Hemiptera
The Hemiptera /hɛˈmɪptərə/ or true bugs are an order of insects comprising some 50,000 to 80,000 species[3] of groups such as the cicadas, aphids, planthoppers, leafhoppers, and shield bugs. They range in size from 1 mm (0.04 in) to around 15 cm (6 in), and share a common arrangement of sucking mouthparts.[4] The name "true bugs" is often limited to the suborder Heteroptera.[5] Many insects commonly known as "bugs", especially in American English, belong to other orders; for example, the lovebug is a fly[6] and the May bug and ladybug are beetles.[7]
| Hemiptera | |
|---|---|
 | |
| Acanthosoma haemorrhoidale, a shield bug | |
 | |
| Aphids | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Class: | Insecta |
| (unranked): | Paraneoptera |
| Superorder: | Condylognatha |
| Order: | Hemiptera Linnaeus, 1758 |
| Suborders[2] | |
Most hemipterans feed on plants, using their sucking and piercing mouthparts to extract plant sap. Some are hematophagous, while others are predators that feed on other insects or small invertebrates. They live in a wide variety of habitats, generally terrestrial, though some species are adapted to life in or on the surface of fresh water. Hemipterans are hemimetabolous, with young nymphs that somewhat resemble adults. Many aphids are capable of parthenogenesis, producing young from unfertilised eggs; this helps them to reproduce extremely rapidly in favourable conditions.
Humans have interacted with the Hemiptera for millennia. Some species, including many aphids, are significant agricultural pests, damaging crops by the direct action of sucking sap, but also harming them indirectly by being the vectors of serious viral diseases. Other species have been used for biological control of insect pests. Hemipterans have been cultivated for the extraction of the dyestuff cochineal (also known as carmine) and for shellac. The bed bug is a persistent parasite of humans, and some kissing bugs can transmit Chagas disease. Cicadas have been used as food, and have appeared in literature from the Iliad in Ancient Greece.
Diversity
Hemiptera is the largest order of hemimetabolous insects (not undergoing complete metamorphosis; though some examples such as male scale insects do undergo a form of complete metamorphosis [8]) containing over 75,000 named species; insect orders with more species all have a pupal stage (i.e. they do undergo complete metamorphosis or "holometabolism"), Coleoptera (370,000 described species), Lepidoptera (160,000), Diptera (100,000) and Hymenoptera (100,000).[9] The group is very diverse. The majority of species are terrestrial, including a number of important agricultural pests, but some are found in freshwater habitats. These include the water boatmen, pond skaters, and giant water bugs.[10]
Taxonomy and phylogeny

The fossil record of hemipterans goes back to the Carboniferous (Moscovian).[11] The oldest fossils are of the Archescytinidae from the Lower Permian and are thought to be basal to the Auchenorrhyncha. Fulguromorpha and Cicadomorpha appear in the Upper Permian, as do Sternorrhyncha of the Psylloidea and Aleurodoidea. Aphids and Coccoids appear in the Triassic. The Coleorrhyncha extend back to the Lower Jurassic.[12] The Heteroptera first appeared in the Triassic.[13]
The present members of the order Hemiptera (sometimes referred to as Rhynchota) were historically placed into two orders, the so-called Homoptera and Heteroptera/Hemiptera, based on differences in wing structure and the position of the rostrum. The order is now divided into four or more suborders, after the "Homoptera" were established as paraphyletic (now the Auchenorrhyncha and the Sternorrhyncha).[14][15][16] The cladogram is based on one analysis of the phylogeny of the Paraneoptera by Hu Li and colleagues in 2015, using mitochondrial genome sequences and homogeneous models. It places the Sternorrhyncha as sister clade to the Thysanoptera and the lice, making the Hemiptera as traditionally understood non-monophyletic.[17] However, when heterogeneous models were used, Hemiptera was found to be monophyletic.[17] The result where Hemiptera was found to be non-monophyletic is likely due to phylogenetic artifacts, such as elevated substitution rates in Sternorrhyncha compared with the other suborders of Hemiptera.[17] English names are given in parentheses where possible.
|
Other insects | |||||||||||||||||||||||||||||||||||||||||||
| Paraneoptera |
| ||||||||||||||||||||||||||||||||||||||||||
.jpg)
| Suborder | No. of Species | First appearance | Examples | Characteristics |
|---|---|---|---|---|
| Auchenorrhyncha | over 42,000[18] | Lower Permian | cicadas, leafhoppers, treehoppers, planthoppers, froghoppers | plant-sucking bugs; many can jump; many make calls, some loud |
| Coleorrhyncha | fewer than 30 | Lower Jurassic | moss bugs (Peloridiidae) | evolved in the southern palaeo-continent of Gondwana |
| Heteroptera | 25,000 | Triassic | shield bugs, seed bugs, assassin bugs, flower bugs, sweetpotato bugs, water bugs | larger bugs, often predatory |
| Sternorrhyncha | 12,500 | Upper Permian | aphids, whiteflies, scale insects | plant-sucking bugs, some major horticultural pests; most are sedentary or fully sessile;[19] |
Biology
Mouthparts
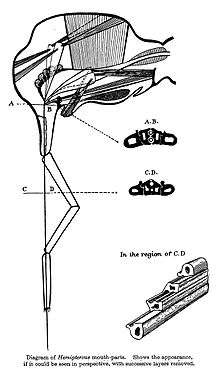
The defining feature of hemipterans is their "beak" in which the modified mandibles and maxillae form a "stylet" which is sheathed within a modified labium. The stylet is capable of piercing tissues and sucking liquids, while the labium supports it. The stylet contains a channel for the outward movement of saliva and another for the inward movement of liquid food. A salivary pump drives saliva into the prey; a cibarial pump extracts liquid from the prey. Both pumps are powered by substantial dilator muscles in the head. The beak is usually folded under the body when not in use. The diet is typically plant sap, but some hemipterans such as assassin bugs are blood-suckers, and a few are predators.[20][21]
Both herbivorous and predatory hemipterans inject enzymes to begin digestion extraorally (before the food is taken into the body). These enzymes include amylase to hydrolyse starch, polygalacturonase to weaken the tough cell walls of plants, and proteinases to break down proteins.[22]
Although the Hemiptera vary widely in their overall form, their mouthparts form a distinctive "rostrum". Other insect orders with mouthparts modified into anything like the rostrum and stylets of the Hemiptera include some Phthiraptera, but for other reasons they generally are easy to recognize as non-hemipteran. Similarly, the mouthparts of Siphonaptera, some Diptera and Thysanoptera superficially resemble the rostrum of the Hemiptera, but on closer inspection the differences are considerable. Aside from the mouthparts, various other insects can be confused with Hemiptera, but they all have biting mandibles and maxillae instead of the rostrum. Examples include cockroaches and psocids, both of which have longer, many-segmented antennae, and some beetles, but these have fully hardened forewings which do not overlap.[23]
Wing structure
The forewings of Hemiptera are either entirely membranous, as in the Sternorrhyncha and Auchenorrhyncha, or partially hardened, as in most Heteroptera. The name "Hemiptera" is from the Greek ἡμι- (hemi; "half") and πτερόν (pteron; "wing"), referring to the forewings of many heteropterans which are hardened near the base, but membranous at the ends. Wings modified in this manner are termed hemelytra (singular: hemelytron), by analogy with the completely hardened elytra of beetles, and occur only in the suborder Heteroptera. In all suborders, the hindwings – if present at all – are entirely membranous and usually shorter than the forewings.[10] The forewings may be held "roofwise" over the body (typical of Sternorrhyncha and Auchenorrhyncha),[24] or held flat on the back, with the ends overlapping (typical of Heteroptera).[10] The antennae in Hemiptera typically consist of four or five segments, although they can still be quite long, and the tarsi of the legs have two or three segments.[25]
Sound production
Many hemipterans can produce sound for communication.[26] The "song" of male cicadas, the loudest of any insect, is produced by tymbal organs on the underside of the abdomen, and is used to attract mates. The tymbals are drumlike disks of cuticle, which are clicked in and out repeatedly, making a sound in the same way as popping the metal lid of a jam jar in and out.[27]
Stridulatory sounds are produced among the aquatic Corixidae and Notonectidae (backswimmers) using tibial combs rubbed across rostral ridges.[28][29]
Life cycle

Hemipterans are hemimetabolous, meaning that they do not undergo metamorphosis, the complete change of form between a larval phase and an adult phase. Instead, their young are called nymphs, and resemble the adults to a greater or lesser degree. The nymphs moult several times as they grow, and each instar resembles the adult more than the previous one. Wing buds grow in later stage nymphs; the final transformation involves little more than the development of functional wings (if they are present at all) and functioning sexual organs, with no intervening pupal stage as in holometabolous insects.[30]
Parthenogenesis and viviparity

Many aphids are parthenogenetic during part of the life cycle, such that females can produce unfertilized eggs, which are clones of their mother. All such young are females (thelytoky), so 100% of the population at these times can produce more offspring. Many species of aphid are also viviparous: the young are born live rather than laid as eggs. These adaptations enable aphids to reproduce extremely rapidly when conditions are suitable.[31]
Locomotion

Hemipterans make use of a variety of modes of locomotion including swimming, skating on a water surface and jumping, as well as walking and flying like other insects.
Swimming and skating
Several families of Heteroptera are water bugs, adapted to an aquatic lifestyle, such as the water boatmen (Corixidae), water scorpions (Nepidae), and backswimmers (Notonectidae). They are mostly predatory, and have legs adapted as paddles to help the animal move through the water.[3] The pondskaters or water striders (Gerridae) are also associated with water, but use the surface tension of standing water to keep them above the surface;[32] they include the sea skaters in the genus Halobates, the only truly marine group of insects.[3]

Marangoni propulsion
Marangoni effect propulsion exploits the change in surface tension when a soap-like surfactant is released on to a water surface, in the same way that a toy soap boat propels itself. Water bugs in the genus Microvelia (Veliidae) can travel at up to 17 cm/s, twice as fast as they can walk, by this means.[32]
Flight
Flight is well developed in the Hemiptera although mostly used for short distance movement and dispersal. Wing development is sometimes related to environmental conditions. In some groups of Hemiptera, there are variations of winged, short-winged, and wingless forms within a single species. This kind of polymorphism tends to be helpful when habitats are temporary with more energy put into reproduction when food is available and into dispersal through flight when food becomes scarce. In aphids, both winged and wingless forms occur with winged forms produced in greater numbers when food resources are depleted. Aphids and whiteflies can sometimes be transported very long distances by atmospheric updrafts and high altitude winds.[33] Wing-length polymorphism is notably rare in tree-living Hemiptera.[34]
Jumping
Many Auchenorrhyncha including representatives of the cicadas, leafhoppers, treehoppers, planthoppers, and froghoppers are adapted for jumping (saltation). Treehoppers, for example, jump by rapidly depressing their hind legs. Before jumping, the hind legs are raised and the femora are pressed tightly into curved indentations in the coxae. Treehoppers can attain a take-off velocity of up to 2.7 metres per second and an acceleration of up to 250 g. The instantaneous power output is much greater than that of normal muscle, implying that energy is stored and released to catapult the insect into the air.[35] Cicadas, which are much larger, extend their hind legs for a jump in under a millisecond, again implying elastic storage of energy for sudden release.[36]
Ecological roles
Feeding modes
Herbivores
.jpg)
Most hemipterans are phytophagous, using their sucking and piercing mouthparts to feed on plant sap. These include cicadas, leafhoppers, treehoppers, planthoppers, froghoppers, aphids, whiteflies, scale insects, and some other groups. Some are monophages, being host specific and only found on one plant taxon, others are oligophages, feeding on a few plant groups, while others again are less discriminating polyphages and feed on many species of plant.[21] The relationship between hemipterans and plants appears to be ancient, with piercing and sucking of plants evident in the Early Devonian period.[38]
Hemipterans can dramatically cut the mass of affected plants, especially in major outbreaks. They sometimes also change the mix of plants by predation on seeds or feeding on roots of certain species.[39] Some sap-suckers move from one host to another at different times of year. Many aphids spend the winter as eggs on a woody host plant and the summer as parthogenetically reproducing females on a herbaceous plant.[40]
.jpg)
Phloem sap, which has a higher concentration of sugars and nitrogen, is under positive pressure unlike the more dilute xylem sap. Most of the Sternorrhyncha and a number of Auchenorrhynchan groups feed on phloem. Phloem feeding is common in the Fulgoromorpha, most Cicadellidae and in the Heteroptera. The Typhlocybine Cicadellids specialize in feeding on non-vascular mesophyll tissue of leaves, which is more nutritious than the leaf epidermis. Most Heteroptera also feed on mesophyll tissue where they are more likely to encounter defensive secondary plant metabolites which often leads to the evolution of host specificity.[41] Obligate xylem feeding is a special habit that is found in the Auchenorrhyncha among Cicadoidea, Cercopoidea and in Cicadelline Cicadellids. Some phloem feeders may take to xylem sap facultatively, especially when facing dehydration.[42] Xylem feeders tend to be polyphagous;[43] to overcome the negative pressure of xylem requires a special cibarial pump.[44] Phloem feeding hemiptera typically have symbiotic micro-organisms in their gut that help to convert amino acids. Phloem feeders produce honeydew from their anus. A variety of organisms that feed on honeydew form symbiotic associations with phloem-feeders.[45][46] Phloem sap is a sugary liquid low in amino acids, so insects have to process large quantities to meet their nutritional requirements. Xylem sap is even lower in amino acids and contains monosaccharides rather than sucrose, as well as organic acids and minerals. No digestion is required (except for the hydrolysis of sucrose) and 90% of the nutrients in the xylem sap can be utilised.[21][47] Some phloem sap feeders selectively mix phloem and xylem sap to control the osmotic potential of the liquid consumed.[48] A striking adaptation to a very dilute diet is found in many hemipterans: a filter chamber, a part of the gut looped back on itself as a countercurrent exchanger, which permits nutrients to be separated from excess water.[49] The residue, mostly water with sugars and amino acids, is quickly excreted as sticky "honey dew", notably from aphids but also from other Auchenorrhycha and Sternorrhyncha.[50]
Some Sternorrhyncha including Psyllids and some aphids are gall formers. These sap-sucking hemipterans inject fluids containing plant hormones into the plant tissues inducing the production of tissue that covers to protects the insect and also act as sinks for nutrition that they feed on. The hackleberry gall psyllid for example, causes a woody gall on the leaf petioles of the hackleberry tree it infests,[51] and the nymph of another psyllid produces a protective lerp out of hardened honeydew.[21]
Predators
Most other hemipterans are predatory, feeding on other insects, or even small vertebrates. This is true of many aquatic species which are predatory, either as nymphs or adults.[25] The predatory shield bug for example stabs caterpillars with its beak and sucks out the body fluids.[52] The saliva of predatory heteropterans contains digestive enzymes such as proteinase and phospholipase, and in some species also amylase. The mouthparts of these insects are adapted for predation. There are toothed stylets on the mandibles able to cut into and abrade tissues of their prey. There are further stylets on the maxillae, adapted as tubular canals to inject saliva and to extract the pre-digested and liquified contents of the prey.[53]
Some species attack pest insects and are used in biological control. One of these is the spined soldier bug (Podisus maculiventris) that sucks body fluids from larvae of the Colorado beetle and the Mexican bean beetle.[54]
Haematophagic "parasites"
A few hemipterans are haematophagic (often described as "parasites"[55]), feeding on the blood of larger animals. These include bedbugs and the triatomine kissing bugs of the assassin bug family Reduviidae, which can transmit the dangerous Chagas disease.[3] The first known hemipteran to feed in this way on vertebrates was the extinct assassin bug Triatoma dominicana found fossilized in amber and dating back about twenty million years. Faecal pellets fossilised beside it show that it transmitted a disease-causing Trypanosoma and the amber included hairs of the likely host, a bat.[56]
As symbionts

Some species of ant protect and farm aphids (Sternorrhyncha) and other sap-sucking hemipterans, gathering and eating the honeydew that these hemipterans secrete. The relationship is mutualistic, as both ant and aphid benefit. Ants such as the yellow anthill ant, Lasius flavus, breed aphids of at least four species, Geoica utricularia, Tetraneura ulmi, Forda marginata and Forda formicaria, taking eggs with them when they found a new colony; in return, these aphids are obligately associated with the ant, breeding mainly or wholly asexually inside anthills.[57] Ants may also protect the plant bugs from their natural enemies, removing the eggs of predatory beetles and preventing access by parasitic wasps.[21]
Some leafhoppers (Auchenorrhyncha) are similarly "milked" by ants. In the Corcovado rain forest of Costa Rica, wasps compete with ants to protect and milk leafhoppers; the leafhoppers preferentially give more honeydew, more often, to the wasps, which are larger and may offer better protection.[58]
As prey: defences against predators and parasites

Hemiptera form prey to predators including vertebrates, such as birds, and other invertebrates such as ladybirds.[59][60] In response, hemipterans have evolved antipredator adaptations. Ranatra may feign death (thanatosis). Others such as Carpocoris purpureipennis secrete toxic fluids to ward off arthropod predators; some Pentatomidae such as Dolycoris are able to direct these fluids at an attacker. Toxic cardenolide compounds are accumulated by the heteropteran Oncopeltus fasciatus when it consumes milkweeds, while the coreid stinkbug Amorbus rubiginosus acquires 2-hexenal from its food plant, Eucalyptus. Some long-legged bugs mimic twigs, rocking to and fro to simulate the motion of a plant part in the wind.[60] The nymph of the Masked hunter bug camouflages itself with sand grains, using its hind legs and tarsal fan to form a double layer of grains, coarser on the outside.[61] The Amazon rain forest cicada Hemisciera maculipennis display bright red deimatic flash coloration on their hindwings when threatened; the sudden contrast helps to startle predators, giving the cicadas time to escape. The coloured patch on the hindwing is concealed at rest by an olive green patch of the same size on the forewing, enabling the insect to switch rapidly from cryptic to deimatic behaviour.[62][lower-alpha 1]

Some hemipterans such as firebugs have bold aposematic warning coloration, often red and black, which appear to deter passerine birds.[64][65] Many hemipterans including aphids, scale insects and especially the planthoppers secrete wax to protect themselves from threats such as fungi, parasitoidal insects and predators, as well as abiotic factors like desiccation.[66] Hard waxy coverings are especially important in the sedentary Sternorrhyncha such as scale insects, which have no means of escaping from predators; other Sternorrhyncha evade detection and attack by creating and living inside plant galls.[37] Nymphal Cicadoidea and Cercopoidea have glands attached to the Malpighian tubules in their proximal segment that produce mucopolysaccharides, which form the froth around spittlebugs, offering a measure of protection.[67]
Parental care is found in many species of Hemiptera especially in members of the Membracidae and numerous Heteroptera. In many species of shield bug, females stand guard over their egg clusters to protect them from egg parasitoids and predators.[68] In the aquatic Belostomatidae, females lay their eggs on the back of the male which guards the eggs.[69] Protection provided by ants is common in the Auchenorrhyncha.[21]
Interaction with humans

As pests
Although many species of Hemiptera are significant pests of crops and garden plants, including many species of aphid and scale insects, other species are harmless. The damage done is often not so much the deprivation of the plant of its sap, but the fact that they transmit serious viral diseases between plants.[9] They often produce copious amounts of honeydew which encourages the growth of sooty mould.[70] Significant pests include the cottony cushion scale, a pest of citrus fruit trees,[71] the green peach aphid and other aphids which attack crops worldwide and transmit diseases,[72] and jumping plant lice which are often host plant-specific and transmit diseases.
For pest control
Members of the families Reduviidae, Phymatidae and Nabidae are obligate predators. Some predatory species are used in biological pest control; these include various nabids,[73] and even some members of families that are primarily phytophagous, such as the genus Geocoris in the family Lygaeidae.[74] Other hemipterans are omnivores, alternating between a plant-based and an animal-based diet. For example, Dicyphus hesperus is used to control whitefly on tomatoes but also sucks sap, and if deprived of plant tissues will die even if in the presence of whiteflies.[75]
Insect products

Other hemipterans have positive uses for humans, such as in the production of the dyestuff carmine (cochineal). The FDA has created guidelines for how to declare when it has been added to a product.[76] The scale insect Dactylopius coccus produces the brilliant red-coloured carminic acid to deter predators. Up to 100,000 scale insects need to be collected and processed to make a kilogram (2.2 lbs) of cochineal dye.[77] A similar number of lac bugs are needed to make a kilogram of shellac, a brush-on colourant and wood finish.[78] Additional uses of this traditional product include the waxing of citrus fruits to extend their shelf-life, and the coating of pills to moisture-proof them, provide slow-release or mask the taste of bitter ingredients.[79]
As human parasites and disease vectors

Chagas disease is a modern-day tropical disease caused by Trypanosoma cruzi and transmitted by kissing bugs, so-called because they suck human blood from around the lips while a person sleeps.[80]
The bed bug, Cimex lectularius, is an external parasite of humans. It lives in bedding and is mainly active at night, feeding on human blood, generally without being noticed.[81][82] Bed bugs mate by traumatic insemination; the male pierces the female's abdomen and injects his sperm into a secondary genital structure, the spermalege. The sperm travel in the female's blood (haemolymph) to sperm storage structures (seminal conceptacles); they are released from there to fertilise her eggs inside her ovaries.[82][83]
As food

Some larger hemipterans such as cicadas are used as food in Asian countries such as China,[84] and they are much esteemed in Malawi and other African countries. Insects have a high protein content and good food conversion ratios, but most hemipterans are too small to be a useful component of the human diet.[85] At least nine species of Hemiptera are eaten worldwide.[86]
In art and literature
Cicadas have featured in literature since the time of Homer's Iliad, and as motifs in decorative art from the Chinese Shang dynasty (1766–1122 B.C.). They are described by Aristotle in his History of Animals and by Pliny the Elder in his Natural History; their mechanism of sound production is mentioned by Hesiod in his poem Works and Days "when the Skolymus flowers, and the tuneful Tettix sitting on his tree in the weary summer season pours forth from under his wings his shrill song".[87]
In mythology and folklore
Among the bugs, cicadas in particular have been used as money, in folk medicine, to forecast the weather, to provide song (in China), and in folklore and myths around the world.[88]
Threats
Large-scale cultivation of the oil palm Elaeis guineensis in the Amazon basin damages freshwater habitats and reduces the diversity of aquatic and semi-aquatic Heteroptera.[89] Climate change may be affecting the global migration of hemipterans including the potato leafhopper, Empoasca fabae. Warming is correlated with the severity of potato leafhopper infestation, so increased warming may worsen infestations in future.[90]
Notes
- The green/red flash coloration earns the insect the name of stop and go cicada.[63]
References
- Wang, Yan-hui; Engel, Michael S.; Rafael, José A.; Wu, Hao-yang; Rédei, Dávid; Xie, Qiang; Wang, Gang; Liu, Xiao-guang; Bu, Wen-jun (2016). "Fossil record of stem groups employed in evaluating the chronogram of insects (Arthropoda: Hexapoda)". Scientific Reports. 6: 38939. Bibcode:2016NatSR...638939W. doi:10.1038/srep38939. PMC 5154178. PMID 27958352.
- "Hemiptera". Integrated Taxonomic Information System.
- Jon Martin; Mick Webb. "Hemiptera...It's a Bug's Life" (PDF). Natural History Museum. Retrieved July 26, 2010.
- "Hemiptera: bugs, aphids and cicadas". Commonwealth Scientific and Industrial Research Organisation. Retrieved May 8, 2007.
- "Suborder Heteroptera – True Bugs". Bug guide. Iowa State University Entomology. n.d.
- Denmark, Harold; Mead, Frank; Fasulo, Thomas (April 2010). "Lovebug, Plecia nearctica Hardy". Featured Creatures. University of Florida/IFAS. Retrieved 22 September 2010.
- "Melolontha melolontha (cockchafer or May bug)". Natural History Museum. Retrieved 12 July 2015.
- Gullan, Penny; Kosztarab, Michael (1997). "Adaptations in scale insects". Annual Review of Entomology. 42: 23–50. doi:10.1146/annurev.ento.42.1.23. PMID 15012306.
- "Insect groups (Orders)". Amateur Entomologists' Society. Retrieved 16 July 2015.
- Coulson, Robert N.; Witter, John A. (1984). Forest Entomology: Ecology and Management. John Wiley & Sons. pp. 71–72. ISBN 978-0-471-02573-3.
- André Nel; Patrick Roques; Patricia Nel; Alexander A. Prokin; Thierry Bourgoin; Jakub Prokop; Jacek Szwedo; Dany Azar; Laure Desutter-Grandcolas; Torsten Wappler; Romain Garrouste; David Coty; Diying Huang; Michael S. Engel; Alexander G. Kirejtshuk (2013). "The earliest known holometabolous insects". Nature. 503 (7475): 257–261. Bibcode:2013Natur.503..257N. doi:10.1038/nature12629. PMID 24132233.
- Gillott, Cedric (2005). Entomology (3 ed.). Springer. p. 213.
- Shcherbakov, D. E. (2000). "Permian faunas of Homoptera (Hemiptera) in relation to phytogeography and the Permo-Triassic crisis" (PDF). Paleontological Journal. 34 (3): S251–S267.
- Song, Nan; Liang, Ai-Ping; Bu, Cui-Ping (2012). "A Molecular Phylogeny of Hemiptera Inferred from Mitochondrial Genome Sequences". PLOS ONE. 7 (11): e48778. Bibcode:2012PLoSO...748778S. doi:10.1371/journal.pone.0048778. PMC 3493603. PMID 23144967.
- Forero, Dimitri (2008). "The Systematics of Hemiptera" (PDF). Revista Colombiana de Entomologia. 34 (1): 1–21. Archived from the original (PDF) on January 13, 2015.
- Cryan, Jason C.; Urban, Julie M. (January 2012). "Higher-level phylogeny of the insect order Hemiptera: is Auchenorrhyncha really paraphyletic?". Systematic Entomology. 37 (1): 7–21. doi:10.1111/j.1365-3113.2011.00611.x.
- Li, Hu; et al. (2015). "Higher-level phylogeny of paraneopteran insects inferred from mitochondrial genome sequences". Scientific Reports. 5: 8527. Bibcode:2015NatSR...5E8527L. doi:10.1038/srep08527. PMC 4336943. PMID 25704094.
- "Suborder Auchenorrhyncha". NCSU. Retrieved 12 July 2015.
- "Sternorrhyncha". Amateur Entomologists' Society. Retrieved 13 July 2015.
- Ruppert, Edward E.; Fox, Richard, S.; Barnes, Robert D. (2004). Invertebrate Zoology, 7th edition. Cengage Learning. pp. 728, 748. ISBN 978-81-315-0104-7.
- Gullan, P.J.; Cranston, P.S. (2014). The Insects: An Outline of Entomology, 5th Edition. Wiley. pp. 80–81, 790–. ISBN 978-1-118-84616-2.
- Wheeler, Alfred George (2001). Biology of the Plant Bugs (Hemiptera: Miridae): Pests, Predators, Opportunists. Cornell University Press. pp. 105–135. ISBN 978-0-8014-3827-1.
- Chinery, Michael (1993). Insects of Britain and Northern Europe (3rd ed.). Collins. ISBN 978-0-00-219918-6.
- Alford, David V. (2012). Pests of Ornamental Trees, Shrubs and Flowers: A Color Handbook. Academic Press. p. 12. ISBN 978-0-12-398515-6.
- "Hemiptera". Discover Life. Retrieved 13 July 2015.
- Virant-Doberlet, Meta; Cokl, Andrej (April 2004). "Vibrational Communication in Insects". Neotropical Entomology. 33 (2): 121–134. doi:10.1590/S1519-566X2004000200001.
- Simmons, Peter; Young, David (1978). "The Tymbal Mechanism and Song Patterns of the Bladder Cicada, Cystosoma saundersii" (PDF). Journal of Experimental Biology. 76: 27–45. Archived from the original (PDF) on 2015-12-24. Retrieved 2015-08-22.
- Wilcox, R. Stimson (1975). "Sound-producing Mechanisms of Bueno macrotibialis Hungerford (Hemiptera: Notonectidae)" (PDF). International Journal of Insect Morphology & Embryology. 4 (21): 169–182. doi:10.1016/0020-7322(75)90015-x. hdl:2027.42/22083.
- Gooderham, John; Tsyrlin, Edward (2002). The Waterbug Book: A Guide to the Freshwater Macroinvertebrates of Temperate Australia. Csiro Publishing. p. 146. ISBN 978-0-643-06668-7.
- Britton, David (9 July 2009). "Metamorphosis: a remarkable change". Australian Museum. Retrieved 13 July 2015.
- Mackean, D.G. "Aphids, an Introduction". Biology Teaching Resources. Retrieved 13 July 2015.
- Bush, John W. M.; Hu, David L. (2006). "Walking on Water: Biolocomotion at the Interface". Annual Review of Fluid Mechanics. 38 (1): 339–369. Bibcode:2006AnRFM..38..339B. doi:10.1146/annurev.fluid.38.050304.092157.
- Loxdale, H. D.; Hardie, J.; Halbert, S.; Foottit, R.; Kidd, N. A. C.; Carter, C. I. (1993). "The relative importance of short- and long-range movement of flying aphids". Biological Reviews. 68 (2): 291–311. doi:10.1111/j.1469-185X.1993.tb00998.x.
- Waloff, N. (1983). "Absence of wing polymorphism in the arboreal, phytophagous species of some taxa of temperate Hemiptera: An hypothesis". Ecological Entomology. 8 (2): 229–232. doi:10.1111/j.1365-2311.1983.tb00502.x.
- Burrows, M. (March 2013). "Jumping mechanisms of treehopper insects (Hemiptera, Auchenorrhyncha, Membracidae)". Journal of Experimental Biology. 216 (5): 788–799. doi:10.1242/jeb.078741. PMID 23155084.
- Gorb, S. N. (July 2004). "The jumping mechanism of cicada Cercopis vulnerata (Auchenorrhyncha, Cercopidae): skeleton-muscle organisation, frictional surfaces, and inverse-kinematic model of leg movements". Arthropod Structure & Development. 33 (3): 201–220. doi:10.1016/j.asd.2004.05.008. PMID 18089035.
- Grimaldi, David; Engel, Michael S. (16 May 2005). Evolution of the Insects. Cambridge University Press. p. 289. ISBN 978-0-521-82149-0.
- Labandeira, Conrad C. (May 1998). "Early History of Arthropod and Vascular Plant Associations". Annual Review of Earth and Planetary Sciences. 26: 329–377. Bibcode:1998AREPS..26..329L. doi:10.1146/annurev.earth.26.1.329.
- Verne, Nole C. (2007). Forest Ecology Research Horizons. Nova Publishers. p. 210. ISBN 978-1-60021-490-5.
- Dixon, A.F.G. (2012). Aphid Ecology: An optimization approach. Springer Science & Business Media. p. 128. ISBN 978-94-011-5868-8.
- Tonkyn, David W.; Whitcomb, Robert F. (1987). "Feeding Strategies and the Guild Concept Among Vascular Feeding Insects and Microorganisms". In Harris, Kerry F. (ed.). Current Topics in Vector Research. pp. 179–199. doi:10.1007/978-1-4612-4712-8_6. ISBN 978-1-4612-9126-8.
- Pompon, Julien; Quiring, Dan; Goyer, Claudia; Giordanengo, Philippe; Pelletier, Yvan (2011). "A phloem-sap feeder mixes phloem and xylem sap to regulate osmotic potential" (PDF). Journal of Insect Physiology. 57 (9): 1317–1322. doi:10.1016/j.jinsphys.2011.06.007. PMID 21726563. Archived from the original (PDF) on 2015-09-23. Retrieved 2015-08-22.
- Den, Francesca F.; Stewart, Alan J. A.; Gibson, Amos; Weiblen, George D.; Novotny, Vojtech (2013). "Low host specificity in species-rich assemblages of xylem- and phloem-feeding herbivores (Auchenorrhyncha) in a New Guinea lowland rain forest" (PDF). Journal of Tropical Ecology. 29 (6): 467–476. doi:10.1017/S0266467413000540.
- Novotny, Vojtech; Wilson, Michael R. (1997). "Why are there no small species among xylem-sucking insects?". Evolutionary Ecology. 11 (4): 419–437. doi:10.1023/a:1018432807165.
- Douglas, A.E. (2006). "Phloem-sap feeding by animals: problems and solutions" (PDF). Journal of Experimental Botany. 57 (4): 747–754. doi:10.1093/jxb/erj067. PMID 16449374.
- Baje, Leontine; Stewart, A.J.A.; Novotny, Vojtech (2014). "Mesophyll cell-sucking herbivores (Cicadellidae: Typhlocybinae) on rainforest trees in Papua New Guinea: local and regional diversity of a taxonomically unexplored guild Ecological Entomology". Ecological Entomology. 39 (3): 325–333. doi:10.1111/een.12104.
- Panizzi, Antônio Ricardo; Parra, José R.P. (2012). Insect Bioecology and Nutrition for Integrated Pest Management. CRC Press. p. 108. ISBN 978-1-4398-3708-5.
- Pompon, Julien; Quiring, Dan; Goyer, Claudia; Giordanengo, Philippe; Pelletier, Yvan (2011). "A phloem-sap feeder mixes phloem and xylem sap to regulate osmotic potential" (PDF). Journal of Insect Physiology. 57 (9): 1317–1322. doi:10.1016/j.jinsphys.2011.06.007. PMID 21726563. Archived from the original (PDF) on 2015-09-23. Retrieved 2015-08-22.
- "Digestive System". Entomology. University of Sydney. Retrieved 22 August 2015.
- Underwood, D. L. A. "Hemipteroids" (PDF). California State University. Retrieved 22 August 2015.
- Hall, Donald W.; Butler, Jerry F.; Cromroy, Harvey L. (1 September 2014). "Hackberry petiole gall psyllid". Featured Creatures. University of Florida/IFAS. Retrieved 12 August 2015.
- "Glossy Shield Bug: Cermatulus nasalis". Brisbane Insects and Spiders. 2010. Retrieved 16 July 2015.
- Cohen, Allen C. (1990). "Feeding Adaptations of Some Predaceous Hemiptera" (PDF). Annals of the Entomological Society of America. 83 (6): 1215–1223. doi:10.1093/aesa/83.6.1215.
- Coll, M.; Ruberson, J.R., eds. (1998). Predatory Heteroptera: their ecology and use in biological control. ISBN 978-0-938522-62-1.
- Goddard, Jerome (2009). "Bed Bugs (Cimex lectularius) and Clinical Consequences of Their Bites". JAMA. 301 (13): 1358–1366. doi:10.1001/jama.2009.405. PMID 19336711.
- Poinar, G. (2005). "Triatoma dominicana sp. n. (Hemiptera: Reduviidae: Triatominae), and Trypanosoma antiquus sp. n. (Stercoraria: Trypanosomatidae), the First Fossil Evidence of a Triatomine-Trypanosomatid Vector Association". Vector-Borne and Zoonotic Diseases. 5 (1): 72–81. doi:10.1089/vbz.2005.5.72. PMID 15815152.
- Ivens, A. B. F.; Kronauer, D. J. C.; Pen, I.; Weissing, F. J.; Boomsma, J. J. (2012). "Reproduction and dispersal in an ant-associated root aphid community". Molecular Ecology. 21 (17): 4257–69. doi:10.1111/j.1365-294X.2012.05701.x. PMID 22804757.
- Choe, Jae (14 February 2012). Secret Lives of Ants. JHU Press. pp. 62–64. ISBN 978-1-4214-0521-6.
- Evans, Edward W. (November 2009). "Lady beetles as predators of insects other than Hemiptera". Biological Control. 51 (2): 255–267. doi:10.1016/j.biocontrol.2009.05.011.
- Edmunds, M. (1974). Defence in Animals. Longman. pp. 172, 191–194, 201, 261 and passim. ISBN 978-0-582-44132-3.
- Wierauch C. (2006). "Anatomy of Disguise: Camouflaging Structures in Nymphs of Some Reduviidae (Heteroptera)". Am. Mus. Novit. 3542: 1–18. doi:10.1206/0003-0082(2006)3542[1:aodcsi]2.0.co;2. hdl:2246/5820.
- Cott, Hugh B. (1940). Adaptive Coloration in Animals. Methuen. pp. 375–376.
- "Better IDs for E.A. Seguy Cicada Illustrations". Retrieved 24 August 2015.
- Wheeler, Alfred George (2001). Biology of the Plant Bugs (Hemiptera: Miridae): Pests, Predators, Opportunists. Cornell University Press. pp. 100ff. ISBN 978-0-8014-3827-1.
- Exnerova, Alice; et al. (April 2003). "Reactions of passerine birds to aposematic and non-aposematic firebugs (Pyrrhocoris apterus ; Heteroptera)". Biological Journal of the Linnean Society. 78 (4): 517–525. doi:10.1046/j.0024-4066.2002.00161.x.
- Lucchi, Andrea; Mazzoni, Emanuele (2004). "Wax Production in Adults of Planthoppers (Homoptera: Fulgoroidea) with Particular Reference to Metcalfa pruinosa (Flatidae)" (PDF). Annals of the Entomological Society of America. 97 (6): 1294–1298. doi:10.1603/0013-8746(2004)097[1294:wpiaop]2.0.co;2.
- Rakitov, Roman A. (2002). "Structure and Function of the Malpighian Tubules, and Related Behaviors in Juvenile Cicadas: Evidence of Homology with Spittlebugs (Hemiptera: Cicadoidea & Cercopoidea)". Zoologischer Anzeiger. 241 (2): 117–130. doi:10.1078/0044-5231-00025.
- Tallamy, Douglas W.; Schaefer, Carl (1997). "Maternal care in the Hemiptera: ancestry, alternatives, and current adaptive value". In Jae C. Choe; Bernard J. Crespi (eds.). The Evolution of Social Behaviour in Insects and Arachnids. Cambridge University Press. pp. 91–115.
- Crowl, T.A.; Alexander Jr., J.E. (1989). "Parental care and foraging ability in male water bugs (Belostoma flumineum)". Canadian Journal of Zoology. 67: 513–515. doi:10.1139/z89-074.
- "Sooty moulds". Royal Horticultural Society. Retrieved 16 July 2015.
- David L. Green (10 August 2003). "Cottony cushion scale: The pest that launched a revolution in pest control methods". Retrieved 16 July 2015.
- Capinera, John L. (October 2005). "Featured creatures". University of Florida website – Department of Entomology and Nematology. University of Florida. Retrieved 16 July 2015.
- Mahr, Susan (1997). "Know Your Friends: Damsel Bugs". Biological Control News. IV (2). Retrieved 16 July 2015.
- James Hagler. Weeden, Catherine R.; Shelton, Anthony M.; Hoffman, Michael P. (eds.). "Geocoris spp. (Heteroptera: Lygaeidae): Bigeyed Bug". Biological Control: A Guide to Natural Enemies in North America. Cornell University. Retrieved 16 July 2015.
- Torres, Jorge Braz; Boyd, David W. (2009). "Zoophytophagy in predatory Hemiptera". Brazilian Archives of Biology and Technology. 52 (5): 1199–1208. doi:10.1590/S1516-89132009000500018.
- FDA Color Additives,"Guidance for Industry: Cochineal Extract and Carmine: Declaration by Name on the Label of All Foods and Cosmetic Products That Contain These Color Additives; Small Entity Compliance Guide". www.fda.gov. Retrieved 2016-02-22.
- "Cochineal and Carmine". Major colourants and dyestuffs, mainly produced in horticultural systems. FAO. Retrieved June 16, 2015.
- "How Shellac Is Manufactured". The Mail (Adelaide, SA : 1912 – 1954). 18 Dec 1937. Retrieved 17 July 2015.
- Pearnchob, N.; Siepmann, J.; Bodmeier, R. (2003). "Pharmaceutical applications of shellac: moisture-protective and taste-masking coatings and extended-release matrix tablets". Drug Development and Industrial Pharmacy. 29 (8): 925–938. doi:10.1081/ddc-120024188. PMID 14570313.
- "American Trypanosomiasis". Centers for Disease Control (CDC). Retrieved 17 July 2015.
- Goddard, J; deShazo, R (1 April 2009). "Bed bugs (Cimex lectularius) and clinical consequences of their bites". JAMA: The Journal of the American Medical Association. 301 (13): 1358–66. doi:10.1001/jama.2009.405. PMID 19336711.
- Reinhardt, Klaus; Siva-Jothy, Michael T. (Jan 2007). "Biology of the Bed Bugs (Cimicidae)" (PDF). Annual Review of Entomology. 52: 351–374. doi:10.1146/annurev.ento.52.040306.133913. PMID 16968204. Archived from the original (PDF) on 5 July 2010. Retrieved 26 May 2010.
- Carayon, J. 1959 Insémination par "spermalège" et cordon conducteur de spermatozoids chez Stricticimex brevispinosus Usinger (Heteroptera, Cimicidae). Rev. Zool. Bot. Afr. 60, 81–104.
- Greenaway, Twilight. "The Best Way to Handle the Coming Cicada Invasion? Heat Up the Deep Fryer". Smithsonian. Retrieved 12 July 2015.
- Anthes, Emily (14 October 2014). "Could insects be the wonder food of the future?". BBC. Retrieved 17 July 2015.
- "Future Prospects for Food and Feed Security". Retrieved 13 April 2016.
- Myers, J. G. (1929). Insect Singers (PDF). G. Routledge and Sons.
- "Cicada". Britannica. Retrieved 12 July 2015.
- Cunha, Erlane Jose; Montag, Luciano; Juen, Leandro (2015). "Oil palm crops effects on environmental integrity of Amazonian streams and Heteropteran (Hemiptera) species diversity". Ecological Indicators. 52: 422–429. doi:10.1016/j.ecolind.2014.12.024.
- Baker, Mitchell B.; Venugopal, P. Dilip; Lamp, William O. (2015). "Climate Change and Phenology: Empoasca fabae (Hemiptera: Cicadellidae) Migration and Severity of Impact". PLOS ONE. 10 (5): e0124915. Bibcode:2015PLoSO..1024915B. doi:10.1371/journal.pone.0124915. PMC 4430490. PMID 25970705.
External links
| Wikimedia Commons has media related to Hemiptera. |
| Wikispecies has information related to Hemiptera |
Insect orders | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
