: 1011–6. doi:10.1016/j.cub.2014.03.030. PMID 24768054.
- Kellner, Alexander W. A.; Caldwell, Michael W.; Holgado, Borja; Vecchia, Fabio M. Dalla; Nohra, Roy; Sayão, Juliana M.; Currie, Philip J. (2019). "First complete pterosaur from the Afro-Arabian continent: insight into pterodactyloid diversity". Scientific Reports. 9 (1). doi:10.1038/s41598-019-54042-z.
- Longrich, N.R., Martill, D.M., and Andres, B. (2018). "Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary." PLoS Biology, 16(3): e2001663. doi:10.1371/journal.pbio.2001663
- Jacobs, M.L., Martill, D.M., Ibrahim, N., Longrich, N. (2019). "A new species of Coloborhynchus (Pterosauria, Ornithocheiridae) from the mid-Cretaceous of North Africa" (PDF). Cretaceous Research. doi:10.1016/j.cretres.2018.10.018.CS1 maint: uses authors parameter (link)
- Pentland, Adele H.; Poropat, Stephen F.; Tischler, Travis R.; Sloan, Trish; Elliott, Robert A.; Elliott, Harry A.; Elliott, Judy A.; Elliott, David A. (December 2019). "Ferrodraco lentoni gen. et sp. nov., a new ornithocheirid pterosaur from the Winton Formation (Cenomanian–lower Turonian) of Queensland, Australia". Scientific Reports. 9 (1): 13454. doi:10.1038/s41598-019-49789-4. ISSN 2045-2322. PMC 6776501. PMID 31582757.
- Martill, D.M. and Unwin, D.M. (2011). "The world's largest toothed pterosaur, NHMUK R481, an incomplete rostrum of Coloborhynchus capito (Seeley 1870) from the Cambridge Greensand of England." Cretaceous Research, (advance online publication). doi:10.1016/j.cretres.2011.09.003
- Kellner, A. W. A.; Campos, D. A.; Sayão, J. M.; Saraiva, A. N. A. F.; Rodrigues, T.; Oliveira, G.; Cruz, L. A.; Costa, F. R.; Silva, H. P.; Ferreira, J. S. (2013). "The largest flying reptile from Gondwana: A new specimen of Tropeognathus cf. T. mesembrinus Wellnhofer, 1987 (Pterodactyloidea, Anhangueridae) and other large pterosaurs from the Romualdo Formation, Lower Cretaceous, Brazil". Anais da Academia Brasileira de Ciências. 85: 113–135. doi:10.1590/S0001-37652013000100009. PMID 23538956.
- Witton 2013, p. 155.
- Martill, David M. (2015), First occurrence of the pterosaur Coloborhynchus (Pterosauria, Ornithocheiridae) from the Wessex Formation (Lower Cretaceous) of the Isle of Wight, England., 126, 3, Proceedings of the Geologists' Association, pp. 377–380, retrieved 2016-10-26
- Steel, L., Martill, D.M., Unwin, D.M. and Winch, J. D. (2005). "A new pterodactyloid pterosaur from the Wessex Formation (Lower Cretaceous) of the Isle of Wight, England". Cretaceous Research. 26: 686–698. doi:10.1016/j.cretres.2005.03.005.CS1 maint: multiple names: authors list (link)
- Witton 2013, pp. 158-159.
- Witton 2013, p. 160.
- Witton 2013, p. 159.
- Wellnhofer 1991, p. 55.
- Witton 2013, p. 162.
- Witton 2013, p. 163.
- Witton 2013, p. 179.
- Witton 2013, pp. 153-154.
- "Abstract: DIET OF ORNITHOCHEIROID PTEROSAURS INFERRED FROM STABLE CARBON ISOTOPE ANALYSIS OF TOOTH ENAMEL (GSA Annual Meeting in Seattle, Washington, USA - 2017)". gsa.confex.com.
- Adele H. Pentland; Stephen F. Poropat, 2018, "Reappraisal of Mythunga camara Molnar & Thulborn, 2007 (Pterosauria, Pterodactyloidea, Anhangueria) from the upper Albian Toolebuc Formation of Queensland, Australia". Cretaceous Research doi: 10.1016/j.cretres.2018.09.011
- Molnar, Ralph E.; Thulborn, R.A. (2008). "An incomplete pterosaur skull from the Cretaceous of north-central Queensland, Australia". Arquivos do Museu Nacional, Rio de Janeiro. 65 (4): 461–470.
- Rodrigo V. Pêgas, Borja Holgado & Maria Eduarda C. Leal (2019) On Targaryendraco wiedenrothi gen. nov. (Pterodactyloidea, Pteranodontoidea, Lanceodontia) and recognition of a new cosmopolitan lineage of Cretaceous toothed pterodactyloids, Historical Biology, doi:10.1080/08912963.2019.1690482
- Andres, B.; Myers, T. S. (2013). "Lone Star Pterosaurs". Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 103 (3–4): 1. doi:10.1017/S1755691013000303.
- Peter Wellnhofer, 1987, "New crested pterosaurs from the Lower Cretaceous of Brazil", Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 27: 175–186; Muenchen
- Bennett, S.C. (1994). "Taxonomy and systematics of the Late Cretaceous pterosaur Pteranodon (Pterosauria, Pterodactyloidea)" (PDF). Occasional Papers of the Natural History Museum, University of Kansas. 169: 1–70.
- Witton 2013, pp. 161-162.
- Witton, M.P.; Habib, M.B. (2010). "On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness". PLOS ONE. 5 (11): e13982. Bibcode:2010PLoSO...513982W. doi:10.1371/journal.pone.0013982. PMC 2981443. PMID 21085624.
- Habib, M. (2011). "Dinosaur Revolution: Anhanguera." H2VP: Paleobiomechanics. Weblog entry, 20-SEP-2011. Accessed 28-SEP-2011: http://h2vp.blogspot.com/2011/09/dinosaur-revolution-anhanguera.html
- Averianov A.O. (2012). "Ornithostoma sedgwicki – valid taxon of azhdarchoid pterosaurs". Proceedings of the Zoological Institute RAS. 316 (1): 40–49.
- Nopcsa, Baron Francis (1923). "Notes on the British dinosaurs, Part IV: Acanthopholis". Geological Magazine. 60 (5): 193–199. doi:10.1017/S0016756800085563.
- Seeley, H. G. (1879). "On the Dinosauria of the Cambridge Greensand". Quarterly Journal of the Geological Society. 35 (1–4): 591–635. doi:10.1144/GSL.JGS.1879.035.01-04.42.
- H.G. Seeley, 1876, "On Macrurosaurus semnus (Seeley), a long tailed animal with procoelous vertebrae from the Cambridge Upper Greensand, preserved in the Woodwardian Museum of the University of Cambridge", Quarterly Journal of the Geological Society of London 32: 440-444 doi:10.1144/GSL.JGS.1876.032.01-04.50
- Harrison, C.J.O.; Walker, C.A. (1973). "Wyleyia: a new bird humerus from the Lower Cretaceous of England" (PDF). Palaeontology. 16 (4): 721–728. Archived from the original (PDF) on 2012-02-06.
- Valentin Fischer; Nathalie Bardet; Myette Guiomar & Pascal Godefroit (2014). "High Diversity in Cretaceous Ichthyosaurs from Europe Prior to Their Extinction". PLoS ONE. 9 (1): e84709. doi:10.1371/journal.pone.0084709. PMC 3897400. PMID 24465427.
- Veldmeijer, A.J. (2006). "Toothed pterosaurs from the Santana Formation (Cretaceous; Aptian-Albian) of northeastern Brazil. A reappraisal on the basis of newly described material Archived 2012-03-17 at the Wayback Machine." Tekst. - Proefschrift Universiteit Utrecht.
- Fastnacht, M (2001). "First record of Coloborhynchus (Pterosauria) from the Santana Formation (Lower Cretaceous) of the Chapada do Araripe of Brazil". Paläontologische Zeitschrift. 75: 23–36. doi:10.1007/bf03022595.
- Dalla, F.V.; Ligabue, G. (1993). On the presence of a giant pterosaur in the Lower Cretaceous (Aptian) of Chapada do Araripe (northeastern Brazil) (PDF). Bollettino della Società Paleontologica Italiana. 32. pp. 131–136.CS1 maint: uses authors parameter (link)
- Ross A. Elgin & Eberhard Frey (2011). "A new ornithocheirid, Barbosania gracilirostris gen. et sp. nov. (Pterosauria, Pterodactyloidea) from the Santana Formation (Cretaceous) of NE Brazil". Swiss Journal of Palaeontology. 130 (2): 259–275. doi:10.1007/s13358-011-0017-4.
- Renan A. M. Bantim; Antônio A. F. Saraiva; Gustavo R. Oliveira; Juliana M. Sayão (2014). "A new toothed pterosaur (Pterodactyloidea: Anhangueridae) from the Early Cretaceous Romualdo Formation, NE Brazil". Zootaxa. 3869 (3): 201–223. doi:10.11646/zootaxa.3869.3.1. PMID 25283914.
- Veldmeijer, A.J., Meijer, H.J.M. & Signore, M. (2009). "Description of Pterosaurian (Pterodactyloidea: Anhangueridae, Brasileodactylus) remains from the Lower Cretaceous of Brazil". DEINSEA. 13: 9–40.CS1 maint: uses authors parameter (link)
- Leonardi, G. & Borgomanero, G. (1985). "Cearadactylus atrox nov. gen., nov. sp.: novo Pterosauria (Pterodactyloidea) da Chapada do Araripe, Ceara, Brasil." Resumos dos communicaçoes VIII Congresso bras. de Paleontologia e Stratigrafia, 27: 75–80.
- P. H. Buisonjé. 1980. Santanadactylus brasilensis nov. gen., nov. sp., a long-necked, large pterosaurier from the Aptian of Brasil. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, B 83:145-172
- Martill, David M. (2011). "A new pterodactyloid pterosaur from the Santana Formation (Cretaceous) of Brazil". Cretaceous Research. 32 (2): 236–243. doi:10.1016/j.cretres.2010.12.008.
- Pinheiro, Felipe L.; Fortier, Daniel C.; Schultz, Cesar L.; De Andrade, José Artur F.G.; Bantim, Renan A.M. (2011). "New information on Tupandactylus imperator, with comments on the relationships of Tapejaridae (Pterosauria)". Acta Palaeontologica Polonica. 56 (3): 567–580. doi:10.4202/app.2010.0057.
- Pêgas, R. V.; Costa, F. R.; Kellner, A. W. A. (2018). "New Information on the osteology and a taxonomic revision of The genus Thalassodromeus (Pterodactyloidea, Tapejaridae, Thalassodrominae)". Journal of Vertebrate Paleontology. 38 (2): e1443273. doi:10.1080/02724634.2018.1443273.
- Witton, Mark P. (October 2009). "A new species of Tupuxuara (Thalassodromidae, Azhdarchoidea) from the Lower Cretaceous Santana Formation of Brazil, with a note on the nomenclature of Thalassodromidae". Cretaceous Research. 30 (5): 1293–1300. doi:10.1016/j.cretres.2009.07.006.
- Kellner, A.W.A.; Campos, D.A. (1996). "First Early Cretaceous theropod dinosaur from Brazil with comments on Spinosauridae". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 199 (2): 151–166. doi:10.1127/njgpa/199/1996/151.
- Martill, D.M.; Cruickshank, A.R.I.; Frey, E.; Small, P.G.; Clarke, M. (1996). "A new crested maniraptoran dinosaur from the Santana Formation (Lower Cretaceous) of Brazil". Journal of the Geological Society. 153 (1): 5–8. Bibcode:1996JGSoc.153....5M. doi:10.1144/gsjgs.153.1.0005.
- Ortega, F. J.; Z. B. Gasparini; A. D. Buscalioni; J. O. Calvo (2000). "A new species of Araripesuchus (Crocodylomorpha, Mesoeucrocodylia) from the Lower Cretaceous of Patagonia (Argentina)". Journal of Vertebrate Paleontology. 20 (1): 57–76. doi:10.1671/0272-4634(2000)020[0057:ANSOAC]2.0.CO;2.
- Zug, George R. "Turtle: Origin and evolution". Encyclopædia Britannica. Retrieved 18 September 2015.
- Hirayama, Ren (1998-04-16). "Oldest known sea turtle". Nature. 392 (6677): 705–708. doi:10.1038/33669.
- Lane, Jennifer A.; Maisey, John G. (2012). "The Visceral Skeleton and Jaw Suspension In the Durophagous Hybodontid Shark Tribodus limae from the Lower Cretaceous of Brazil". Journal of Paleontology. 86 (5): 886–905. doi:10.1666/11-139.1. ISSN 0022-3360.
- Lee, Y.-N. (1994). "The Early Cretaceous Pterodactyloid Pterosaur Coloborhynchus wadleighi from North America". Palaeontology. 37 (4): 755–763.
- Paulina-Carabajal, A.; Lee, Y.N.; Jacobs, L.L. (2016). "Endocranial Morphology of the Primitive Nodosaurid Dinosaur Pawpawsaurus campbelli from the Early Cretaceous of North America". PLoS ONE. 11 (3): e0150845. doi:10.1371/journal.pone.0150845. PMC 4805287. PMID 27007950.
- Coombs, W. P. 1995. A nodosaurid ankylosaur (Dinosauria: Ornithischia) from the Lower Cretaceous of Texas. Journal of Vertebrate Paleontology 15(2):298-312. doi:10.1080/02724634.1995.10011231
- Weishampel, David B; et al. (2004). "Dinosaur distribution (Early Cretaceous, North America)." In: Weishampel, David B.; Dodson, Peter; and Osmólska, Halszka (eds.): The Dinosauria, 2nd, Berkeley: University of California Press. Pp. 553-556. ISBN 0-520-24209-2.
- William A.C. (1969). The Late Cretaceous Ammonites Scaphites leei Reeside and Scaphites hippocrepis (DeKay) in the Western Interior of the United States (PDF). United States Government Printing Office. pp. 3–7.
- Cappetta H. 1987. Chondrichthyes II. Mesozoic and Cenozoic Elasmobranchii. Schultze H.-P. (ed.), Handbook of Paleoichthyology, Volume 3B. Gustav Fischer Verlag, Stuttgart, 193 p.
Further reading
- Wellnhofer, Peter (1991). The Illustrated Encyclopedia of Pterosaurs: An illustrated natural history of the flying reptiles of the Mesozoic Era. Crescent Books. ISBN 0517037017.CS1 maint: ref=harv (link)
- Witton, Mark (2013). Pterosaurs: Natural History, Evolution, Anatomy. Princeton University Press. ISBN 978-0691150611.CS1 maint: ref=harv (link)
External links