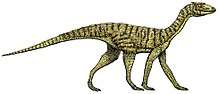
Tanystropheus
Tanystropheus (Greek τανυ~ “long” + στροφευς “hinged”), was a 6-meter-long (20 ft) reptile that dated from the Middle to Late Triassic epochs. It is recognizable by its extremely elongated neck, which measured 3 m (9.8 ft) long—longer than its body and tail combined.[1] The neck was composed of 12–13 extremely elongated vertebrae.[2] With its very long but relatively stiff neck, Tanystropheus has been often proposed and reconstructed as an aquatic or semi-aquatic reptile, a theory supported by the fact that the creature is most commonly found in semi-aquatic fossil sites wherein known terrestrial reptile remains are scarce. Fossils have been found in Europe. Complete skeletons of small individuals are common in the Besano Formation at Monte San Giorgio in Italy and Switzerland; other fossils have been found in the Middle East and China, dating from the Middle Triassic to the early part of the Late Triassic (Anisian, Ladinian, and Carnian stages).[3]
| Tanystropheus | |
|---|---|
 | |
| Restored Tanystropheus skeleton | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Family: | †Tanystropheidae |
| Genus: | †Tanystropheus Meyer, 1852 |
| Type species | |
| †Tanystropheus conspicuus nomen dubium Von Meyer, 1855 | |
| Other species | |
| |
| Synonyms | |
|
Genus synonymy
Species synonymy
| |
History
Tribelesodon, originally considered to be a pterosaur by Francesco Bassani in 1886, is now recognized as a junior synonym to Tanystropheus. Another junior synonym of Tanystropheus is Procerosaurus, whose type species, P. cruralis, was erected by von Huene in 1902. Several supposed Tanystropheus species described from the southwest United States in the late 19th century were later determined to belong to a new genus of theropod dinosaur, Coelophysis.[4]
The type species of Tanystropheus, T. conspicuus, is based on postcranial specimens from the Upper Muschelkalk of Germany and the Lower Keuper of Poland. These remains were previously known as Macroscelosaurus (a nomen oblitum) before being named as Tanystropheus in 1855. They were later regarded as undiagnostic, rendering T. conspicuus a nomen dubium.[4]
T. antiquus, which was named in the late 1900s from the Gogolin Formation of Poland, was based on vertebrae shorter than those of other Tanystropheus species. Its specimens were subsequently lost until they were rediscovered in the 21st century. T. antiquus is currently considered one of the few valid species of Tanystropheus.[4]
The best-known valid species of Tanystropheus is T. longobardicus, which comes from the Besano Formation (also known as the Grenzbitumenzone), an Anisian-Ladinian formation exposed at Monte San Giorgio on the Italy-Switzerland border.[5] The first T. longobardicus specimens were described in 1930s, and Monte San Giorgio continues to produce new specimens up to the present day. In 2005, Dr. Silvio Renesto described a T. longobardicus specimen from Switzerland which preserved the impressions of skin and other soft tissue. Five new specimens of T. longobardicus were described by Stefania Nosotti in 2007, allowing for a more comprehensive view of the anatomy of the species.[6]
Tanystropheus specimens from the Makhtesh Ramon in Israel were described as a new species, T. haasi, in 2001. However, this species may be dubious due to the difficulty of distinguishing its vertebrae from T. conspicuus or T. longobardicus. Another new species, T. biharicus, was described from Romania in 1975. It has also been considered possibly synonymous with T. longobardicus. A small but well-preserved skull and neck, specimen PIMUZ T 3901, was found in the Meride Limestone at Monte San Giorgio. It was given a new species, T. meridensis, in 1980. The skull was later referred to T. longobardicus, rendering T. meridensis a junior synonym of that species. Another species, T. fossai from the Norian-age Argillite di Riva di Solto in Italy, was given its own genus Sclerostropheus in 2019.[4]
Specimens similar to T. longobardicus have been found in the Ladinian or early Carnian Zhuganpo Formation in the Guizhou province of China, as described by Li (2007) and Rieppel (2010).[2] In 2014, fossils of an indeterminate species of Tanystropheus were discovered in the Wolfville Formation in the Bay of Fundy in Nova Scotia, Canada.[7] Indeterminate Tanystropheus remains are also known from the Jilh Formation of Saudi Arabia and various Anisian-Ladinian sites in Spain, Italy, and Switzerland.[4]
A 2019 revision of Tanystropheus found that only T. longobardicus and T. antiquus were valid species.[4] In 2020, large Tanystropheus specimens from Monte San Giorgio originally assigned to T. longobardicus were given a new species, T. hydroides.[8]
Anatomy

By far the most recognizable feature of Tanystropheus is its hyperelongate neck, equivalent to the combined length of the body and tail.[9] The neck was composed of 12–13 hyperelongate cervical vertebrae.[2] Cervical elongation reached its peak with cervical vertebra 9, which was ten times longer than it was tall. Weak development of cervical spines suggest that epaxial musculature was underdeveloped in Tanystropheus and that intrinsic back muscles (e.g., m. longus cervicis) were the driving force behind neck movement.[9] Subvertical placement of the pre- and postzygapophyses suggested limited lateral movement of the neck, whereas cervical ribs extending off these vertebrae would have formed a ventral brace that would transmit the forces from the weight of head and neck down to the pectoral girdle, providing passive support by limiting dorsoventral flexion.[10] The body contained 13 hourglass-shaped dorsal vertebrae, two sacral vertebrae and approximately twelve caudal vertebrae with weakly developed dorsal and haemal spines along with well-developed transverse processes.[9][11]
The pectoral and pelvic girdles are notably distinct. The forelimbs are smaller and more gracile than the hindlimbs, suggesting that the center of mass for Tanystropheus was closer towards the pelvic girdle. Attachment sites for the m. caudofemoralis muscle complex, coupled with soft-tissue preservation of relative muscle size, further support the proposition that Tanystropheus was a fairly bottom-heavy animal.[9]
Paleoecology

Diet
The diet of Tanystropheus has been controversial in the past, although most recent studies consider it a piscivorous (fish-eating) reptile. The teeth at the front of the narrow snout were long, conical, and interlocking, similar to those of nothosaurs and plesiosaurs. This was likely an adaptation for catching aquatic prey. Additionally, hooklets from cephalopod tentacles and what may be fish scales have been found near the belly regions of some specimens, further support for a piscivorous lifestyle.[6]
However, small specimens of the genus possess an additional, more unusual form of teeth. This form of teeth, which occurred in the rear part of the jaws behind the interlocking front teeth, were tricuspid (three-pronged), with a long and pointed central cusp and smaller cusps in front of and behind the central cusp. Wild (1974) considered these three-cusped teeth to be an adaptation for gripping insects. Cox (1985) noted that marine iguanas also had three-cusped teeth, and that Tanystropheus likely fed on marine algae like that species of lizard. Taylor (1989) rejected both of these hypotheses, as he considered the neck of Tanystropheus to be too inflexible for the animal to be successful at either diet.[6]

The most likely function of these teeth, as explained by Nosotti (2007), was that they assisted the piscivorous diet of the reptile by helping to grip slippery prey such as fish or squid. Several modern species of seals, such as the hooded seal and crabeater seal, also have multi-cusped teeth which assist their diet to a similar effect.[6] Similar teeth patterns have also been found in the pterosaur Eudimorphodon and the fellow tanystropheid Langobardisaurus, both of whom are considered piscivores. Large individuals of Tanystropheus, over 2 meters (6.6 ft) in length, lack these three-cusped teeth, instead possessing typical conical teeth at the back of the mouth. They also lack teeth on the pterygoid and palatine bones on the roof of the mouth, which possess teeth in smaller specimens. The two morphotypes were originally considered to represent juvenile and adult specimens of T. longobardicus. However, histology of the small specimens and restudy of the large specimens has shown that they each represent adult forms of two different species. The larger one-cusped morphotype was given a new species, T. hydroides, while the smaller tricuspid morphotype retained the name T. longobardicus.[8]
Soft tissue

The specimen described by Renesto in 2005 displayed an unusual "black material" around the rear part of the body, with smaller patches in the middle of the back and tail. Although most of the material could not have its structure determined, the portion just in front of the hip seemingly preserved scale impressions, indicating that the black material was the remnants of soft tissue. The scales seem to be semi-rectangular and do not overlap with each other, similar to the integument reported in a juvenile Macrocnemus described in 2002.[12] The portion of the material at the base of the tail is particularly thick and rich in phosphate. Many small spherical structures are also present in this portion, which upon further preparation were revealed to be composed of calcium carbonate. These chemicals suggest that the black material was formed as a product of the specimen's proteins decaying in a warm, stagnant, and acidic environment. As in Macrocnemus, the concentration of this material at the base of the tail suggests that the specimen had a quite noticeable amount of muscle behind its hips.[9]
Lifestyle
The lifestyle of Tanystropheus is controversial, with different studies favoring a terrestrial or aquatic lifestyle for the animal. In the 1980s, various studies determined that Tanystropheus lacked the musculature to raise its neck above the ground, and that it was likely completely aquatic, swimming by undulating its body and tail side-to-side like a snake or crocodile.
However, in 2005 Renesto found that Tanystropheus lacked many aquatic adaptations. Although the tail of Tanystropheus was flattened, it was compressed from top-to-bottom, so that it would have been useless for side-to-side movement. The long neck and short front limbs compared to the long hind limbs would have made four-limbed swimming inefficient and unstable if that was the preferred form of locomotion. Thrusting with only the hind limbs, as in swimming frogs, was also considered an inefficient form of locomotion for a large animal such as Tanystropheus, although a later study found support for this hypothesis. Renesto's study also found that the neck was lighter than previously suggested, and that the entire front half of the body was more lightly-built than the rear half, which would have possessed a large amount of muscle mass. In addition to containing powerful hind limb muscles, this unusually large muscle mass would have shifted the animal's weight to its rear, stabilizing the animal as it swung and maneuvered its massive neck.[9]
Renesto's analysis was the basis for later analyses of the genus. In 2015, paleoartist Mark Witton concluded that, although the neck made up half of the entire animal's length, it made up only 20% of the entire animal's mass due to having light and hollow vertebrae. In comparison, the heads and necks of pterosaurs of the family Azhdarchidae made up almost 50% of the animal's mass, yet they were clearly land based carnivores. The animal is also poorly equipped for aquatic life, with the only adaptation being a lengthened fifth toe, which suggests that it visited the water some of the time, though was not wholly dependent on it. This same research has also shown that Tanystropheus would have hunted prey like a heron.[13][14] This interpretation is supported by taphonomic evidence, which indicates that the preservation of Tanystropheus specimens is more similar to the terrestrial Macrocnemus than the aquatic Serpianosaurus where all three co-occur.[15]

Renesto and Franco Saller's 2018 follow-up to Renesto (2005)'s study offered more information on the reconstructed musculature of Tanystropheus. This study determined that the first few tail vertebrae of Tanystropheus would have housed powerful tendons and ligaments that would have made the body more stiff, keeping the belly off the ground and preventing the neck from pulling the body over. In addition, the hind limbs would have been quite flexible and powerful according to muscle correlations on the legs, pelvis, and tail vertebrae. This analysis shows that Tanystropheus was capable of a specific mode of movement while swimming. Namely, a Tanystropheus could extend its hind limbs forward and then simultaneously retract them, creating a powerful 'jump' forward. Further support for this hypothesis lies in the fact that a close relative of Tanystropheus, likely the small Tanytrachelos, is believed to have left an ichnogenus (trackway fossil) called Gwyneddichnium. Some Gwyneddichnium tracks seem to represent a succession of paired footprints that can be assigned to the hind limbs, but with no hand prints. These tracks were almost certainly created by the same form of movement which Renesto and Saller theorized was the preferred form of swimming in Tanystropheus.[17]
With this information, the most likely lifestyle for Tanystropheus was that the animal was a shallow-water predator that used its long neck to stealthily approach schools of fish or squid without disturbing its prey due to its large body size. Upon selecting a suitable prey item, it would have dashed forward by propelling itself along the seabed or through the water, with both hind limbs pushing off at the same time. However, this style of swimming is most common in amphibious creatures such as frogs, and likewise Tanystropheus would also have been capable of walking around on land. The idea that Tanystropheus evolved this form of swimming over much more efficient, yet specialized, styles is evidence that it did not live an exclusively aquatic life as in most other marine reptiles such as ichthyosaurs or plesiosaurs.[17]
A 2020 digital reconstruction of Tanystropheus skulls suggested that the animals, especially the Tanystropheus hydroides, lived primarily in the water because of the position of the nostrils.[18]
References
- Elbein, Asher (12 August 2020). "Making Sense of 'One of the Most Baffling Animals That Ever Lived' - Important mysteries have been solved about a reptile with a giraffe-like neck that hunted prey 242 million years ago". The New York Times. Retrieved 14 August 2020.
- Rieppel, Olivier; Jiang, Da-Yong; Fraser, Nicholas C.; Hao, Wei-Cheng; Motani, Ryosuke; Sun, Yuan-Lin; Sun, Zuo-Yu (2010). "Tanystropheus cf. T. Longobardicus from the early Late Triassic of Guizhou Province, southwestern China". Journal of Vertebrate Paleontology. 30 (4): 1082–1089. doi:10.1080/02724634.2010.483548. JSTOR 40864387.
- Dal Sasso, C. and Brillante, G. (2005). Dinosaurs of Italy. Indiana University Press. ISBN 0-253-34514-6, ISBN 978-0-253-34514-1.
- Spiekmann, Stephan N.F.; Scheyer, Torsten M. (December 2019). "A taxonomic revision of the genus Tanystropheus (Archosauromorpha, Tanystropheidae)". Palaeontologia Electronica. 22.3.80. doi:10.26879/1038.
- Tanystropheus. Vertebrate Palaeontology at Milano University. Retrieved 2007-02-19.
- Nossotti, Stefania (2007). "Tanystropheus longobardicus (Reptilia Protorosauria): Re-interpretations of the anatomy based on new specimens from the Middle Triassic of Besano (Lombardy, northern Italy)". Memoire della Societa Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano. 35 (3).
- Stratigraphic and Temporal Context and Faunal Diversity of Permian-Jurassic Continental Tetrapod Assemblages from the Fundy Rift Basin, Eastern Canada. Hans-Dieter Sues and Paul E. Olsen. Atlantic Geology 51; 2015; 139-205
- Spiekman, Stephan N. F.; Neenan, James M.; Fraser, Nicholas C.; Fernandez, Vincent; Rieppel, Olivier; Nosotti, Stefania; Scheyer, Torsten M. (2020-08-06). "Aquatic Habits and Niche Partitioning in the Extraordinarily Long-Necked Triassic Reptile Tanystropheus". Current Biology. 0 (0). doi:10.1016/j.cub.2020.07.025. ISSN 0960-9822.
- Renesto, S. (2005). "A new specimen of Tanystropheus (Reptilia, Protorosauria) from the Middle Triassic of Switzerland and the ecology of the genus". Rivista Italiana di Paleontologia e Stratigrafia. 111 (3): 377–394.
- Tschanz, K. 1988. Allometry and Heterochrony in the Growth of the Neck of Triassic Prolacertiform Reptiles. Paleontology. 31:997–1011.
- Wild, R. 1973. Tanystropheus longbardicus (Bassani) (Neue Egerbnisse). in Kuhn-Schnyder, E., Peyer, B. (eds) — Triasfauna der Tessiner Kalkalpen XXIII. Schweiz. Paleont. Abh. Vol. 95 Basel, Germany.
- Renesto, Silvio; Avanzini, Marco (2002). "Skin remains in a juvenile Macrocnemus bassanii Nopsca (Reptilia, Prolacertiformes) from the Middle Triassic of Northern Italy". Neues Jahrbuch für Geologie und Paläontologie. 224 (1): 31–48.
- Witton, Mark (13 November 2015). "Mark Witton.com Blog: The lifestyle of Tanystropheus, part 1: was that neck too heavy for use on land?". Retrieved 23 September 2018.
- Witton, Mark (11 December 2015). "Mark Witton.com Blog: The lifestyle of Tanystropheus, part 2: coastal fisher or first-day-on-the-job aquatic predator?". Retrieved 23 September 2018.
- Beardmore, S.R.; Furrer, H. (2017). "Land or water: using taphonomic models to determine the lifestyle of the Triassic protorosaur Tanystropheus (Diapsida, Archosauromorpha)". Palaeobiodiversity and Palaeoenvironments. 98 (2): 243–258. doi:10.1007/s12549-017-0299-7.
- Scheyer et al. (2014): Early Triassic Marine Biotic Recovery: The Predators' Perspective. PLoS ONE https://doi.org/10.1371/journal.pone.0088987
- Renesto, Silvio; Saller, Franco (2018). "EVIDENCES FOR A SEMI AQUATIC LIFE STYLE IN THE TRIASSIC DIAPSID REPTILE TANYSTROPHEUS". Rivista Italiana di Paleontologia e Stratigrafia (Research in Paleontology and Stratigraphy). 124 (1): N. 1 (2018). doi:10.13130/2039-4942/9541. ISSN 2039-4942.
- "Scientists follow the nose to solve mystery of long-necked reptile". the Guardian. 6 August 2020. Retrieved 8 August 2020.
Sources
- George Olshevsky expands on the history of "P." exogyrarum, on the Dinosaur Mailing List
- Huene, 1902. "Übersicht über die Reptilien der Trias" [Review of the Reptilia of the Triassic]. Geologische und Paläontologische Abhandlungen. 6, 1-84.
- Fritsch, 1905. "Synopsis der Saurier der böhm. Kreideformation" [Synopsis of the saurians of the Bohemian Cretaceous formation]. Sitzungsberichte der königlich-böhmischen Gesellschaft der Wissenschaften, II Classe. 1905(8), 1-7.