Triassic–Jurassic extinction event
The Triassic–Jurassic (Tr-J) extinction event, sometimes called the end-Triassic extinction, marks the boundary between the Triassic and Jurassic periods, 201.3 million years ago,[1] and is one of the major extinction events of the Phanerozoic eon, profoundly affecting life on land and in the oceans. In the seas, a whole class (conodonts)[2] and 23–34% of marine genera disappeared.[3][4] On land, all archosauromorphs other than crocodylomorphs, pterosaurs, and dinosaurs went extinct; some of the groups which died out were previously abundant, such as aetosaurs, phytosaurs, and rauisuchids. Some remaining therapsids and many of the large temnospondyl amphibians had gone extinct prior to the Jurassic as well. However, there is still much uncertainty regarding a connection between the Tr-J boundary and terrestrial vertebrates, due to a paucity of terrestrial fossils from the Rhaetian period of the Triassic.[5]
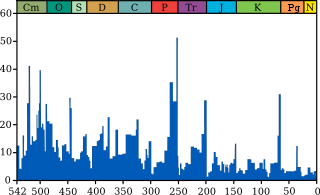
Statistical analysis of marine losses at this time suggests that the decrease in diversity was caused more by a decrease in speciation than by an increase in extinctions.[6] Nevertheless, a pronounced turnover in plant spores and a collapse of coral reef communities indicates that an ecological catastrophe did occur at the Triassic-Jurassic boundary. Older hypotheses on extinction have proposed that gradual climate or sea level change may be the culprit,[7] or perhaps one or more asteroid strikes.[8][9] However, the most well-supported and widely-held theory for the cause of the Tr-J extinction places the blame on the start of volcanic eruptions in the Central Atlantic Magmatic Province (CAMP). The CAMP is the geographically largest known large igneous province, and was responsible for outputting a high amount of Carbon Dioxide to induce profound global warming and ocean acidification.[10][11]
Effects
This event vacated terrestrial ecological niches, allowing the dinosaurs to assume the dominant roles in the Jurassic period. This event happened in less than 10,000 years and occurred just before Pangaea started to break apart. In the area of Tübingen (Germany), a Triassic–Jurassic bonebed can be found, which is characteristic for this boundary.[12]
The extinction event marks a floral turnover as well. About 60% of the diverse monosaccate and bisaccate pollen assemblages disappear at the Tr–J boundary, indicating a major extinction of plant genera. Early Jurassic pollen assemblages are dominated by Corollina, a new genus that took advantage of the empty niches left by the extinction.[13]
Marine invertebrates
Ammonites were impacted substantially by the Triassic-Jurassic extinction. Ceratitidans, the most prominent group of ammonites in the Triassic, went extinct at the end of the Rhaetian after having their diversity reduced significantly in the Norian. Other ammonite groups such as the Ammonitina, Lytoceratina, and Phylloceratina diversified from the Early Jurassic onward. Bivalves experienced high extinction rates at the early and middle Rhaetian. Plankton and gastropod diversity was barely affected at the T-J boundary, although there may have been local extinctions in radiolarians. Brachiopods slowly declined in diversity during the late Triassic before re-diversifying in the Early Jurassic. Conulariids seemingly completely died out at the end of the Triassic.[5] There is good evidence for a collapse in the reef community, as corals practically disappeared from the Tethys Ocean at the end of the Triassic and would not return to their previous abundance until the late Sinemurian. This reef collapse was likely driven by ocean acidification resulting from CO2 supplied to the atmosphere by the CAMP eruptions.[14][15][16]
Marine vertebrates

Fish did not suffer a mass extinction at the end of the Triassic. The late Triassic in general did experience a gradual drop in actinopterygiian diversity after an evolutionary explosion in the middle Triassic. Though this may have been due to falling sea levels or the Carnian pluvial event, it may instead be a result of sampling bias considering that middle Triassic fish have been more extensively studied than late Triassic fish.[17] Despite the apparent drop in diversity, neopterygiians (which include most modern bony fish) suffered less than more "primitive" actinopterygiians, indicating a biological turnover where modern groups of fish started to supplant earlier groups.[5] Conodonts, which were prominent index fossils throughout the Paleozoic and Triassic, finally went extinct at the T-J boundary following declining diversity.[5]
Like fish, marine reptiles experienced a substantial drop in diversity between the middle Triassic and the Jurassic. However, their extinction rate at the Triassic–Jurassic boundary was not elevated. The highest extinction rates experienced by Mesozoic marine reptiles actually occurred at the end of the Ladinian stage, which corresponds to the end of the middle Triassic. The only marine reptile families which went extinct at or slightly before the Triassic–Jurassic boundary were the placochelyids (the last family of placodonts), and giant ichthyosaurs such as shastasaurids and shonisaurids.[18] Nevertheless, some authors have argued that the end of the Triassic acted as a genetic "bottleneck" for ichthyosaurs, which never regained the level of anatomical diversity and disparity which they possessed during the Triassic.[19]
Terrestrial vertebrates

One of the earliest pieces of evidence for a late Triassic extinction was a major turnover in terrestrial tetrapods such as amphibians, reptiles, and synapsids. Edwin H. Colbert drew parallels between the system of extinction and adaptation between the Triassic–Jurassic and Cretaceous-Paleogene boundaries. He recognized how dinosaurs, lepidosaurs (lizards and their relatives), and crocodyliforms (crocodilians and their relatives) filled the niches of more ancient groups of amphibians and reptiles which were extinct by the start of the Jurassic.[7] Olson (1987) estimated that 42% of all terrestrial tetrapods went extinct at the end of the Triassic, based on his studies of faunal changes in the Newark Supergroup of eastern North America.[8] More modern studies have debated whether the turnover in Triassic tetrapods was abrupt at the end of the Triassic, or instead more gradual.[5]
During the Triassic, amphibians were mainly represented by large, crocodile-like members of the order Temnospondyli. Although the earliest lissamphibians (modern amphibians like frogs and salamanders) did appear during the Triassic, they would become more common in the Jurassic while the temnospondyls diminished in diversity past the Triassic–Jurassic boundary.[8] Although the decline of temnospondyls did send shockwaves through freshwater ecosystems, it was probably not as abrupt as some authors have suggested. Brachyopoids, for example, survived until the Cretaceous according to new discoveries in the 1990s. Several temnospondyl groups did go extinct near the end of the Triassic despite earlier abundance, but it is uncertain how close their extinctions were to the end of the Triassic. The last known metoposaurids ("Apachesaurus") were from the Redonda Formation, which may have been early Rhaetian or late Norian. Gerrothorax, the last known plagiosaurid, has been found in rocks which are probably (but not certainly) Rhaetian, while a capitosaur humerus was found in Rhaetian-age deposits in 2018. Therefore, plagiosaurids and capitosaurs were likely victims of an extinction at the very end of the Triassic, while most other temnospondyls were already extinct.[20]

Terrestrial reptile faunas were dominated by archosauromorphs during the Triassic, particularly phytosaurs and members of Pseudosuchia (the reptile lineage which leads to modern crocodilians). In the early Jurassic and onwards, dinosaurs and pterosaurs became the most common land reptiles, while small reptiles were mostly represented by lepidosauromorphs (such as lizards and tuatara relatives). Among pseudosuchians, only small crocodylomorphs did not go extinct by the end of the Triassic, with both dominant herbivorous subgroups (such as aetosaurs) and carnivorous ones (rauisuchids) having died out.[8] Phytosaurs, drepanosaurs, trilophosaurids, tanystropheids, and procolophonids, which were other common reptiles in the late Triassic, had also become extinct by the start of the Jurassic. However, pinpointing the extinction of these different land reptile groups is difficult, as the last period of the Triassic (the Rhaetian) and the first period of the Jurassic (the Hettangian) each have few records of large land animals. Some paleontologists have considered only phytosaurs and procolophonids to have gone extinct at the Triassic-Jurassic boundary, with other groups having gone extinct earlier.[5] However, it is likely that many other groups survived up until the boundary according to British fissure deposits from the Rhaetian. Aetosaurs, kuehneosaurids, drepanosaurs, "thecodontosaurids", "saltoposuchids" (like Terrestrisuchus), trilophosaurids, and various non-crocodylomorph pseudosuchians[21][22] are all examples of Rhaetian reptiles which may have gone extinct at the Triassic-Jurassic boundary.[23]
Possible causes
Gradual processes
Gradual climate change, sea-level fluctuations, or a pulse of oceanic acidification[24] during the late Triassic may have reached a tipping point. However, the effect of such processes on Triassic animal and plant groups is not well understood.
The extinctions at the end of the Triassic were initially attributed to gradually changing environments. Within his 1958 study recognizing biological turnover between the Triassic and Jurassic, Edwin H. Colbert's 1958 proposal was that this extinction was a result of geological processes decreasing the diversity of land biomes. He considered the Triassic period to be an era of the world experiencing a variety of environments, from towering highlands to arid deserts to tropical marshes. On the other hand, the Jurassic period was much more uniform both in climate and elevation due to excursions by shallow seas.[7]
Later studies noted a clear trend towards increased aridification towards the end of the Triassic. Although high-latitude areas like Greenland and Australia actually became wetter, most of the world experienced more drastic changes in climate as indicated by geological evidence. This evidence includes an increase in carbonate and evaporite deposits (which are most abundant in dry climates) and a decrease in coal deposits (which primarily form in humid environments such as coal forests).[5] In addition, the climate may have become much more seasonal, with long droughts interrupted by severe monsoons.[25]
Geological formations in Europe seem to indicate a drop in sea levels in the late Triassic, and then a rise in the early Jurassic. Although falling sea levels have sometimes been considered a culprit for marine extinctions, evidence is inconclusive since many sea level drops in geological history are not correlated with increased extinctions. However, there is still some evidence that marine life was affected by secondary processes related to falling sea levels, such as decreased oxygenation (caused by sluggish circulation), or increased acidification. These processes do not seem to have been worldwide, but they may explain local extinctions in European marine fauna.[5]
Extraterrestrial impact

Some have hypothesized that an impact from an asteroid or comet may have caused the Triassic–Jurassic extinction, similar to the extraterrestrial object which was the main factor in the Cretaceous–Paleogene extinction about 66 million years ago, as evidenced by the Chicxulub crater in Mexico. However, so far no impact crater of sufficient size has been dated to precisely coincide with the Triassic–Jurassic boundary.
Nevertheless, the late Triassic did experience several impacts, including the second-largest confirmed impact in the Mesozoic. The Manicouagan Reservoir in Quebec is one of the most visible large impact craters on Earth, and at 100 km (62 mi) in diameter it is tied with the Eocene Popigai crater in Siberia as the fourth-largest impact crater on Earth. Olsen et al. (1987) were the first scientists to link the Manicouagan crater to the Triassic–Jurassic extinction, citing its age which at the time was roughly considered to be late Triassic.[8] More precise radiometric dating by Hodych & Dunning (1992) has shown that the Manicouagan impact occurred about 214 million years ago, about 13 million years before the Triassic–Jurassic boundary. Therefore, it could not have been responsible for an extinction precisely at the Triassic–Jurassic boundary.[26] Nevertheless, the Manicougan impact did have a widespread effect on the planet; a 214-million-year-old ejecta blanket of shocked quartz has been found in rock layers as far away as England[27] and Japan. There is still a possibility that the Manicouagan impact was responsible for a small extinction midway through the late Triassic at the Carnian–Norian boundary,[26] although the disputed age of this boundary (and whether an extinction actually occurred in the first place) makes it difficult to correlate the impact with extinction.[27] Onoue et al. (2016) alternatively proposed that the Manicouagan impact was responsible for a marine extinction in the middle of the Norian which impacted radiolarians, sponges, conodonts, and Triassic ammonoids. Thus, the Manicouagan impact may have been partially responsible for the gradual decline in the latter two groups which culminated in their extinction at the Triassic–Jurassic boundary.[28] The boundary between the Adamanian and Revueltian land vertebrate faunal zones, which involved extinctions and faunal changes in tetrapods and plants, was possibly also caused by the Manicouagan impact, although discrepancies between magnetochronological and isotopic dating lead to some uncertainty.[29]
Other Triassic craters are closer to the Triassic–Jurassic boundary but also much smaller than the Manicouagan reservoir. The eroded Rochechouart crater in France has most recently been dated to 201±2 million years ago,[30] but at 25 km (16 mi) across (possibly up to 50 km (30 mi) across originally), it appears to be too small to have affected the ecosystem.[31] Other putative or confirmed Triassic craters include the 80 km (50 mi) wide Puchezh-Katunki crater in Eastern Russia (though it may be Jurassic in age), the 40 km (25 mi) wide Saint Martin crater in Manitoba, the 15 km (9 mi) wide Obolon' crater in Ukraine, and the 9 km (6 mi) wide Red Wing Creek structure in North Dakota. Spray et al. (1998) noted an interesting phenomenon, that being how the Manicoagan, Rochechoart, and Saint Martin craters all seem to be at the same latitude, and that the Obolon' and Red Wing craters form parallel arcs with the Rochechoart and Saint Martin craters, respectively. Spray and his colleagues hypothesized that the Triassic experienced a "multiple impact event", a large fragmented asteroid or comet which broke up and impacted the earth in several places at the same time.[9] Such an impact has been observed in the present day, when Comet Shoemaker-Levy 9 broke up and hit Jupiter in 1992. However, the "multiple impact event" hypothesis for Triassic impact craters has not been well-supported; Kent (1998) noted that the Manicouagan and Rochechoart craters were formed in eras of different magnetic polarity,[32] and radiometric dating of the individual craters has shown that the impacts occurred millions of years apart.[5]
Volcanic eruptions
Massive volcanic eruptions, specifically the flood basalts of the Central Atlantic Magmatic Province (CAMP), would release carbon dioxide or sulfur dioxide and aerosols, which would cause either intense global warming (from the former) or cooling (from the latter).[33][11] In addition to these climatic effects, oceanic uptake of volcanogenic carbon and sulphur dioxide would have led to a significant decrease of seawater pH known as ocean acidification, which is discussed as a relevant driver of marine extinction.[34][35][36] Evidence for ocean acidification as an extinction mechanism comes from the preferential extinction of marine organisms with thick aragonitic skeletons and little biotic control of biocalcification (e.g., corals, hypercalcifying sponges).[35] Global interruption of carbonate deposition at the Triassic-Jurassic boundary has been cited as additional evidence for catastrophic ocean acidification.[34] The record of CAMP degassing shows several distinct pulses of carbon dioxide immediately following each major pulse of magmatism, at least two of which amount to a doubling of atmospheric CO2.[37]
The isotopic composition of fossil soils of the Late Triassic and Early Jurassic has been tied to a large negative carbon isotope excursion (Whiteside et al. 2010). Carbon isotopes of lipids (n-alkanes) derived from leaf wax and lignin, and total organic carbon from two sections of lake sediments interbedded with the CAMP in eastern North America have shown carbon isotope excursions similar to those found in the mostly marine St. Audrie’s Bay section, Somerset, England; the correlation suggests that the end-Triassic extinction event began at the same time in marine and terrestrial environments, slightly before the oldest basalts in eastern North America but simultaneous with the eruption of the oldest flows in Morocco (Also suggested by Deenen et al., 2010), with both a critical CO
2 greenhouse and a marine biocalcification crisis.
Contemporaneous CAMP eruptions, mass extinction, and the carbon isotopic excursions are shown in the same places, making the case for a volcanic cause of a mass extinction. The catastrophic dissociation of gas hydrates (suggested as one possible cause of the largest mass extinction of all time, the so-called "Great Dying" at the end of the Permian Period) may have exacerbated greenhouse conditions.
Some scientists initially rejected the volcanic eruption theory, because the Newark Supergroup, a section of rock in eastern North America that records the Triassic–Jurassic boundary, contains no ash-fall horizons and its oldest basalt flows were estimated to lie around 10 m above the transition zone.[38] However, updated dating protocol and wider sampling has confirmed that the CAMP eruptions started in Nova Scotia and Morocco only a few thousand years before the extinction, and continued in several more pulses for the next 600,000 years.[11]
References
- Some sources (Whiteside et al 2010) give a date 181.8268 Ma.
- The extinction of conodonts — in terms of discrete elements — at the Triassic–Jurassic boundary
- Graham Ryder; David E. Fastovsky; Stefan Gartner (1996). The Cretaceous-Tertiary Event and Other Catastrophes in Earth History. Geological Society of America. p. 19. ISBN 9780813723075.
- Sepkoski, J. John (1984). "A kinetic model of Phanerozoic taxonomic diversity. III. Post-Paleozoic families and mass extinctions". Paleobiology. 10 (2): 246–267. doi:10.1017/s0094837300008186. ISSN 0094-8373.
- Tanner LH, Lucas SG, Chapman MG (2004). "Assessing the record and causes of Late Triassic extinctions" (PDF). Earth-Science Reviews. 65 (1–2): 103–139. Bibcode:2004ESRv...65..103T. doi:10.1016/S0012-8252(03)00082-5. Archived from the original (PDF) on October 25, 2007. Retrieved 2007-10-22.
- Bambach, R.K.; Knoll, A.H.; Wang, S.C. (December 2004). "Origination, extinction, and mass depletions of marine diversity". Paleobiology. 30 (4): 522–542. doi:10.1666/0094-8373(2004)030<0522:OEAMDO>2.0.CO;2. ISSN 0094-8373.
- Colbert, Edwin H. (1958-09-15). "Tetrapod Extinctions at the End of the Triassic Period" (PDF). Proceedings of the National Academy of Sciences. 44 (9): 973–977. Bibcode:1958PNAS...44..973C. doi:10.1073/pnas.44.9.973. ISSN 0027-8424. PMC 528676. PMID 16590299.
- Olsen, P. E.; Shubin, N. H.; Anders, M. H. (1987-08-28). "New early Jurassic tetrapod assemblages constrain Triassic–Jurassic tetrapod extinction event" (PDF). Science. 237 (4818): 1025–1029. Bibcode:1987Sci...237.1025O. doi:10.1126/science.3616622. ISSN 0036-8075. PMID 3616622.
- Spray, John G.; Kelley, Simon P.; Rowley, David B. (12 March 1998). "Evidence for a late Triassic multiple impact event on Earth" (PDF). Nature. 392 (6672): 171–173. Bibcode:1998Natur.392..171S. doi:10.1038/32397. ISSN 1476-4687.
- Hautmann, Michael (28 July 2004). "Effect of end-Triassic CO2 maximum on carbonate sedimentation and marine mass extinction". Facies. 50 (2): 257–261. doi:10.1007/s10347-004-0020-y.
- Blackburn, Terrence J.; Olsen, Paul E.; Bowring, Samuel A.; McLean, Noah M.; Kent, Dennis V; Puffer, John; McHone, Greg; Rasbury, Troy; Et-Touhami7, Mohammed (2013). "Zircon U-Pb Geochronology Links the End-Triassic Extinction with the Central Atlantic Magmatic Province" (PDF). Science. 340 (6135): 941–945. Bibcode:2013Sci...340..941B. CiteSeerX 10.1.1.1019.4042. doi:10.1126/science.1234204. PMID 23519213.
- Johannes Baier: Der Geologische Lehrpfad am Kirnberg (Keuper; SW-Deutschland) Archived 2011-10-02 at the Wayback Machine. - Jber. Mitt. oberrhein. geol. Ver, N. F. 93, 9–26, 2011.
- Fowell, S. J.; Cornet, B.; Olsen, P. E. (1994), "Geologically rapid Late Triassic extinctions: Palynological evidence from the Newark Supergroup", Geological Society of America Special Papers, Geological Society of America, pp. 197–206, doi:10.1130/spe288-p197, ISBN 978-0813722887
- Hönisch, Bärbel; Ridgwell, Andy; Schmidt, Daniela N.; Thomas, Ellen; Gibbs, Samantha J.; Sluijs, Appy; Zeebe, Richard; Kump, Lee; Martindale, Rowan C.; Greene, Sarah E.; Kiessling, Wolfgang (2012-03-02). "The Geological Record of Ocean Acidification". Science. 335 (6072): 1058–1063. Bibcode:2012Sci...335.1058H. doi:10.1126/science.1208277. ISSN 0036-8075. PMID 22383840.
- Greene, Sarah E.; Martindale, Rowan C.; Ritterbush, Kathleen A.; Bottjer, David J.; Corsetti, Frank A.; Berelson, William M. (2012-06-01). "Recognising ocean acidification in deep time: An evaluation of the evidence for acidification across the Triassic-Jurassic boundary". Earth-Science Reviews. 113 (1): 72–93. Bibcode:2012ESRv..113...72G. doi:10.1016/j.earscirev.2012.03.009. ISSN 0012-8252.
- Greene, Sarah E.; Bottjer, David J.; Corsetti, Frank A.; Berelson, William M.; Zonneveld, John-Paul (2012-11-01). "A subseafloor carbonate factory across the Triassic-Jurassic transition". Geology. 40 (11): 1043–1046. Bibcode:2012Geo....40.1043G. doi:10.1130/G33205.1. ISSN 0091-7613.
- Romano, Carlo; Koot, Martha B.; Kogan, Ilja; Brayard, Arnaud; Minikh, Alla V.; Brinkmann, Winand; Bucher, Hugo; Kriwet, Jürgen (27 November 2014). "Permian–Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolution". Biological Reviews of the Cambridge Philosophical Society. 91 (1): 106–147. doi:10.1111/brv.12161. ISSN 1469-185X. PMID 25431138.
- Bardet, Nathalie (1994-07-01). "Extinction events among Mesozoic marine reptiles". Historical Biology. 7 (4): 313–324. doi:10.1080/10292389409380462. ISSN 0891-2963.
- Thorne, Philippa M.; Ruta, Marcello; Benton, Michael J. (2011-05-17). "Resetting the evolution of marine reptiles at the Triassic–Jurassic boundary". Proceedings of the National Academy of Sciences. 108 (20): 8339–8344. Bibcode:2011PNAS..108.8339T. doi:10.1073/pnas.1018959108. ISSN 0027-8424. PMC 3100925. PMID 21536898.
- Konietzko-Meier, Dorota; Werner, Jennifer D.; Wintrich, Tanja; Martin Sander, P. (2018-10-31). "A large temnospondyl humerus from the Rhaetian (Late Triassic) of Bonenburg (Westphalia, Germany) and its implications for temnospondyl extinction". Journal of Iberian Geology. 45 (2): 287–300. doi:10.1007/s41513-018-0092-0. ISSN 1886-7995.
- Patrick, Erin L.; Whiteside, David I.; Benton, Michael J. (2019). "A new crurotarsan archosaur from the Late Triassic of South Wales" (PDF). Journal of Vertebrate Paleontology. Online edition (3): e1645147. doi:10.1080/02724634.2019.1645147. Archived from the original (PDF) on 30 August 2019.
- Tolchard, Frederick; Nesbitt, Sterling J.; Desojo, Julia B.; Viglietti, Pia; Butler, Richard J.; Choiniere, Jonah N. (2019-12-01). "'Rauisuchian' material from the lower Elliot Formation of South Africa and Lesotho: Implications for Late Triassic biogeography and biostratigraphy". Journal of African Earth Sciences. 160: 103610. Bibcode:2019JAfES.16003610T. doi:10.1016/j.jafrearsci.2019.103610. ISSN 1464-343X.
- Whiteside, D. I.; Marshall, J. E. A. (2008-01-01). "The age, fauna and palaeoenvironment of the Late Triassic fissure deposits of Tytherington, South Gloucestershire, UK". Geological Magazine. 145 (1): 105–147. Bibcode:2008GeoM..145..105W. doi:10.1017/S0016756807003925. ISSN 0016-7568.
- T.M. Quan, B. van de Schootbrugge, M.P. Field, "Nitrogen isotope and trace metal analyses from the Mingolsheim core (Germany): Evidence for redox variations across the Triassic–Jurassic boundary", Global Biogeochemical Cycles, 22 2008: "a series of events resulting in a long period of stratification, deep-water hypoxia, and denitrification in this region of the Tethys Ocean basin"; M. Hautmann, M.J. Benton, A. Toma, "Catastrophic ocean acidification at the Triassic–Jurassic boundary", Neues Jahrbuch für Geologie und Paläontologie 249.1, July 2008:119-127.
- T Parrish, Judith (1993). "Climate of the Supercontinent Pangea" (PDF). The Journal of Geology. 101 (2): 215–233. Bibcode:1993JG....101..215P. doi:10.1086/648217. JSTOR 30081148.
- Hodych, J. P.; Dunning, G. R. (1992-01-01). "Did the Manicouagan impact trigger end-of-Triassic mass extinction?". Geology. 20 (1): 51–54. Bibcode:1992Geo....20...51H. doi:10.1130/0091-7613(1992)020<0051:dtmite>2.3.co;2. ISSN 0091-7613.
- Racki, Grzegorz (2010). "The Alvarez impact theory of mass extinction; limits to its applicability and the "great expectations syndrome"" (PDF). Acta Palaeontologica Polonica. 57 (4): 681–702. doi:10.4202/app.2011.0058.
- Onoue, Tetsuji; Sato, Honami; Yamashita, Daisuke; Ikehara, Minoru; Yasukawa, Kazutaka; Fujinaga, Koichiro; Kato, Yasuhiro; Matsuoka, Atsushi (8 July 2016). "Bolide impact triggered the Late Triassic extinction event in equatorial Panthalassa". Scientific Reports. 6: 29609. Bibcode:2016NatSR...629609O. doi:10.1038/srep29609. ISSN 2045-2322. PMC 4937377. PMID 27387863.
- Kent, Dennis V.; Olsen, Paul E.; Lepre, Christopher; Rasmussen, Cornelia; Mundil, Roland; Gehrels, George E.; Giesler, Dominique; Irmis, Randall B.; Geissman, John W.; Parker, William G. (16 October 2019). "Magnetochronology of the entire Chinle Formation (Norian age) in a scientific drill core from Petrified Forest National Park (Arizona, USA) and implications for regional and global correlations in the Late Triassic". Geochemistry, Geophysics, Geosystems. 0 (ja): 4654–4664. Bibcode:2019GGG....20.4654K. doi:10.1029/2019GC008474. hdl:10150/636323. ISSN 1525-2027.
- Schmieder, M.; Buchner, E.; Schwarz, W. H.; Trieloff, M.; Lambert, P. (2010-10-05). "A Rhaetian 40Ar/39Ar age for the Rochechouart impact structure (France) and implications for the latest Triassic sedimentary record". Meteoritics & Planetary Science. 45 (8): 1225–1242. Bibcode:2010M&PS...45.1225S. doi:10.1111/j.1945-5100.2010.01070.x.
- Smith, Roff (2011-11-16). "Dark days of the Triassic: Lost world". Nature. 479 (7373): 287–289. Bibcode:2011Natur.479..287S. doi:10.1038/479287a. PMID 22094671.
- Kent, Dennis V. (10 September 1998). "Impacts on Earth in the Late Triassic" (PDF). Nature. 395 (6698): 126. Bibcode:1998Natur.395..126K. doi:10.1038/25874.
- Tanner, L. H.; J. F. Hubert; et al. (7 June 2001). "Stability of atmospheric CO
2 levels across the Triassic/Jurassic boundary". Nature. 411 (6838): 675–677. doi:10.1038/35079548. PMID 11395765. - Hautmann, Michael (28 July 2004). "Effect of end-Triassic CO2 maximum on carbonate sedimentation and marine mass extinction". Facies. 50 (2). doi:10.1007/s10347-004-0020-y.
- Hautmann, Michael; Benton, Michael J.; Tomašových, Adam (1 July 2008). "Catastrophic ocean acidification at the Triassic-Jurassic boundary". Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 249 (1): 119–127. doi:10.1127/0077-7749/2008/0249-0119.
- Greene, Sarah E.; Martindale, Rowan C.; Ritterbush, Kathleen A.; Bottjer, David J.; Corsetti, Frank A.; Berelson, William M. (June 2012). "Recognising ocean acidification in deep time: An evaluation of the evidence for acidification across the Triassic-Jurassic boundary". Earth-Science Reviews. 113 (1–2): 72–93. Bibcode:2012ESRv..113...72G. doi:10.1016/j.earscirev.2012.03.009.
- Schaller, Morgan F.; Wright, James D.; Kent, Dennis V. (2011-03-18). "Atmospheric Pco2 Perturbations Associated with the Central Atlantic Magmatic Province". Science. 331 (6023): 1404–1409. Bibcode:2011Sci...331.1404S. doi:10.1126/science.1199011. ISSN 0036-8075. PMID 21330490.
- Fowell, S.J.; Olsen, P.E. (May 1995). "Time calibration of Triassic/Jurassic microfloral turnover, eastern North America—Reply". Tectonophysics. 245 (1–2): 96–99. Bibcode:1995Tectp.245...96F. CiteSeerX 10.1.1.383.7663. doi:10.1016/0040-1951(94)00256-9. ISSN 0040-1951.
Literature
- Hodych, J. P.; G. R. Dunning (1992). "Did the Manicougan impact trigger end-of-Triassic mass extinction?". Geology. 20 (1): 51–54. Bibcode:1992Geo....20...51H. doi:10.1130/0091-7613(1992)020<0051:DTMITE>2.3.CO;2.
- McElwain, J. C.; D. J. Beerling; F. I. Woodward (27 August 1999). "Fossil Plants and Global Warming at the Triassic–Jurassic Boundary". Science. 285 (5432): 1386–1390. doi:10.1126/science.285.5432.1386. PMID 10464094.
- McHone, J.G. (2003), Volatile emissions of Central Atlantic Magmatic Province basalts: Mass assumptions and environmental consequences, in Hames, W.E. et al., eds., The Central Atlantic Magmatic Province: Insights from Fragments of Pangea. American Geophysical Union Monograph 136, p. 241-254.
- Tanner, L.H.; S.G. Lucas; M.G. Chapman (2004). "Assessing the record and causes of Late Triassic extinctions". Earth-Science Reviews. 65 (1–2): 103–139. Bibcode:2004ESRv...65..103T. doi:10.1016/S0012-8252(03)00082-5.
- Whiteside, Jessica H.; Paul E. Olsen; Timothy Eglinton; Michael E. Brookfield; Raymond N. Sambrotto (March 22, 2010). "Compound-specific carbon isotopes from Earth's largest flood basalt eruptions directly linked to the end-Triassic mass extinction". PNAS. 107 (15): 6721–5. Bibcode:2010PNAS..107.6721W. doi:10.1073/pnas.1001706107. PMC 2872409. PMID 20308590.
- Deenen, M.H.L.; M. Ruhl; N.R. Bonis; W. Krijgsman; W. Kuerscher; M. Reitsma; M.J. van Bergen (2010). "A new chronology for the end-Triassic mass extinction". Earth and Planetary Science Letters. 291 (1–4): 113–125. Bibcode:2010E&PSL.291..113D. doi:10.1016/j.epsl.2010.01.003.
- Hautmann, M. (2012). "Extinction: End-Triassic Mass Extinction". eLS. John Wiley & Sons, Ltd: Chichester. doi:10.1002/9780470015902.a0001655.pub3. ISBN 978-0470016176.
- Tetsuji Onoue; Honami Sato; Daisuke Yamashita; Minoru Ikehara; Kazutaka Yasukawa; Koichiro Fujinaga; Yasuhiro Kato; Atsushi Matsuoka (2016-07-08). "Bolide impact triggered the Late Triassic extinction event in equatorial Panthalassa". Scientific Reports. 6: 29609. Bibcode:2016NatSR...629609O. doi:10.1038/srep29609. PMC 4937377. PMID 27387863.
External links
- Theories on the Triassic–Jurassic Extinction
- The Triassic–Jurassic Mass Extinction
- 200 million year old mystery BBC News story, 12-Oct-2011
| Phenomena |  | ||||
|---|---|---|---|---|---|
| Models | |||||
| Causes | |||||
| Theories and concepts | |||||
| Extinction events |
| ||||
| Extinct species |
| ||||
| Organizations |
| ||||
| See also | |||||
| |||||