Equisetum
Equisetum (/ˌɛkwɪˈsiːtəm/; horsetail, snake grass, puzzlegrass) is the only living genus in Equisetaceae, a family of vascular plants that reproduce by spores rather than seeds.[2]
| Equisetum | |
|---|---|
 | |
| "Candocks" of Equisetum telmateia subsp. telmateia (great horsetail), showing whorls of branches and the tiny dark-tipped leaves | |
| Scientific classification | |
| Kingdom: | Plantae |
| Clade: | Tracheophytes |
| Class: | Polypodiopsida |
| Subclass: | Equisetidae |
| Order: | Equisetales |
| Family: | Equisetaceae |
| Genus: | Equisetum L. |
| Type species | |
| Equisetum arvense | |
| Species | |
|
See text | |
Equisetum is a "living fossil", the only living genus of the entire subclass Equisetidae, which for over 100 million years was much more diverse and dominated the understorey of late Paleozoic forests. Some equisetids were large trees reaching to 30 m (98 ft) tall.[3] The genus Calamites of the family Calamitaceae, for example, is abundant in coal deposits from the Carboniferous period. The pattern of spacing of nodes in horsetails, wherein those toward the apex of the shoot are increasingly close together, inspired John Napier to invent logarithms.[4]
A superficially similar but entirely unrelated flowering plant genus, mare's tail (Hippuris), is occasionally referred to as "horsetail", and adding to confusion, the name "mare's tail" is sometimes applied to Equisetum.[5]
Despite centuries of use in traditional medicine, there is no evidence that Equisetum has any medicinal properties.
Etymology
The name "horsetail", often used for the entire group, arose because the branched species somewhat resemble a horse's tail. Similarly, the scientific name Equisetum is derived from the Latin equus ("horse") + seta ("bristle").[6]
Other names include candock for branching species, and snake grass or scouring-rush for unbranched or sparsely branched species. The latter name refers to the rush-like appearance of the plants and to the fact that the stems are coated with abrasive silicates, making them useful for scouring (cleaning) metal items such as cooking pots or drinking mugs, particularly those made of tin. ('E. hyemale, rough horsetail, is still boiled and then dried in Japan to be used for the final polishing process on woodcraft to produce a smoother finish than any sandpaper.) In German, the corresponding name is Zinnkraut ("tin-herb"). In Spanish-speaking countries, these plants are known as cola de caballo, meaning "horsetail".
Description

Equisetum leaves are greatly reduced and usually non-photosynthetic. They contain a single, non-branching vascular trace, which is the defining feature of microphylls. However, it has recently been recognised that horsetail microphylls are probably not ancestral as in lycophytes (clubmosses and relatives), but rather derived adaptations, evolved by reduction of megaphylls.[7]
The leaves of horsetails are arranged in whorls fused into nodal sheaths. The stems are usually green and photosynthetic, and are distinctive in being hollow, jointed and ridged (with sometimes 3 but usually 6–40 ridges). There may or may not be whorls of branches at the nodes.
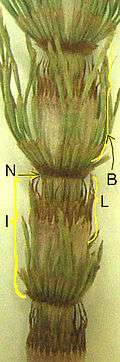
B = branch in whorl
I = internode
L = leaves
N = node


The small white protuberances are accumulated silicates on cells.
Spores
The spores are borne under sporangiophores in strobili, cone-like structures at the tips of some of the stems. In many species the cone-bearing shoots are unbranched, and in some (e.g. E. arvense, field horsetail) they are non-photosynthetic, produced early in spring. In some other species (e.g. E. palustre, marsh horsetail) they are very similar to sterile shoots, photosynthetic and with whorls of branches.
Horsetails are mostly homosporous, though in the field horsetail, smaller spores give rise to male prothalli. The spores have four elaters that act as moisture-sensitive springs, assisting spore dispersal through crawling and hopping motions after the sporangia have split open longitudinally.[8]
Equisetum cell walls
The crude cell extracts of all Equisetum species tested contain mixed-linkage glucan : Xyloglucan endotransglucosylase (MXE) activity.[9] This is a novel enzyme and is not known to occur in any other plants. In addition, the cell walls of all Equisetum species tested contain mixed-linkage glucan (MLG), a polysaccharide which, until recently, was thought to be confined to the Poales.[10][11] The evolutionary distance between Equisetum and the Poales suggests that each evolved MLG independently. The presence of MXE activity in Equisetum suggests that they have evolved MLG along with some mechanism of cell wall modification. Non-Equisetum land plants tested lack detectable MXE activity. An observed negative correlation between XET activity and cell age led to the suggestion that XET may be catalysing endotransglycosylation in controlled wall-loosening during cell expansion.[12] The lack of MXE in the Poales suggests that there it must play some other, currently unknown, role. Due to the correlation between MXE activity and cell age, MXE has been proposed to promote the cessation of cell expansion.
Taxonomy
Species
The living members of the genus Equisetum are divided into three distinct lineages, which are usually treated as subgenera. The name of the type subgenus, Equisetum, means "horse hair" in Latin, while the name of the other large subgenus, Hippochaete, means "horse hair" in Greek. Hybrids are common, but hybridization has only been recorded between members of the same subgenus.[13] While plants of subgenus Equisetum are usually referred to as horsetails, those of subgenus Hippochaete are often called scouring rushes, especially when unbranched.
Two Equisetum plants are sold commercially under the names Equisetum japonicum (barred horsetail) and Equisetum camtschatcense (Kamchatka horsetail). These are both types of E. hyemale var. hyemale, although they may also be listed as separate varieties of E. hyemale.[14]

Subgenus Paramochaete
- Equisetum bogotense Kunth – Andean horsetail; upland South America up to Costa Rica; includes E. rinihuense, sometimes treated as a separate species. Previously included in subg. Equisetum, but Christenhusz et al. (2019)[15] transfer this here, as E. bogotense appears to be sister to the remaining species in the genus.
Subgenus Equisetum
- Equisetum arvense L. – field horsetail, common horsetail or mare's tail; circumboreal down through temperate zones
- Equisetum diffusum L. – Himalayan horsetail; Himalayan India and China and adjacent nations above about 1500 feet (450 m)
- Equisetum fluviatile L. – water horsetail; circumboreal down through temperate zones
- Equisetum palustre L. – marsh horsetail; circumboreal down through temperate zones
- Equisetum pratense Ehrh. – meadow horsetail, shade horsetail, shady horsetail; circumboreal except for tundra down through cool temperate zones
- Equisetum sylvaticum L. – wood horsetail; circumboreal down through cool temperate zones, more restricted in east Asia
- Equisetum telmateia Ehrh. – great horsetail, northern giant horsetail; Europe to Asia Minor and north Africa, also west coast of North America. The North American subspecies Equisetum telmateia braunii (Milde) Hauke. may be treated as a separate species Equisetum braunii Milde[15]

Subgenus Hippochaete
- Equisetum giganteum L. – southern giant horsetail or giant horsetail; temperate to tropical South America and Central America north to southern Mexico
- Equisetum hyemale L. – rough horsetail, rough scouring rush; most of non-tropical northern hemisphere. The North American subspecies Equisetum hyemale affine (Engelm.) A.A.Eat. may be treated as a separate species Equisetum prealtum Raf.[15]
- Equisetum laevigatum A.Braun – smooth horsetail, smooth scouring rush; western 3/4 of North America down into northwestern Mexico; also sometimes known as Equisetum kansanum
- Equisetum myriochaetum Schltdl. & Cham. – Mexican giant horsetail; from central Mexico south to Peru
- Equisetum ramosissimum Desf. (including E. debile) – branched horsetail; Asia, Europe, Africa, southwest Pacific islands
- Equisetum scirpoides Michx. – dwarf horsetail, dwarf scouring rush; northern (cool temperate) zones worldwide
- Equisetum variegatum Schleich. ex Weber & Mohr – variegated horsetail, variegated scouring rush; northern (cool temperate) zones worldwide, except for northeasternmost Asia
Unplaced to subgenus
- †Equisetum dimorphum – Early Jurassic, Argentina
- †Equisetum thermale – Middle to Late Jurassic, Argentina
Named hybrids

Hybrids between species in subgenus Equisetum
- Equisetum × bowmanii C.N.Page (Equisetum sylvaticum × Equisetum telmateia)
- Equisetum × dycei C.N.Page (Equisetum fluviatile × Equisetum palustre)
- Equisetum × font-queri Rothm. (Equisetum palustre × Equisetum telmateia)
- Equisetum × litorale Kühlew ex Rupr. (Equisetum arvense × Equisetum fluviatile)
- Equisetum × mchaffieae C.N.Page (Equisetum fluviatile × Equisetum pratense)
- Equisetum × mildeanum Rothm. (Equisetum pratense × Equisetum sylvaticum)
- Equisetum × robertsii Dines (Equisetum arvense × Equisetum telmateia)
- Equisetum × rothmaleri C.N.Page (Equisetum arvense × Equisetum palustre)
- Equisetum × willmotii C.N.Page (Equisetum fluviatile × Equisetum telmateia)
Hybrids between species in subgenus Hippochaete
- Equisetum × ferrissii Clute (Equisetum hyemale × Equisetum laevigatum)
- Equisetum × moorei Newman (Equisetum hyemale × Equisetum ramosissimum)
- Equisetum × nelsonii (A.A.Eaton) Schaffn. (Equisetum laevigatum × Equisetum variegatum)
- Equisetum × schaffneri Milde (Equisetum giganteum × Equisetum myriochaetum)
- Equisetum × trachyodon (A.Braun) W.D.J.Koch (Equisetum hyemale × Equisetum variegatum)
Phylogeny
The phylogeny of extant species (excluding hybrids), according to Christenhusz et al. (2019), is shown in the following cladogram.[15]
| Equisetum |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Distribution and ecology
The genus Equisetum as a whole, while concentrated in the non-tropical northern hemisphere, is near-cosmopolitan, being absent only from Antarctica, though they are not known to be native to Australia, New Zealand nor the islands of the Pacific. They are most common in northern North America (Canada and the northernmost United States), where the genus is represented by nine species (E. arvense, E. fluviatile, E. hyemale, E. laevigatum, E. palustre, E. pratense, E. scirpoides, E. sylvaticum', and E. variegatum). Only four (E. bogotense, E. giganteum, E. myriochaetum, and E. ramosissimum) of the fifteen species are known to be native south of the Equator. They are perennial plants, herbaceous and dying back in winter as most temperate species, or evergreen as most tropical species and the temperate species E. hyemale (rough horsetail), E. ramosissimum (branched horsetail), E. scirpoides (dwarf horsetail) and E. variegatum (variegated horsetail). They typically grow 20 cm–1.5 m (8 in–5 ft) tall, though the "giant horsetails" are recorded to grow as high as 2.5 m (8 ft) (E. telmateia, northern giant horsetail), 5 m (16 ft) (E. giganteum, southern giant horsetail) or 8 m (26 ft) (E. myriochaetum, Mexican giant horsetail), and allegedly even more.[16]
One species, Equisetum fluviatile, is an emergent aquatic, rooted in water with shoots growing into the air. The stalks arise from rhizomes that are deep underground and difficult to dig out. Field horsetail (E. arvense) can be a nuisance weed, readily regrowing from the rhizome after being pulled out. It is unaffected by many herbicides designed to kill seed plants.[17] Since the leaves have a waxy coat, the plant is resistant to contact weedkillers like glyphosate.[18] However, as E. arvense prefers an acid soil, lime may be used to assist in eradication efforts to bring the soil pH to 7 or 8.[19] Members of the genus have been declared noxious weeds in Australia and in the US state of Oregon.[20][21]
All the Equisetum are classed as "unwanted organisms" in New Zealand and are listed on the National Pest Plant Accord.[22]
Consumption
People have regularly consumed horsetails. For example, the fertile stems bearing strobili of some species are cooked and eaten like asparagus (a dish called tsukushi[23]) in Japan.[24] Native Americans in the Pacific Northwest eat the young shoots of this plant raw.[25][26] The young plants are eaten cooked or raw, but considerable care must be taken.[27]
If eaten over a long enough period of time, some species of horsetail can be poisonous to grazing animals, including horses.[28] The toxicity appears to be due to thiaminase, which can cause thiamin (vitamin B1) deficiency.[27][29][30][31]
Equisetum species may have been a common food for herbivorous dinosaurs. With studies showing silicate within hadrosaur teeth and that horsetails are nutritionally of high quality, it is assumed that horsetails were an important component of herbivorous dinosaur diets.[32]
Folk medicine and safety concerns
Extracts and other preparations of E. arvense have served as herbal remedies, with records dating over centuries.[27][29][33] In 2009, the European Food Safety Authority concluded there was no evidence for the supposed health effects of E. arvense, such as for invigoration, weight control, skincare, hair health or bone health.[34] As of 2018, there is insufficient scientific evidence for its effectiveness as a medicine to treat any human condition.[27][33][34]
E. arvense contains thiaminase, which metabolizes the B vitamin, thiamine, potentially causing thiamine deficiency and associated liver damage, if taken chronically.[27][29] Horsetail might produce a diuretic effect.[27][29] Further, its safety for oral consumption has not been sufficiently evaluated and it may be toxic, especially to children and pregnant women.[27]
See also
References
- Channing, Alan; Zamuner, Alba; Edwards, Dianne; Guido, Diego (April 2011). "Equisetum thermale sp. nov. (Equisetales) from the Jurassic San Agustín hot spring deposit, Patagonia: anatomy, paleoecology, and inferred paleoecophysiology". American Journal of Botany. 98 (4): 680–97. doi:10.3732/ajb.1000211. PMID 21613167.
- Sunset Western Garden Book, 1995:606–607
- "An Introduction to the Genus Equisetum and the Class Sphenopsida as a whole". Florida International University. Archived from the original on 2009-07-14. Retrieved 2009-07-22.
- Sacks, Oliver (August 2011). "Field Trip: Hunting Horsetails". The New Yorker.
- Oxford English Dictionary.
- Daniel F. Austin (2004). Florida Ethnobotany (illustrated ed.). CRC Press. p. 283. ISBN 9780203491881.
- Rutishauser, R (November 1999). "Polymerous leaf whorls in vascular plants: Developmental morphology and fuzziness of organ identities". International Journal of Plant Sciences. 160 (S6): S81–S103. doi:10.1086/314221. PMID 10572024. S2CID 4658142.
- "Horsetail plant spores use 'legs' to walk and jump – BBC News". BBC News. Retrieved 2015-11-30.
- Fry, S. C.; Mohler, K. E.; Nesselrode, B. H. W. A.; Frankov, L. (2008). "Mixed-linkage -glucan:xyloglucan endotransglucosylase, a novel wall-remodelling enzyme from Equisetum (horsetails) and charophytic algae". The Plant Journal. 55 (2): 240–252. doi:10.1111/j.1365-313X.2008.03504.x. PMID 18397375.
- Fry, Stephen C.; Nesselrode, Bertram H. W. A.; Miller, Janice G.; Mewburn, Ben R. (2008). "Mixed-linkage (1→3,1→4)-β-d-glucan is a major hemicellulose of Equisetum (horsetail) cell walls". New Phytologist. 179 (1): 104–15. doi:10.1111/j.1469-8137.2008.02435.x. PMID 18393951.
- Sørensen, Iben; Pettolino, Filomena A.; Wilson, Sarah M.; Doblin, Monika S.; Johansen, Bo; Bacic, Antony; Willats, William G. T. (2008). "Mixed-linkage (1→3),(1→4)-β-d-glucan is not unique to the Poales and is an abundant component of Equisetum arvense cell walls". The Plant Journal. 54 (3): 510–21. doi:10.1111/j.1365-313X.2008.03453.x. PMID 18284587.
- Simmons, Thomas J.; Fry, Stephen C. (2017). "Bonds broken & formed during the mixed-linkage glucan: xyloglucan endotransglucosylase reaction catalysed by Equisetum hetero-trans-β-glucanase". Biochemistry. 474 (7): 1055–1070. doi:10.1042/BCJ20160935. PMC 5341106. PMID 28108640. Retrieved 2019-07-17.
- Pigott, Anthony (4 October 2001). "Summary of Equisetum Taxonomy". National Collection of Equisetum. Archived from the original on 21 October 2012. Retrieved 17 June 2013.
- "Weed Management Guide: Horsetails - 'Equisetum' specius" (PDF). environment.gov.au. 2013. Retrieved 2019-07-17.
- Christenhusz, Maarten J M; Bangiolo, Lois; Chase, Mark W; Fay, Michael F; Husby, Chad; Witkus, Marika; Viruel, Juan (April 2019). "Phylogenetics, classification and typification of extant horsetails (Equisetum, Equisetaceae)". Botanical Journal of the Linnean Society. 189 (4): 311–352. doi:10.1093/botlinnean/boz002.
- Husby, Chad E. (2003): How large are the giant horsetails? Version of 2003-03-19. Retrieved 2008-11-20. Archived April 4, 2004, at the Wayback Machine
- Altland, James (2003). "Horsetail - 'Equisetum arvense'". oregonstate.edu. Retrieved 2019-07-17.
- "Control Horse or Mare's Tail - Equisetum Arvense". Controlling Horsetail with Contact Herbicides. allotment-garden.org. 2016. Retrieved 2019-07-17.
- Kress, Henriette, Getting rid of horsetail, Henriette's Herbal Homepage, April 7th, 2005. Retrieved May 19, 2010.
- William Thomas Parsons; Eric George Cuthbertson (2001). Noxious weeds of Australia. CSIRO Publishing. p. 14. ISBN 978-0-643-06514-7.
- "Equisetum telmateia Ehrh. giant horsetail". USDA. Retrieved 2010-05-18.
- "National Pest Plant Accord" (PDF). rnzih.org.nz. 2001. Retrieved 2019-07-17.
- Michael Ashkenazi, Jeanne Jacob. 2003. Food culture in Japan. Greenwood Publishing Group. 232 p.
- Plants For A Future Database.
- Erna Gunther. 1973. Ethnobotany of western Washington: The knowledge and use of indigenous plants by Native Americans.
- Robin Harford Is field-horsetail edible
- "Horsetail". Drugs.com. 11 June 2018. Retrieved 19 August 2018.
- Israelsen, Clark E.; McKendrick, Scott S. & Bagley, Clell V. (2006): Poisonous Plants and Equine. PDF fulltext Archived January 12, 2011, at the Wayback Machine
- "Horsetail". MedlinePlus, US National Library of Medicine, National Institutes of Health. 8 December 2017. Retrieved 14 November 2013.
- Henderson JA, Evans EV, McIntosh RA (June 1952). "The antithiamine action of Equisetum". Journal of the American Veterinary Medical Association. 120 (903): 375–8. PMID 14927511.
- Fabre, B; Geay, B.; Beaufils, P. (1993). "Thiaminase activity in Equisetum arvense and its extracts". Plant Med Phytother. 26: 190–7.
- Dragos, D; Gilca, M; Gaman, L; Vlad, A; Iosif, L; Stoian, I; Lupescu, O (2017). "Phytomedicine in Joint Disorders". Nutrients. 9 (1): 70. doi:10.3390/nu9010070. PMC 5295114. PMID 28275210.
- "Scientific opinion on the substantiation of health claims related to Equisetum arvense L. and invigoration of the body (ID 2437), maintenance of skin (ID 2438), maintenance of hair (ID 2438), maintenance of bone (ID 2439), and maintenance or achievement of a normal body weight (ID 2783) pursuant to Article 13 of Regulation (EC) No 1924/2006". EFSA Journal. European Food Safety Authority. 7 (10): 1289. 2009. doi:10.2903/j.efsa.2009.1289.
Further reading
- Radosław Janusz Walkowiak, Equisetum Taxonomy, International Equisetological Association
- Husby Ch. E., Walkowiak R. J., An Introduction to the Genus Equisetum ( Horsetail ) and the Class Equisetopsida ( Sphenopsida ) as a whole. IEA Paper 2012
- Weber, Reinhard (2005): Equisetites aequecaliginosus sp. nov., ein Riesenschachtelhalm aus der spättriassischen Formation Santa Clara, Sonora, Mexiko; Equisetites aequecaliginosus sp. nov., a tall horsetail from the Late Triassic Santa Clara Formation, Sonora, Mexico Revue de Paléobiologie 24(1): 331–364 (German, English abstract)
External links
- Equisetum at the Tree of Life Web Project
- National Collection of Equisetum
- International Equisetological Association
- . Encyclopædia Britannica (11th ed.). 1911.
| Wikimedia Commons has media related to Equisetaceae. |
| Wikispecies has information related to Equisetum |