Drzewica Formation
The Drzewica Formation is a geologic formation in Szydłowiec, Poland. It is late Pliensbachian age.[1][2][3] Vertebrate fossils have been uncovered from this formation, including dinosaur tracks.[1] The Drzewica Formation is part of the Depositional sequence IV-VII of the late lower Jurassic Polish Basin, with the IV showing the presence of local Alluvial deposits, with possible meandriform deposition origin, dominated in Jagodne and Szydłowiec, while delta system occurred through the zone of the modern Budki.[1] The sequence V shows a reduction of the erosion in the Zychorzyn borehole of the Drzewica Formation, showing changes on the extension of the marine facies, where upper deposits change from Alluvial to Deltaic-Seashore depositional settings.[1] VI-VII facies where recovered on the Brody-Lubienia borehole, with a lower part exposed on the village of Smilów that shows a small fall of the Sea level.[1] The stathigraphic setting of the dinosaur tracks reported from the formation suggest a Seashore or Deltaic barrier. Body fossils reported include bivalves, palynology, fossil trunks, roots. Trunks of coniferous wood, especially Cheirolepidiaceae and Araucariaceae trees show the occurrence of vast coniferous forests around the tracksite.[4][5] The association of forests and dinosaur megafauna on the Pliensbachian suggests also a colder and specially damp ecosystem. As many studies of the formation share, Drzewica shows in part to be a gigantic shore barrel, setting at the time where the Polish basin sea was at its lowest point.[1] Other related units are Fjerritslev or Gassum Formation (Danish Basin), lower Bagå Formation (Bornholm), upper Neringa Formation (Lithuania). Abandoned informal units in Poland: upper Sawêcin beds, Wieluñ series or Bronów series.[6]
| Drzewica Formation Stratigraphic range: late Pliensbachian ~183 Ma [1] | |
|---|---|
 Quarry with Drzewica strata at the bottom | |
| Type | Geological formation |
| Unit of | Kamienna Group[1] |
| Underlies | Ciechocinek Formation, Borucice Formation[1] |
| Overlies | Zagaje, Lobez, Ostrowiec & Gielniów Formations |
| Thickness | Up to ~100 m (330 ft)[1] |
| Lithology | |
| Primary | Siliciclastic sandstones |
| Other | Several types of heteroliths and mudstones |
| Location | |
| Region | Szydłowiec, Southern Poland |
| Country | |
| Extent | Polish Liassic Basin[1] |
| Type section | |
| Named for | Drzewica, Poland |
| Named by | Karaszewski (as an informal unit)[2] |
Stratygraphic Profile
| Unit | Lithology | Thickness (metres) | Fossils |
|---|---|---|---|
|
Non Drzewica Formation (Ciechocinek Formation, Lower Toarcian)[7] |
Clay, Claystone |
2108,5–2111,0 m on the General Lithological Profile |
Fish Remains, Bivalves, Ostracodans, Crustaceans and Plants |
|
N1[7] |
Gray Sandstones |
2111.0–2115.0 m |
Bivalve Trace Fossils, Plant Remains |
|
N2[7] |
2115.0-2118.0 m |
Dinoflagellates, Bivalves, Bivalve Trace Fossils, Limuloids, Ammonites, Leech Cocoons, Fish Teeth, Rhynchocephalia footprints, Testudinata footpints, Crocodrylomorpha footprints, Dinosaur Footprints, Plant Remains & Pollen | |
|
N3[7] |
Sandstones, in the upper section salted, less clayy downwards |
2118.0–2134.5 m |
Bivalves, Bivalve Trace Fossils, Limuloids, Ammonites, Leech Cocoons, Fish Teeth, Rhynchocephalia footprints, Testudinata footpints, Synapsid Footprints, Crocodrylomorpha footprints, Dinosaur Footprints, Plant Remains, Pollen, Plant & Fungus Spores. |
|
N4[7] |
Mudstone with sandstone inserts |
2134.5–2149.0 m |
Bivalves, Bivalve Trace Fossils, Limuloids, Ammonites, Plant Remains, Pollen, Plant & Fungus Spores. |
|
N5[7] |
Sandstone and infiltrated sandstone |
2149.0–2167.0 m |
Bivalve Trace Fossils & Pollen |
|
N6[7] |
Claystone |
2,167.0–2168.5 m |
Dinoflagellates, Bivalve Trace Fossils & Pollen |
|
N7[7] |
Sandstones |
2168.5–2175.0 m |
Dinoflagellates, Bivalve Trace Fossils & Pollen |
|
Core[7] |
|
2,175.0–2181.0+2.00 m |
Dinoflagellates, Bivalve Trace Fossils, Fish Teeth, Fungus, Plant Remains & Pollen |
|
SUB-Core (No Core)[7] |
Sandstones |
According to geophysical measurements up to a depth of 2178.0 m |
Dinoflagellates & Pollen |
Depositional systems
.jpg)
There are several Types os Seashore Stratification Seen On the Drzewica Formation. Starting with the shore prograding cycle, where the nearshore deposits show detailed Hummocky cross-stratification, at the time are covered by thin beach-welded facies showing irregular “massive” bedding and numerous Plant roots penetrating down from the undulated bounding surface.[1] Superimposed are fine grained, very well sorted Sandstones with giant tabular Cross-bedding set interpreted as beach (Backshore depositional subsystem) Eolian Dune with complete Plant remains buried in the wind-transported Sand (Mostly, Herbaceous-Built Plants). The Direction of the Wind sedimentation is clearly visible on this deposits, where the stratification of the sand is well oriented. Those deposits are abundant on the Antecki quarry.[1] The most related to the shores, probably barrier-Lagoon and Delta depositional systems are clearly visible on the Smilów Quarry, where the exposure shows different lithofacies and strata, including fine-grained, white and light-grey Sandstones with Hummocky cross-stratification, with a series of tabular and trough Cross-bedding sets, probably related to nearmarine depositional subsystem.[1] A more fine-medium-grained, grey to brownish sandstones also with flat-tabular-horizontal bedding with abundant drifted Plant Fossils and plant remains in situ is considered to be part of a clear foreshoreb–arrier–Eolian depositional subsystem, proabably related to a series of Dune fields, related to the Sea Barriers.[1] Kaolinized grey Mudstones with abundant Dispersed plant roots are related to more lagoonal depositional subsystems.[1] The slightly smaller and fine built, brownish and yellow Sandstones that show detailes Cross-bedding are related to a more alluvial deposition, proabablly from the influence of a local Channel.[1] On the Main Smilów Quarry the dominant shoreface depositional subsystem show detailed marks of clear Palaeocurrent directions, dispersed trough Cross-bedding inclinations that point to the West.[1] There has been measurements of the Wave-formed ripple that approximately reflects a shoreline orientation.[1] Thanks to the wave ripple crests orientation and the orientation of fossil tree logs it was indicated an inclination of the inclined tabular Foreset bed continuing across the whole outcrop.[1] These Foreset bed show a more pronounced inclination thanks to seaward-dipping Clinoforms, what reflects the original inclination of a barrier slope, with the sum of palaeocurrent pointing to south-west.[1]
Fungi
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Indeterminate |
|
Fungal spores of different sizes associated with wood |
Associated with a high rate of organic burial, the presence of Fungal Matter increased on the Uppermost layers of the Drzewica Formation, with a continue deposition between the T-OAE extincion, and several ups and downs on the Ciechocinek Formation, related with local climate and humidity changes. This is rather a reflection of the efficiency of terrestrial biodegradation.[8] Measured increasing of temperature favoured local fungal-mediated decomposition of plant litter, specifically of normally resistant wood.[8] |
 Xylophagous Fungi like Fomitopsis are probably the main origin for the Fungal Spores found on the Drzewica Formation | |
Paleofauna
Along the Early Jurassic, there was a mostly marginal-marine (mainly mesohaline), deltaic and continental sedimentation that prevailed in the area of Poland. The Marine of Polyhaline character conditions occurred mainly in the Pliensbachian, due to a flooding of the Polish Basin.[9] due to that, Pliensbachian sedimentation in Pomerania developed in a shelf basin that attained depths of about 100 m, below the storm wave base.[9] As the latest Pliensbachian began, the climate changes to a more cold environment, as prove the presence of ice sheets on northern latitude facies of the same age. That led to changes in salinity.[9] The Drzewica Formation was influenced by an ancient swampy-lagoonal sub-system, with Deltaic currents depositing fluvial sediments. The Main Boreholes show eustatic changes on the marine conditions, with a retreat of the sea level, leading to the appearance of vegetation and abundant pollen on the uppermost parts of the formation. The regressive phase of sea level is represented by the Komorowo Beds and Upper Slawecin Beds along with the Drzewica Formation. As the level of the sea recovered on the Toarcian, mostly of the setting turn out to be a bathial marine bottom.[10] There is a high report of footprints (+200), with different sizes, that range from small to large animals of various groups. The Smilow Quarry Tracksite is the main Borehole and setting for the Footprints.[4] Those are the only ones recovered from the Late Liassic of Europe.[4] The footprints are related to mostly lagoonar facies, unlike the Vegetation, that appears on Deltaic facies. Brackish deposits are relates to Brachiopods and other molluscs and lime animals. The presence of Late Pliensbachian common ammonoids and several vertebrate clades connects the marine fauna with similar environments of the same age across Europe. The Drzewica Fauna is also, one of the few from the Pliensbachian-Toarcian Anoxic event recorded worldwide, specially referred to the vertebrate fauna reported.[11]
Invertebrates
Annelida
Color key
|
Notes Uncertain or tentative taxa are in small text; |
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Dictyothylakos[12] |
|
Szydłowiec sandstones |
Leech cocoons |
Hirudinea cocoons, identified with palynological residues. The cocoons Dictyothylakos are common on flooded basin sediments, and implies not only the presence of parasitic leeches, but also the presence of large hosts nearby, that is proven on the case of Drzewica thanks to the large footprints. |
 Example of Leech Cocoon |
Bivalve Ichnofossils
Blackish bivalves were reported from the formation. They may be evidence of an ancient water system.[4]
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Traces |
These forms represent the resting traces of Bivalves. The Lockeia traces trend to accumulate on Brackish waters on the case of the Drzewica Formation. They can occur in large aggregates, where occasionally traces present on agglomerations show a clear orientation, probably parallel to the palaeocurrent direction, which is associated with locomotion of the burrowing animal. |
 Example of Lockeia Fossils | |
Bivalvia
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Pleuromya[14] |
|
Szydłowiec sandstones |
Cunchs |
A Clam, type member of the family Pleuromyidae inside Pholadida. A species heretofore known solely from the Scania and Bornholm region, derived there from the Doshult and Katslosa series. |
|
|
Szydłowiec sandstones |
Cunchs |
A Clam, type member of the family Cardiniidae inside Carditida. On the Pliensbachian strata there isn´t a clear consensus if the Polish basin reached stenohaline conditions, but the presence of this genus on the Parkoszowice 58 BN borehole along with foramiferans and marine phytoplankton tends to support more polyhaline-mesohaline conditions.[15] |
||
|
Pronoella[14] |
|
Szydłowiec sandstones |
Cunchs |
A Clam, member of the family Arcticidae inside Cardiida. |
|
|
Nuculana[14] |
|
Szydłowiec sandstones |
Cunchs |
A Pointed Nut Clam, type member of the family Nuculanidae inside Protobranchia. |
|
Ammonites
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Tragophylloceras loscombi |
Kaszewy 1 borehole |
MUZ.PGI.80.VI.149, single 1,7 cm Shell.[16] |
An Ammonite, member of the family Juraphyllitidae inside Ammonoidea. Latest Pliensbachian index fossil, Tragophylloceras is considered to be part of the Mediterranean-Submediterranean fossil region, but it has exceptions, with specimens coming from southern England and northwestern Germany.[16] In this case, the rocks that hosted the specimen also have poorly preserved dinoflagellate cysts, belonging to the genus Mendicodinium, that cannot expose its age beyond Late Pliensbachian.[16] This is fundamented by the presence of the fossil 20 m below the Lower Toarcian deposits. It looks also to be related to a large flooding surface, with the specimen being probably washed to nearshore or epicontinental waters.[16] |
 Example of
Tragophylloceras Fossils | |
Limuloidea
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Limulitella cf. liasokeuperinus |
Szydłowiec sandstones |
Head and partial upper body |
A Xiphosuran, member of the family Protostomia. Only representative Limuloid from the Lower Jurassic, Limulitella cf. liasokeuperinus was found associated with brackish and deltatic facies, related to nearshore deposits.[17] Its affinities with the genus Limulitella has not been proved. It has been suggested as Limulitella incertade sedis, on the basis that there is not enougth material to prove its affinities. The status of the species remain controversial, since the original fossil has been lost or in an unknown collection, more probably a private one.[18] |
.jpg) Limulus reconstruction, Limulitella was similar in morphology | |
Vertebrates
Chondrichthyes
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Indeterminate |
Szydłowiec sandstones |
Teeth |
A Guitarfish-like Chimaera, member of Chimaeriformes. Can be related to contemporaneous Squalorajoidei genera from Germany and England. Its affinities have not been tested. |
||
|
Szydłowiec sandstones |
Teeth |
A Shark, Type member of the family Hybodontidae inside Hybodontiformes. Contemporaneous Hybodus fossils are known from Rumania. Due to the abundance of Hybodus on Lower Jurassic Strata this has been a temporal assignation. Beyond that, the specimen hasn't been compared. |
 Restoration of Hybodus | |
|
Indeterminate |
Szydłowiec sandstones |
Teeth |
A Shark, member of the family Paleospinacidae inside Synechodontiformes. Due to be based on a few inclomplete Teeth it has dubious affinities. |
 Fossil of Synechodus | |
Actinopteri
Unidentified fin spines are known from this formation.[4]
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Pholidophorus? angustus |
Szydłowiec sandstones |
20 Specimens, complete and incomplete |
An Osteichthyes, type member of the family Pholidophoridae inside Teleostei. Possible Synonym of Furo angustus, was recovered from marine regresion deposits. |
 Pholidophorus | |
|
Proleptolepis sp. |
Szydłowiec sandstones |
Teeth |
An Osteichthyes, member of the family Leptolepididae inside Teleostei. |
||
|
Heterolepidotus latus |
Szydłowiec sandstones |
Teeth |
An Osteichthyes, member of the family Eugnathidae inside Ionoscopiformes. |
||
Sarcopterygii
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Cf. Coelacanthidae |
Szydłowiec sandstones |
Scales |
A Coelacanth, member of the family Coelacanthiformes inside Actinistia. The scales recovered are homologous to that found on the Triassic coelacanths Axelia & Wimania. |
||
|
Ptychoceratodus rectangulus |
Szydłowiec sandstones |
Teeth |
A Lungfish, type member of the family Ptychoceratodontidae inside Dipnoi. A Lungfish of relative small size, related with Freshwater Debris, found associated with Deltaic Deposits. |
||
|
Szydłowiec sandstones |
|
A Lungfish, member of the Neoceratodontidae inside Ceratodontiformes. A Lungfish, abundant on Europe since the Hettangian to the Bajocian on the Jurassic. The Species are known by a few scarce fragments, and are related with Triassic species found along Germany. |
 Illustration of Ceratodus by Heinrich Harder | |
Rhynchocephalia
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Rhynchosauroides[22] |
|
|
Tracks |
Lacertoid tracks done by possible Lepidosaurians, such as Squamatans (Relatives os the Jurassic basal squamatans, such as Scincomorpha members) but also to several Sphenodonts, such as Clevosaurus, Polysphenodon, and specially members of Gephyrosauridae, such as Gephyrosaurus and Deltadectes. Lizard-built floor dwelling animals of different sizes, but rather small. |
 Rhynchosauroides Footprints were similar to the feet of the genus Gephyrosaurus |
Testudinata
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Ancyropus[4] |
|
|
Tracks |
Basal Testudine Ichnotaxon, the relationships with Ancyropus are controversial, since the genus is known from the Early Liassic of North America. The tracks resemble Proganochelys, Palaeochersis, and probably other genera such as Kayentachelys or Australochelys. Small sized Turtles, with stimated less than 40 cm long, probably semiacuatic, related to the deltaic environments. Can be related to the Sinemurian and Toarcian Turtle fossils from Germany. |
|
Cynodontia
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Ameghinichnus[22] |
|
|
Tracks |
Represent tracks from Advanced Cynodonts, such as members of Tritylodontidae. The morphotypes resemble Oligokyphus, which is known from Pliensbachian Strata From England. Dinnebitodon, known from Pliensbachian strata of the Kayenta Formation, and larger ones to Bienotherium. That implicates the presence of various members of the late Cynodonts families, related to the modern mammals. Probably ground dwelling small animals that hunted insects or small vertebrates. |
 A genus similar to Oligokyphus is most probably the best candidate for the local Ameghinichnus footprints |
|
Brasilichnium[4] |
|
|
Tracks |
Represent tracks from Mammaliformes. Resemble creatures such as Sinoconodon, Morganucodonta such as Erythrotherium, Megazostrodon or Dinnetherium; more advanced mammaliformes, such as members of Haramiyida, including arborear forms such as Shenshou. As show by Middle-Late Jurassic Finds, this group was ecologically diverse. |
 A genus similar to Morganucodon is most probably the best candidate for the local Brasilichnium footprints |
Crocodrylomorpha
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Batrachopus[4] |
|
|
Tracks |
Terrestrial crocodrylomorph tracks, the only ones know from the Late liassic of Europe. It resemble Protosuchus-like small terrestrial crocodriles, and is probably related with the genera from the Kayenta Formation, such as Kayentasuchus or Eopneumatosuchus, or the Chinese genera Strigosuchus or Platyognathus. Seashore dwelling animals, with Carnivorous and Hervivorous representatives. |
_(20335774633).jpg) Terrestrial crocodriles such as Protosuchus, where most likely the Batrachopus trackmakers. |
Theropoda
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Cf. Stenonyx sp. |
|
Series of 30 mm Footprints |
Considered a very Small Dinosaur, probably a juvenile. The Foot resembles those of young Coelophysis from Ghost Ranch Quarries. Can also be tracks from primitive Theropods, such as Daemonosaurus, due to have ornithischian-pes like apperance. |
 Stenonyx footprints can come from a genus similar to Daemonosaurus | |
|
|
|
Assigned to Coelophysidae-alike dinosaurs. Small to medium slender primitive predatory dinosaurs, related with genera such as Liliensternus, Tachiraptor, Zupaysaurus, Procompsognathus or contemporaneous taxa such as the North American Segisaurus.[22][22][24] |
 Anchisauripus footprints belong to a genus similar to Procompsognathus | |
|
|
|
Assigned to Coelophysidae-alike dinosaurs. Small to medium slender primitive predatory dinosaurs, related with genera such as Liliensternus, Zupaysaurus, Procompsognathus or contemporaneous taxa such as the North American Segisaurus.[22][22][24] |
 Grallator footprints belong to a genus similar to Zupaysaurus | |
|
Kayentapus soltykovensis |
|
|
The tracks resemble Sinosaurus and are related with genera such as Notatesseraeraptor, Tachiraptor, Sarcosaurus and other basal Neotheropoda related to Tetanureae and Ceratosauriade. Small to medium-sized theropods. |
||
|
|
Eubrontes is related to the Genus Dilophosaurus, representing a basal Neotheropods. Include some large tracks, suggestings specimens up to 7 m, using Dilophosaurus as reference. Dilophosaurus is found or North America Pliensbachian strata, although the assignation of these tracks to the genus is controversial. Similar tracks have been described from Hettangian deposits from Poland and Sweden. There is a distinctive crouching theropod trace assigned to Eubrontes plexus, suggested a feather ingument on Dilophosauroids or relatives[13][27] |
 Model nicknamed "Dyzio", who represents a feathered reconstruction based on Drzewica and North American finds | ||
|
Megalosauripus sp. |
|
60 to 68 cm Footprints |
Largest Theropod Footprints from the Late Lower Jurassic. They have resemblance with the non-Tetanureae Sinosaurus, but match with Late Jurassic Orionides trackmakers. The trackmaker is expected to reach a length of 8.7-9.1 m long, with a weigth over 1.9-2.05 tonnes, making it among the largest Early Jurassic know theropod Trackmakers.[26] The footprints found on Drzewica seem even larger because of their large metatarsophalangeal area, which is often observed in the large theropod footprints from the post-Liassic strata.[26] |
 Megalosauripus footprints can belong to a large relative of Chuandongocoelurus | |
Sauropodomorpha
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
Kalosauropus? sp |
|
30 cm Footprints |
Sauropodomorph footprints that resemble Massopoda pes, specially the Massospondylidae genera. Lower Flora dwelling hervivores related to contemporaneous taxa, such as the North American genus Sarahsaurus or other genera, such as Seitaad, Lufengosaurus and Massospondylus. A massopod femur has been found recently on same age strata from Bornholm (Denmark)[28] |
 Local Kalosauropus can belong to a genus similar to Irisosaurus | |
|
|
40–50 cm Footprints |
Two different casts of footprints of primitive Sauropods, some with a foot model identical to the genus Barapasaurus, with an estimated body size of 6.8 m and others to Vulcanodon. The small size of the footprints can suggest juvenile individuals travelled along the deltaic facies of the formation. There are also Large Cetiosauriscus pes & Mamenchisauridae-like footprints, nearly identical to those found on the Chinese Lower Jurassic Yimen Formation and the Middle Jurassic Chuanjie Formation. They are the largest footprints reported from central Europe Early Jurassic. Can be related to the contemporaneous genus Tonganosaurus. Large hervivores, probably the major representatives of the local fauna. There are a series of tracks that also resemble the Ichnogenus Polyonyx, that is assigned to members of Turiasauria. |
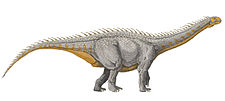 Local Parabrontopodus resemble the feet of the genus Barapasaurus | |
Ornithischia
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
|
Is considered synonymous with the ichnogenus Anomoepus. The tracks adscribed resemble basal Thyreorphora foots. The tracks related to the genus are vinculated with genera such as Scelidosaurus, whose fossils have been found on Pliensbachian strata of England. In this case, implicates a ~4 m long animal. |
Scelidosaurus matches with the bodyplan assigned to the Moyenisauropus trackmaker | |
|
|
5–25 cm Footprints |
A foot similar to Lesothosaurus, impliying 2.2 m long specimens. On the other casts there are tracks that resemble the feet of Stormbergia and various Genasauria of different sizes, between 90–130 cm. Fast-running small hervivorous ornithischians, probably gregarious. |
 Lesothosaurus matches with the bodyplan assigned to the Anomoepus trackmaker | |
Plants
The sediments of this formation in Smiłów Quarry are mainly fine-grained Sandstones with small over-minerals. Diagonal layers and wave strata appear in them, where there is also traces of root plants with numerous plant remains. Juzuz Karaszewski observed in the Drzewica Sandstone vertically preserved plants, and on this basis he derived a possible Dune-related origin of these sediments.[30] More recent studies suggest an environment of shallow seafront and beach, which of course does not exclude the presence of Aeolian zones there.[1] Lithofacies on the Smilów Quarry shows slightly inclined Beach foresets and abundant Plant roots in a series of barrier/foreshore–ackshore–eolian dune facies, that include even plants buried in whole Dune, while drifted plant fossils and tree logs occur on the main depositional surface. This inclined surface exposes most likely a long period of non deposition and washed plants, recovering at the same time the original tilt of the local beach.[1]
Palynology
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
Spores |
Affinities with Lycophyta inside Trachaeophyta. This Spores, rather abundant are correlated with humid settings, and come from mostly herbaceous-arbustive plants. |
 Restoration of the inland flora, including several species common at the time on Europe | |
|
Acanthotriletes[32] |
|
|
Spores |
Affinities with Lycophyta inside Trachaeophyta. Spores from low herbaceous flora, linked mostly to humid environments with abundant freshwater. | |
|
Hughesisporites[12] |
|
|
Spores |
Affinities with Lycophyta inside Trachaeophyta. | |
|
Maexisporites[33] |
|
|
Spores |
Affinities with Lycophyta inside Trachaeophyta. | |
|
Lycopodiumsporites[32] |
|
|
Spores |
Affinities with Lycopodiaceae inside Lycopodiopsida. Resemble spores of the modern genus Lycopodium. If it belongs to a similar genus, represent low herbaceous flora spores. | |
|
Reticulatisporites[32] |
|
|
Spores |
Affinities with Lycopodiaceae inside Lycopodiopsida. Resembles the modern genera Lycopodiella and Huperzia, characteristic small to medium (20-90 cm) herbaceous Lycopites, found on environments with abundant water. | |
|
Triletes[12] |
|
|
Spores |
Affinities with the Selaginellaceae inside Lycopsida. This genus resemble the Spores of the modern Selaginella selaginoides, representing probably very low herbaceous flora, found on very humid low envrionments. | |
|
Foveosporites[32] |
|
|
Spores |
Affinities with the Selaginellaceae inside Lycopsida. Less abundant than othe Lycophite Spores, it´s abundance increases near Deltaic deposits. | |
|
Klukisporites[32] |
|
|
Spores |
Affinities with the Selaginellaceae inside Lycopsida. | |
|
Nathorstisporites[31] |
|
|
Spores |
Affinities with Pleuromeiales inside Lycophyta. The Plueromeiales where tall Lycophites (2 to 6 m) common on the Trassic. Probably come from a relict genus. | |
|
Echitriletes[33] |
|
|
Spores |
Affinities with Isoetales inside Lycopsida. Spores related with modern Isoetes, representing Small plants related with water bodies. | |
|
Minerisporites[12] |
|
|
Spores |
Affinities with Isoetaceae inside Lycopsida. Related to plant similar to Isoetes lacustris, present on flooded basins, and other ecosystems with relative abundant water supply. | |
|
Perinopollenites[32] |
|
|
Pollen |
Affinities with Gymnospermopsida. Spores related to the Paleozoic Glossopteridales, such as Gangamopteris. Plants with arboreal built. | |
|
Chasmatosporites[32] |
|
|
Spores |
Affinities with Gymnospermopsida. Spores related to the Paleozoic Glossopteridales, such as Gangamopteris. Plants with arboreal built. | |
|
Apiculatisporites[32] |
|
|
Spores |
Affinities with Pteridopsida inside Filicopsida. Fern Spores of uncertain conrete placement. | |
|
Pilosisporites[32] |
|
|
Spores |
Affinities with Pteridopsida inside Filicopsida. Fern Spores of uncertain conrete placement. | |
|
Baculatisporites[32] |
|
|
Spores |
Affinities with Pteridopsida inside Filicopsida. Fern Spores of uncertain conrete placement. | |
|
Cingulatisporites[32] |
|
|
Spores |
Affinities with Pteridopsida inside Filicopsida. Fern Spores of uncertain conrete placement. | |
|
Concavisporites[32] |
|
|
Spores |
Affinities with Pteridopsida inside Filicopsida. Fern Spores of uncertain conrete placement. | |
|
Camptotriletes[32] |
|
|
Spores |
Affinities with Marattiaceae inside Filicopsida. Spores from large ferns related with hot and humid climate. | |
|
|
Pollen |
Affinities with Cycadopsida inside Gymnospermae. Pollen coming from Cycas or related genera, arbustive to arboreal species. | ||
|
Psophosphaera[32] |
|
|
Pollen |
Affinities with Cycadopsida inside Gymnospermae. | |
|
Bennettistemon[32] |
|
|
Spores |
Affinities with Bennettitales inside Cycadopsida. | |
|
Pseudowalchia[32] |
|
|
Pollen |
Affinities with Voltziales inside Coniferales. A possible relict genus, altrought it´s affinities are rather controversial. | |
|
Classopollis[34] |
|
|
Pollen |
Affinities with Cheirolepidiaceae inside Coniferales. Pollen from arboreal plants, the most abundant from its type locally. | |
|
Applanopsipollenites[32] |
|
|
Pollen |
Affinities with Araucariaceae inside Coniferales | |
|
Cupressacites[32] |
|
|
Pollen |
Affinities with Cupressaceae inside Coniferales. | |
|
Inaperturopollenites[32] |
|
|
Pollen |
Affinities with Pinidae inside Coniferales. | |
|
Rotundipollenites[32] |
|
|
Pollen |
Affinities with Pinidae inside Coniferales. | |
|
Tricolpites[34] |
|
|
Spores |
Affinities with Angiospermae and probably Magnoliophyta. Possible Pollen from primigenial Flowering plants or relatives, such as Schmeissneria or Sanmiguelia. | |
|
Clavatipollenites[32] |
|
|
Pollen |
Affinities with Angiospermae and probably Magnoliophyta. Possible Pollen from primigenial Flowering plants or relatives, such as Schmeissneria or Sanmiguelia. | |
Megaflora
| Genus | Species | Location | Material | Notes | Images |
|---|---|---|---|---|---|
|
|
MUZ PIG 76.III.1, complete plant |
A problematical plant that has been reported only on the Formation.[35] A plant buried in whole in an eolian dune, Sandstone, barrier-lagoon depositional system ( possible backshore/backbarrier depositional subsystem).[35] With leaves of 5–25 cm long and about 2–5 mm wide, and an angle between them and the short main axis ranges within 20-30°, This taxon has a morphology that resembles Schizeacean ferns (Specially Schizaea pectinata), but also that of the Voltziales Aethophyllum stipulare from Anisian of France (A Herbaceous Conifer From the Triassic). Grass-Like Plant with a very probable dune origin of these sediments.[35] Alternatively, can be leaves similar to Podozamites (Podocarpaceae). If not herbaceous plants, it can be leaves from trees or shrubs, but all probably from an Aeolian-Dunar Environment.[35] |
||
See also
- List of fossiliferous stratigraphic units in Poland
- Bagå Formation
- Neringa Formation
- Pliensbachian formations
References
- Pieñkowski, G. (2004). The epicontinental Lower Jurassic of Poland. Polish Geological Institute Special Papers, 12, 1-154.
- KARASZEWSKI W., 1962 — The stratigraphy of the Lias in the Northern Mesozoic Zone surrounding the Święty Krzyż Mountains (Central Poland) [Eng. Sum.]. Pr. Inst. Geol., 30, cz. 3:333–416
- Złonkiewicz, Z., & Fijałkowska-Mader, A. (2018). Kamień w architekturze zespołu koœcielno-klasztornego na Karczówce w Kielcach (Góry Œwiętokrzyskie).
- Pienkowski, Grzegorz (2008). "Preliminary report about spectacular late Pliensbachian dinosaur tracksite from the Holy Cross Mountains, Poland. The 5th International Symposium of IGCP 506, Marine and non-marine Jurassic: global correlation and major geological events". 110111. Cite journal requires
|journal=(help) - Ziaja, J. (2006). Lower Jurassic spores and pollen grains from Odrowąż, Mesozoic margin of the Holy Cross Mountains, Poland. Acta Palaeobotanica, 46(1), 3-83.
- Gierliński, G. D., Ploch, I., Sabath, K., & Ziaja, J. (2006). Zagadkowa roślina z wczesnej jury Gór Świętokrzyskich. Przegląd Geologiczny, 54(2), 139-141.
- Marek, S., KasińsKi, J., Krassowska, A., Leszczyński, K., Niemczycka, T., Feldman-Olszewska, A., ... & Gajewska, I. (2018). Szczegółowy profil litologiczno-stratygraficzny.
- Pieńkowski, G., Hodbod, M., & Ullmann, C. V. (2016). Fungal decomposition of terrestrial organic matter accelerated Early Jurassic climate warming. Scientific reports, 6, 31930.
- Pieñkowski, G., & Waksmundzka, M. (2009). Palynofacies in Lower Jurassic epicontinental deposits of Poland: tool to interpret sedimentary environments. Episodes, 32(1), 21-32.
- Feldman-Olszewska, A. (1997). Depositional systems and cyclicity in the intracratonic Early Jurassic basin in Poland. Geological Quarterly, 41(4), 475-490.
- BRAŃSKI, P. (2012). The mineralogical record of the Early Toarcian stepwise climate changes and other environmental variations (Ciechocinek Formation, Polish Basin). Volumina Jurassica, 10(10), 1-24.
- Marcinkiewicz,T. (1971) The stratigraphy of the Rhaetian and Liasin Poland based on megaspore investigations [ Stratygrafia Retyku i Liasu w Polsce na Podstawie badan megasporowych. ] Prace Instytut Geologiczny,(Warsaw) Vol. 65 P. 1- 57
- Gerard Dariusz Gierlinski, Martin G. Lockley, Grzegorz Niedźwiedzki. "A distinctive crouching theropod trace from the Lower Jurassic of Poland". Geological Quarterly. 53(4): 23 281.CS1 maint: uses authors parameter (link)
- Kopik, J. (1960). O kilku morskich małżach z serii gielniowskiej liasu Gór Świętokrzyskich [About several sea clams from the series of Gielniowska lias of the Świętokrzyskie Mountains]. Geological Quarterly , 4 (1), 95-104.
- Kilhams, B., Kukla, P. A., Mazur, S., McKie, T., Mijnlieff, H. F., van Ojik, K., & Rosendaal, E. (2018). Mesozoic resource potential in the Southern Permian Basin area: the geological key to exploiting remaining hydrocarbons whilst unlocking geothermal potential. Geological Society, London, Special Publications, 469(1), 1-18.
- Pienkowski, Grzegorz (2014): The first Early Jurassic ammonite find in central Poland. Volumina Jurassica, 2014, XII (1): 99–104:https://www.researchgate.net/publication/297380409_The_first_Early_Jurassic_ammonite_find_in_central_Poland
- Braun, C.F., 1860, Die Thiere in den Pflanzenschiefern der Gegend von Bayreuth: 1771, Zoologica Fundamenta
- Błażejowski, B., Niedźwiedzki, G., Boukhalfa, K., & Soussi, M. (2017). Limulitella tejraensis, a new species of limulid (Chelicerata, Xiphosura) from the Middle Triassic of southern Tunisia (Saharan Platform). Journal of Paleontology, 91(5), 960-967.
- Kopik J. and Marcinkiewicz T. 1997. Jura dolna: Biostratygrafia. In: S. Marek and M. Pajchlowa (Eds), The epicontinental Permian and Mesozoic in Poland (in Polish with English summary). Prace Paƒstwowego Instytutu Geologicznego, 153: 196-205.
- Kopik, J. (1998). Lower and Middle Jurassic of the north-eastern margin of the Upper Silesian Coal Basin. Biuletyn Państwowego Instytutu Geologicznego, 378, 67-129.
- KOPIK, J. (1960). OF THE LIAS IN THE ŚWIĘTY KRZYŻ MOUNTAINS. Kwartalnik geologiczny, 4(1), 103.
- Niedźwiedzki, G., Remin, Z., Roszkowska, J., & Meissner, U. (2008). New finds of dinosaur tracks in the Liassic strata of the Holy Cross Mountains, Poland. Przegląd Geologiczny, 57, 252-262.
- Jesper Milàn, Finn Surlyk. 2014: An enigmatic, diminutive theropod footprint in the shallow marine Pliensbachian Hasle Formation, Bornholm, Denmark
- Gierlinski, G. D. (2007). New dinosaur tracks in the Triassic, Jurassic and Cretaceous of Poland. Jornadas Internacionales sobre Paleontoloia de Dinosaurios y su Entrono, Salas de los Infantes, Burgos, Spain, 13-16.
- G. Gierlinski and G. Niedzwiedzki. 2005. New saurischian dinosaur footprints from the Lower Jurassic of Poland. Geological Quarterly 49(1):99-104
- Grzegorz Niedźwiedzki, zbyszek Remink: Gigantic theropod dinosaur footprints from the Upper Pliensbachian of the Holy Cross Mountains, Poland [in Polish with English abstract].Article in Przeglad Geologiczny 56(9):823-825 · September 2008
- Li, D. Q., Xing, L. D., Lockley, M. G., Romilio, A., Yang, J. T., & Li, L. F. (2019). The first theropod tracks from the Middle Jurassic of Gansu, Northwest China: new and rare evidence of quadrupedal progression in theropod dinosaurs. Journal of Palaeogeography, 8(1), 10.
- Milàn, J., Cuny, G. (2019):Knogle fra mini sauropod fundet ved Hasle. Geomuseum Faxe Danmark. University of Lyon Frankrig. https://www.researchgate.net/publication/332291953_Knogle_fra_mini_sauropod_fundet_ved_Hasle
- Gierliński, G. D. (2009). A preliminary report on new dinosaur tracks in the Triassic, Jurassic and Cretaceous of Poland. Actas de las IV Jornadas Internacionales sobre Paleontología de Dinosaurios y su Entorno Salas de los In fan tes, Burgos, 75-90.
- KARASZEWSKI W. & KOPIK J. 1970 — Jura dolna. Inst. Geol. Pr.,56: 65–98
- Dadlez,R. (1964) Outline of the Lias stratigraphy in Western Poland and correlation with the Lias of Central Poland. [ Zarys Stratygrafii Liasu w Polsce Zachodniej i Jego Korelacja z Liasem Polski Srodkowej. ] Kwartalnik Geologiczny (Instytut Geologiczy) Vol. 8 # 1 P. 122- 144
- Rogalska,M. (1971) Division of the Liassic deposits in Poland (Except for the Carpathian area) based on microscope examinations. M©moires du Bureau des Recherches G©ologiques et MiniЁres (BRGM) Vol. 75 P. 201- 210
- Jurkiewiczowa,I. (1967) The Lias of the western part of the Mesozoic Zone Surrounding the Swietokrzyskie (Holly Cross) Mountains and its correlation with the Lias of the Cracow-Wielun Range. (In: Stratigraphical-Palaeontological Investigations in Poland Vol.2) Biuletyn - Instytutu Geologiczny Vol. 200 P. 5- 132
- Dyakowska,J. (1958) The angiospermoid pollen from the Liassic flora in Poland Veroffentlichungen des Geobotanischen Inst.Rubel i N Zurich,Verhandlu,4th Int.Meet,Quat.Botan. Vol. 34 P. 42- 43
- Gierliñski, G.D., Ploch, I., Sabath, K., Ziaja, J., 2006. An Early Jurassic problematical plant from the Holy Cross Mountains, central Poland. Prz. Geol. 54, 139–141. https://www.researchgate.net/publication/289572652_An_Early_Jurassic_problematical_plant_from_the_Holy_Cross_Mountains_central_Poland