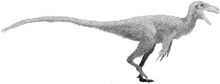
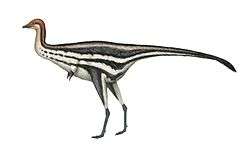
Oviraptoridae
Oviraptoridae is a group of bird-like, herbivorous and omnivorous maniraptoran dinosaurs. Oviraptorids are characterized by their toothless, parrot-like beaks and, in some cases, elaborate crests. They were generally small, measuring between one and two metres long in most cases, though some possible oviraptorids were enormous. Oviraptorids are currently known only from the Late Cretaceous of Asia, with the most well-known species and complete specimens found only in the Gobi Desert of Mongolia and northwestern China.
| Oviraptoridae | |
|---|---|
 | |
| Fossil skeleton of Khaan mckennai | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | Theropoda |
| Superfamily: | †Caenagnathoidea |
| Family: | †Oviraptoridae Barsbold, 1976 |
| Type species | |
| †Oviraptor philoceratops Osborn, 1924 | |
| Subgroups | |
| |
Description
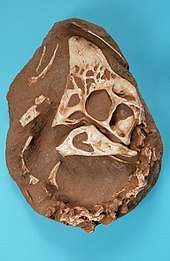
The most characteristic feature of this group is the skull structure. Oviraptorids had short snouts and very deep mandibles. Some taxa (Oviraptor, Citipati, Rinchenia) had a midline crest on top of the skull, resembling that of a cassowary. Other distinguishing characteristics include a bony spike intruding on the mandibular fenestra, nostrils placed very high and far back on the snout, an extremely thin bony bar beneath the eye, and highly pneumatized skull bones. Like their relatives the caenagnathids, the jaws were edentulous (with no teeth), having instead two small bony projections on the roof of the mouth.
Classification and species
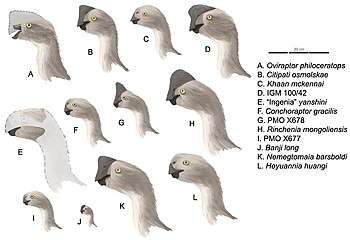
The classification of the oviraptorids has been controversial. Most studies divide oviraptorosaurs into two primary sub-groups, the Caenagnathidae and the Oviraptoridae. However, some phylogenetic studies have suggested that many traditional members of the Caenagnathidae may more closely related to the crested oviraptorids. Because of this, and the fact that at least one study found that Caenagnathus itself may not have been part of the 'caenagnathid' group, Tom Holtz (2010) placed that group close to the Oviraptoridae and termed it the Elmisauridae,[1] though this idea has not gained consensus among other researchers.
The Oviraptoridae itself is traditionally divided into two "subfamilies": the small, short-armed, and mainly crestless subfamily Heyuanniinae and the larger, crested, long-armed Oviraptorinae (Oviraptor + Citipati). Some phylogenetic studies have shown that Oviraptor is the most primitive known oviraptorid, thus making Citipati a closer relative of the "ingeniines" and this traditional division into crestless and crested forms artificial.[2]
Other possible oviraptorids include Nomingia gobiensis, Gigantoraptor erlianensis, Jiangxisaurus ganzhouensis and Shixinggia oblita. All four have been suggested to be oviraptorids, caenagnathids, or more primitive than either group. The below cladogram was found in the description of the genus Anzu.[2]
| Caenagnathoidea |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Paleobiology
Habitat
Almost all oviraptorids come from desert deposits of the Gobi Desert. Even in the late Cretaceous period, much of this area was desert, or at least very dry, habitat. In many of the localities where they are found, oviraptorids are among the most abundant dinosaurs present, second only to ankylosaurs and protoceratopsids. This is consistent with the idea that they were primarily herbivores, which tend to far outnumber carnivores in a given environment. Oviraptorids appear to have been far more abundant in arid habitats dominated mainly by small dinosaurs (such as those preserved in the Barun Goyot and Djadochta Formations) than in wetter ecosystems where large dinosaurs are common (such as the Nemegt Formation). The same pattern holds true for protoceratopsids, indicating that both groups preferred dry, desert-like habitat, and fed mainly on the types of tough, low-growing plant life that grows in arid climates.[3]
Diet
The diet of oviraptorids is not fully understood. Though some appear to have been at least partially carnivorous, they were probably primarily herbivorous.[3]
Originally, oviraptorids were thought to be specialized egg raiders, based on a Mongolian find showing Oviraptor on top of a nest erroneously attributed to the ceratopsian dinosaur Protoceratops. However, discoveries in the 1990s, including Citipati specimens clearly brooding (rather than preying on) the same types of nests, and a Citipati embryo inside the same type of egg preserved in these nests, showed that the "specialized egg thief" idea was incorrect.[4] Still, some scientists have suggested that oviraptorids may have fed on shelled food items like eggs or shellfish. However, animals specialized for eating shelled food typically have broad, crushing beaks or teeth. In contrast, the jaws of oviraptorids had thin, sharp edges probably supporting shearing beaks, ill-suited for cracking shells. Among other known animals, the beaks of oviraptorids most closely resemble those of herbivorous dicynodont synapsids, which are usually considered herbivorous. Their beaks also share similarities with the beaks of herbivorous parrots and tortoises.[3]
Evidence of partial carnivory among some oviraptorines comes from a lizard skeleton preserved in the body cavity of the type specimen of Oviraptor[5] and two hatchling Byronosaurus skulls found in a Citipati nest.[6] Some scientists have also suggested that some oviraptorids (especially the small-handed, weak-clawed "ingeniines") fed mainly on plant material.[3]
Reproduction

Although fossilized dinosaur eggs are generally rare, oviraptorid eggs are relatively well known. Several oviraptorid nests, eggs, and embryos are known, mostly uncovered in the Gobi Desert. Some specimens of Oviraptor philoceratops, Citipati osmolskae, Nemegtomaia and cf. Machairasaurus have been found in brooding positions in association with nests.[5][7][8] All of the nesting specimens are situated on top of egg clutches, with their limbs spread symmetrically on each side of the nest, front limbs covering the nest perimeter. This brooding posture is found today only in birds and supports a behavioral link between birds and theropod dinosaurs.[7]
Oviraptorid eggs are shaped like elongated ovals (elongatoolithid) and resemble the eggs of ratite birds (such as ostriches) in texture and shell structure. In the nest, eggs are typically found in pairs and arranged in concentric circles of up to three layers, with complete clutches consisting of as many of 22 eggs in some species.[9] The eggs of Citipati are the largest known definitive oviraptorid eggs, at 18 cm. In contrast, eggs associated with Oviraptor are only up to 14 cm long.[7]

The first oviraptorid eggs (of the genus Oviraptor, which mean "Egg thief") were found in close proximity to the remains of the ceratopsian dinosaur Protoceratops and it was assumed that the oviraptorids were preying upon the eggs of the ceratopsians.[10] It was not until 1993, when a Citipati embryo was discovered inside an egg of the type assigned to Protoceratops, that the error was corrected.[4] Norell et al., who recognized the embryo as oviraptorid, assigned it to the genus Citipati. The egg containing the embryo was smaller than most known Citipati eggs at only 12 cm, though it was partially eroded and broken into three pieces, making an accurate estimate of its original size difficult.[7] The embryo-bearing egg was otherwise identical to other oviraptorid eggs in shell structure and was found in an isolated nest, again arranged in a circular pattern.[4]

An oviraptorosaurian specimen from China described in 2005 was found to have two unlaid eggs within the pelvic canal. This suggests that, unlike modern crocodilians, oviraptorosaurs did not produce and lay many eggs at the same time. Rather, the eggs were produced within the reproductive organs in pairs, and laid two at a time, with the mother positioned in the center of the nest and rotating in a circle as each pair was laid. This behavior is supported by the fact that the eggs oval shape, with the more narrow end pointing backward from the birth canal, matching their orientation toward the center of the nest after being laid.[11]

The presence of two shelled eggs within the birth canal shows that oviraptorosaurs were intermediate between the reproductive biology of crocodilians and modern birds. Like crocodilians, they had two oviducts. However, crocodilians produce multiple shelled eggs per oviduct at a time, whereas oviraptorosaurs, like birds, produced only one egg per oviduct at a time.[11]
In 2017, paleontologists discovered colored pigments in some fossilized oviraptorid embryos of the egg-shell genus Macroolithus (which may represent eggs of Heyuannia). Examinations of eggs attributed to Heyuannia by Jasmina Wiemann and Tzu-Ruei Yang et al revealed the eggs preserved the blue-green pigment biliverdin and the reddish-brown pigment protoporphyrin, the same pigments found in many modern birds' eggshells. The eggs are thought to have been a blue-green color, because biliverdin is preserved in much greater abundance the photoporphyrin. In modern bird eggs, coloration can camouflage the eggs or help parents identify eggs, and it is correlated with more intensive parental care.[12]
Metabolism
A study by Robert Eagle et al. of the University of California-Los Angeles indicates that from specimens of eggs found in Mongolia and examination of the isotopes carbon-13 and oxygen 18 found within, Oviraptorids had body temperatures that could be elevated higher than that of the surrounding environment but lower than that of birds. This is very different from the isotope ratios of sauropod dinosaurs like Brachiosaurus, which had body temperatures of up to 100 °F (38 °C) and were fully endothermic.[13]
Feathers
Oviraptorids were probably feathered, since some close relatives were found with feathers preserved (including species of Caudipteryx, Protarchaeopteryx and Similicaudipteryx).[14][15] Another finding pointing to this is the discovery in Nomingia of a pygostyle, a bone that results from the fusion of the last tail vertebrae and is responsible in birds to hold a fan of feathers in the tail.[16] Finally, the arm position of the brooding Citipati would have been far more effective if feathers were present to cover the eggs.[17]
Pathology
The brooding oviraptorid specimen IGM 100/979 showed a callus and possible longitudinal groove left over from a healed fracture of the right ulna. Other oviraptorids have had pathological features reported in their phalanges but these have not been described in detail in the scientific literature.[18]
See also
References
- Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix.
- Lamanna, M. C.; Sues, H. D.; Schachner, E. R.; Lyson, T. R. (2014). "A New Large-Bodied Oviraptorosaurian Theropod Dinosaur from the Latest Cretaceous of Western North America". PLoS ONE. 9 (3): e92022. Bibcode:2014PLoSO...992022L. doi:10.1371/journal.pone.0092022. PMC 3960162. PMID 24647078.
- Nicholas R. Longrich; Philip J. Currie; Dong Zhi-Ming (2010). "A new oviraptorid (Dinosauria: Theropoda) from the Upper Cretaceous of Bayan Mandahu, Inner Mongolia". Palaeontology. 53 (5): 945–960. doi:10.1111/j.1475-4983.2010.00968.x.
- Norell, M. A., Clark, J. M., Dashzeveg, D., Barsbold, R., Chiappe, L. M., Davidson, A. R., McKenna, M. C., Altangerel, P. and Novacek, M. J. (November 1994). "A theropod dinosaur embryo and the affinities of the Flaming Cliffs Dinosaur eggs". Science. 266 (5186): 779–782. Bibcode:1994Sci...266..779N. doi:10.1126/science.266.5186.779. PMID 17730398.CS1 maint: multiple names: authors list (link)
- Norell, M.A.; Clark, J.M.; Chiappe, L.M.; Dashzeveg, D. (1995). "A nesting dinosaur". Nature. 378 (6559): 774–776. Bibcode:1995Natur.378..774N. doi:10.1038/378774a0.
- Bever, G.S. and Norell, M.A. (2009). "The perinate skull of Byronosaurus (Troodontidae) with observations on the cranial ontogeny of paravian theropods." American Museum Novitates, 3657: 51 pp.
- Clark, J.M., Norell, M.A., & Chiappe, L.M. (1999). "An oviraptorid skeleton from the Late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avianlike brooding position over an oviraptorid nest." American Museum Novitates, 3265: 36 pp., 15 figs.; (American Museum of Natural History) New York. (5.4.1999).
- Fanti, F.; Currie, P. J.; Badamgarav, D. (2012). Lalueza-Fox, Carles (ed.). "New Specimens of Nemegtomaia from the Baruungoyot and Nemegt Formations (Late Cretaceous) of Mongolia". PLoS ONE. 7 (2): e31330. Bibcode:2012PLoSO...731330F. doi:10.1371/journal.pone.0031330. PMC 3275628. PMID 22347465.
- Varricchio, D.J. (2000). "Reproduction and Parenting," in Paul, G.S. (ed.). The Scientific American Book of Dinosaurs. New York: St. Martin's Press, pp. 279-293.
- Osborn, H.F. (1924). "Three new Theropoda, Protoceratops zone, central Mongolia." American Museum Novitates, 144: 12 pp., 8 figs.; (American Museum of Natural History) New York. (11.7.1924).
- Sato, T.; Cheng, Y.; Wu, X.; Zelenitsky, D.K.; Hsaiao, Y. (2005). "A pair of shelled eggs inside a female dinosaur". Science. 308 (5720): 375. doi:10.1126/science.1110578. PMID 15831749.
- Wiemann, J.; Yang, T.-R.; Sander, P.N.; Schneider, M.; Engeser, M.; Kath-Schorr, S.; Müller, C.E.; Sander, P.M. (2017). "Dinosaur origin of egg color: oviraptors laid blue-green eggs". PeerJ. 5: e3706. doi:10.7717/peerj.3706. PMC 5580385. PMID 28875070.
- https://www.sciencedaily.com/releases/2015/10/151013135543.htm
- Ji, Q.; Currie, P.J.; Norell, M.A.; Ji, S. (1998). "Two feathered dinosaurs from northeastern China" (PDF). Nature. 393 (6687): 753–761. Bibcode:1998Natur.393..753Q. doi:10.1038/31635. Archived from the original (PDF) on 2008-12-17.
- Ji, Q., and Ji, S. (1997). "A Chinese archaeopterygian, Protarchaeopteryx gen. nov." Geological Science and Technology (Di Zhi Ke Ji), 238: 38-41. Translated By Will Downs Bilby Research Center Northern Arizona University January, 2001
- Barsbold, R.; Osmólska, H.; Watabe, M.; Currie, P.J.; Tsogtbaatar, K. (2000). "New Oviraptorosaur (Dinosauria, Theropoda) From Mongolia: The First Dinosaur With A Pygostyle". Acta Palaeontologica Polonica. 45 (2): 97–106.
- Paul, G.S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press.
- Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 337-363.