FTO gene
Fat mass and obesity-associated protein also known as alpha-ketoglutarate-dependent dioxygenase FTO is an enzyme that in humans is encoded by the FTO gene located on chromosome 16. As one homolog in the AlkB family proteins, it is the first mRNA demethylase that has been identified.[5] Certain alleles of the FTO gene appear to be correlated with obesity in humans.[6][7]



Function
The amino acid sequence of the transcribed FTO protein shows high similarity with the enzyme AlkB which oxidatively demethylates DNA.[8][9] FTO is a member of the superfamily of alpha-ketoglutarate-dependent hydroxylase, which are non-heme iron-containing proteins. Recombinant FTO protein was first discovered to catalyze demethylation of 3-methylthymine in single-stranded DNA, and 3-methyluridine in single-stranded RNA, with low efficiency.[8] The nucleoside N6-methyladenosine, an abundant modification in RNA, was then found to be a major substrate of FTO.[5][10] The FTO gene expression was also found to be significantly upregulated in the hypothalamus of rats after food deprivation and strongly negatively correlated with the expression of orexigenic galanin-like peptide which is involved in the stimulation of food intake.[11]
Increases in hypothalamic expression of FTO are associated with the regulation of energy intake but not feeding reward.[12]
People with two copies of the risk allele for the rs9939609 single nucleotide polymorphism (SNP) showed differing neural responses to food images via fMRI.[13] However, rs9939609's association with FTO is controversial, and may actually affect another gene, called Iroquois homeobox protein 3 (IRX3).[14]
FTO demethylates RNA
N6-methyladenosine (m6A) is an abundant modification in mRNA and is found within some viruses,[15][16] and most eukaryotes including mammals,[17][18][19][20] insects,[21] plants,[22][23][24] and yeast.[25][26] It is also found in tRNA, rRNA, and small nuclear RNA (snRNA) as well as several long non-coding RNA, such as Xist.[10][27] Adenosine methylation is directed by a large m6A methyltransferase complex containing METTL3 as the SAM-binding sub-unit.[28] In vitro, this methyltransferase complex preferentially methylates RNA oligonucleotides containing GGACU[29] and a similar preference was identified in vivo in mapped m6A sites in Rous sarcoma virus genomic RNA[30] and in bovine prolactin mRNA.[31] In plants, the majority of the m6A is found within 150 nucleotides before the start of the poly(A) tail.[32]
Mapping of m6A in human and mouse RNA has identified over 18,000 m6A sites in the transcripts of more than 7,000 human genes with a consensus sequence of [G/A/U][G>A]m6AC[U>A/C][10][27] consistent with the previously identified motif.[29] Sites preferentially appear in two distinct landmarks—around stop codons and within long internal exons—and are highly conserved between human and mouse.[10][27] A subset of stimulus-dependent, dynamically modulated sites has been identified. Silencing the m6A methyltransferase significantly affects gene expression and alternative RNA splicing patterns, resulting in modulation of the p53 (also known as TP53) signalling pathway and apoptosis.
FTO has been demonstrated to efficiently demethylate the related modified ribonucleotide, N6,2'-O-dimethyladenosine, and to an equal or lesser extent, m6A, in vitro .[5][33] FTO knockdown with siRNA led to increased amounts of m6A in polyA-RNA, whereas overexpression of FTO resulted in decreased amounts of m6A in human cells.[10] FTO partially co-localizes with nuclear speckles, which supports the notion that in the nucleus, m6A can be a substrate of FTO. Function of FTO could affect the processing of pre-mRNA, other nuclear RNAs, or both. The discovery of the FTO-mediated oxidative demethylation of RNA may initiate further investigations on biological regulation based on reversible chemical modification of RNA, and identification of RNA substrates for which FTO has the highest affinity.[5][10][33]
FTO can oxidize m6A to generate N6 -hydroxymethyladenosine(hm6A) as an intermediate modification and N6 - formyladenosine(f6A) as a further oxidized product in mammalian cells.[34]
Tissue distribution
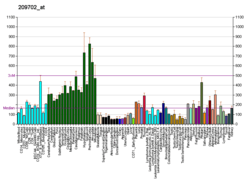
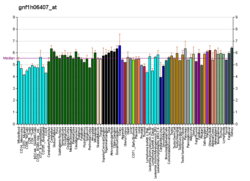
The FTO gene is widely expressed in both fetal and adult tissues.[35]
Clinical significance
Obesity
_Protein.png)
38,759 Europeans were studied for variants of FTO obesity risk allele.[35] In particular, carriers of one copy of the allele weighed on average 1.2 kilograms (2.6 lb) more than people with no copies. Carriers of two copies (16% of the subjects) weighed 3 kilograms (6.6 lb) more and had a 1.67-fold higher rate of obesity than those with no copies. The association was observed in ages 7 and upwards. This gene is not directly associated with diabetes; however, increased body-fat also increases the risk of developing type 2 diabetes.[36]
Simultaneously, a study in 2,900 affected individuals and 5,100 controls of French descent, together with 500 trios (confirming an association independent of population stratification) found association of SNPs in the very same region of FTO (rs1421085).[37] The authors found that this variation, or a variation in strong LD with this variation explains 1% of the population BMI variance and 22% of the population attributable risk of obesity. The authors of this study claim that while obesity was already known to have a genetic component (from twin studies), no replicated previous study has ever identified an obesity risk allele that was so common in the human population. The risk allele is a cluster of 10 single nucleotide polymorphism in the first intron of FTO called rs9939609. According to HapMap, it has population frequencies of 45% in the West/Central Europeans, 52% in Yorubans (West African natives) and 14% in Chinese/Japanese. Furthermore, morbid obesity is associated with a combination of FTO and INSIG2 single nucleotide polymorphisms.[38]
In 2009, variants in the FTO gene were further confirmed to associate with obesity in two very large genome wide association studies of body mass index (BMI).[39][40]
In adult humans, it was shown that adults bearing the at risk AT and AA alleles at rs9939609 consumed between 500 and 1250 kJ more each day than those carrying the protective TT genotype (equivalent to between 125 and 280 kcal per day more intake).[41] The same study showed that there was no impact of the polymorphism on energy expenditure. This finding of an effect of the rs9939609 polymorphism on food intake or satiety has been independently replicated in five subsequent studies (in order of publication).[42][43][44][45][46] Three of these subsequent studies also measured resting energy expenditure and confirmed the original finding that there is no impact of the polymorphic variation at the rs9939609 locus on energy expenditure. A different study explored the effects of variation in two different SNPs in the FTO gene (rs17817449 and rs1421085) and suggested there might be an effect on circulating leptin levels and energy expenditure, but this latter effect disappeared when the expenditure was normalised for differences in body composition.[47] The accumulated data across seven independent studies therefore clearly implicates the FTO gene in humans as having a direct impact on food intake but no effect on energy expenditure.
The obesity-associated noncoding region within the FTO gene interacts directly with the promoter of IRX3, a homeobox gene, and IRX5, another homeobox gene. The noncoding region of FTO interacts with the promoters of IRX3 and FTO in human, mouse and zebrafish, and with IRX5. Results suggest that IRX3 and IRX5 are linked with obesity and determine body mass and composition. This is further supported by the fact that obesity-associated single nucleotide polymorphisms, in which cytosine is substituted for thymine, are involved in the expression of IRX3 and IRX5 (not FTO) in human brains. The enhanced expression of IRX3 and IRX5 resulting from this single nucleotide alteration promoted a shift from energy-dissipating beige adipocytes to energy-storing white adipocytes and a subsequent reduction in mitochondrial thermogenesis by a factor of 5.[48][49] Another study found indications that the FTO allele associated with obesity represses mitochondrial thermogenesis in adipocyte precursor cells in a tissue-autonomous manner, and that there is a pathway for adipocyte thermoregulation which involves the proteine ARID5B, the single-nucleotide variant rs1421085, and the IRX3 and IRX5 genes.[50]
Alzheimer's disease
Recent studies revealed that carriers of common FTO gene polymorphisms show both a reduction in frontal lobe volume of the brain[51] and an impaired verbal fluency performance.[52] Fittingly, a population-based study from Sweden found that carriers of the FTO rs9939609 A allele have an increased risk for incident Alzheimer disease.[53]
Other diseases
The presence of the FTO rs9939609 A allele was also found to be positively correlated with other symptoms of the metabolic syndrome, including higher fasting insulin, glucose, and triglycerides, and lower HDL cholesterol. However all these effects appear to be secondary to weight increase since no association was found after correcting for increases in body mass index.[54] Similarly, the association of rs11076008 G allele with the increased risk for degenerative disc disease was reported.[55]
Model organisms
Model organisms have been used in the study of FTO function. In contrast to the findings in humans deletion, analysis of the Fto gene in mice showed loss of function is associated with no differences in energy intake but greater energy expenditure and this results in a reduction of body weight and fatness.[56]
| Characteristic | Phenotype |
|---|---|
| Homozygote viability | Normal |
| Fertility | Normal |
| Body weight | Normal |
| Anxiety | Normal |
| Neurological assessment | Normal |
| Grip strength | Normal |
| Hot plate | Normal |
| Dysmorphology | Normal |
| Indirect calorimetry | Normal |
| Glucose tolerance test | Normal |
| Auditory brainstem response | Normal |
| DEXA | Normal |
| Radiography | Abnormal[57] |
| Body temperature | Normal |
| Eye morphology | Normal |
| Clinical chemistry | Normal |
| Plasma immunoglobulins | Normal |
| Haematology | Normal |
| Micronucleus test | Normal |
| Heart weight | Normal |
| Skin Histopathology | Normal |
| Brain histopathology | Normal |
| Eye Histopathology | Normal |
| Salmonella infection | Normal[58] |
| Citrobacter infection | Normal[59] |
| All tests and analysis from[60][61] |
Another conditional knockout mouse line, called Ftotm1a(EUCOMM)Wtsi[62][63] was generated as part of the International Knockout Mouse Consortium program — a high-throughput mutagenesis project to generate and distribute animal models of disease to interested scientists.[64][65][66] Male and female animals from this line underwent a standardized phenotypic screen to determine the effects of deletion.[60][67] Twenty five tests were carried out on mutant mice and only significant skeletal abnormalities were observed, including kyphosis and abnormal vertebral transverse processes, and only in female homozygous mutant animals.[60]
The reasons for the differences in FTO phenotype between humans and different lines of mice is presently uncertain. However, many other genes involved in regulation of energy balance exert effects on both intake and expenditure.
Origin of name
By exon trapping, Peters et al. (1999) cloned a novel gene from a region of several hundred kb deleted by the mouse 'fused toes' (FT) mutation. They named the gene 'fatso' (Fto) due to its large size.[68][69]
References
- GRCh38: Ensembl release 89: ENSG00000140718 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000055932 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Jia G, Fu Y, Zhao X, Dai Q, Zheng G, Yang Y, Yi C, Lindahl T, Pan T, Yang YG, He C (October 2011). "N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO". Nature Chemical Biology. 7 (12): 885–7. doi:10.1038/nchembio.687. PMC 3218240. PMID 22002720.
- Loos RJ, Yeo GS (January 2014). "The bigger picture of FTO: the first GWAS-identified obesity gene". Nature Reviews. Endocrinology. 10 (1): 51–61. doi:10.1038/nrendo.2013.227. PMC 4188449. PMID 24247219.
- Claussnitzer M, Dankel SN, Kim KH, Quon G, Meuleman W, Haugen C, et al. (September 2015). "FTO Obesity Variant Circuitry and Adipocyte Browning in Humans". The New England Journal of Medicine. 373 (10): 895–907. doi:10.1056/NEJMoa1502214. PMC 4959911. PMID 26287746.
- Gerken T, Girard CA, Tung YC, Webby CJ, Saudek V, Hewitson KS, Yeo GS, McDonough MA, Cunliffe S, McNeill LA, Galvanovskis J, Rorsman P, Robins P, Prieur X, Coll AP, Ma M, Jovanovic Z, Farooqi IS, Sedgwick B, Barroso I, Lindahl T, Ponting CP, Ashcroft FM, O'Rahilly S, Schofield CJ (November 2007). "The obesity-associated FTO gene encodes a 2-oxoglutarate-dependent nucleic acid demethylase". Science. 318 (5855): 1469–72. Bibcode:2007Sci...318.1469G. doi:10.1126/science.1151710. PMC 2668859. PMID 17991826.
- Sanchez-Pulido L, Andrade-Navarro MA (November 2007). "The FTO (fat mass and obesity associated) gene codes for a novel member of the non-heme dioxygenase superfamily". BMC Biochemistry. 8: 23. doi:10.1186/1471-2091-8-23. PMC 2241624. PMID 17996046.
- Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, Jaffrey SR (June 2012). "Comprehensive analysis of mRNA methylation reveals enrichment in 3' UTRs and near stop codons". Cell. 149 (7): 1635–46. doi:10.1016/j.cell.2012.05.003. PMC 3383396. PMID 22608085.
- Fredriksson R, Hägglund M, Olszewski PK, Stephansson O, Jacobsson JA, Olszewska AM, Levine AS, Lindblom J, Schiöth HB (May 2008). "The obesity gene, FTO, is of ancient origin, up-regulated during food deprivation and expressed in neurons of feeding-related nuclei of the brain". Endocrinology. 149 (5): 2062–71. doi:10.1210/en.2007-1457. PMID 18218688.
- Olszewski PK, Fredriksson R, Olszewska AM, Stephansson O, Alsiö J, Radomska KJ, Levine AS, Schiöth HB (October 2009). "Hypothalamic FTO is associated with the regulation of energy intake not feeding reward". BMC Neuroscience. 10: 129. doi:10.1186/1471-2202-10-129. PMC 2774323. PMID 19860904.
- Wiemerslage L, Nilsson EK, Solstrand Dahlberg L, Ence-Eriksson F, Castillo S, Larsen AL, Bylund SB, Hogenkamp PS, Olivo G, Bandstein M, Titova OE, Larsson EM, Benedict C, Brooks SJ, Schiöth HB (May 2016). "An obesity-associated risk allele within the FTO gene affects human brain activity for areas important for emotion, impulse control and reward in response to food images" (PDF). The European Journal of Neuroscience. 43 (9): 1173–80. doi:10.1111/ejn.13177. PMID 26797854.
- Rask-Andersen M, Almén MS, Schiöth HB (November 2015). "Scrutinizing the FTO locus: compelling evidence for a complex, long-range regulatory context". Human Genetics. 134 (11–12): 1183–93. doi:10.1007/s00439-015-1599-5. PMID 26340902.
- Aloni Y, Dhar R, Khoury G (October 1979). "Methylation of nuclear simian virus 40 RNAs". Journal of Virology. 32 (1): 52–60. doi:10.1128/JVI.32.1.52-60.1979. PMC 353526. PMID 232187.
- Beemon K, Keith J (June 1977). "Localization of N6-methyladenosine in the Rous sarcoma virus genome". Journal of Molecular Biology. 113 (1): 165–79. doi:10.1016/0022-2836(77)90047-X. PMID 196091.
- Desrosiers R, Friderici K, Rottman F (October 1974). "Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells". Proceedings of the National Academy of Sciences of the United States of America. 71 (10): 3971–5. Bibcode:1974PNAS...71.3971D. doi:10.1073/pnas.71.10.3971. PMC 434308. PMID 4372599.
- Adams JM, Cory S (May 1975). "Modified nucleosides and bizarre 5'-termini in mouse myeloma mRNA". Nature. 255 (5503): 28–33. Bibcode:1975Natur.255...28A. doi:10.1038/255028a0. PMID 1128665.
- Wei CM, Gershowitz A, Moss B (January 1976). "5'-Terminal and internal methylated nucleotide sequences in HeLa cell mRNA". Biochemistry. 15 (2): 397–401. doi:10.1021/bi00647a024. PMID 174715.
- Perry RP, Kelley DE, Friderici K, Rottman F (April 1975). "The methylated constituents of L cell messenger RNA: evidence for an unusual cluster at the 5' terminus". Cell. 4 (4): 387–94. doi:10.1016/0092-8674(75)90159-2. PMID 1168101.
- Levis R, Penman S (April 1978). "5'-terminal structures of poly(A)+ cytoplasmic messenger RNA and of poly(A)+ and poly(A)- heterogeneous nuclear RNA of cells of the dipteran Drosophila melanogaster". Journal of Molecular Biology. 120 (4): 487–515. doi:10.1016/0022-2836(78)90350-9. PMID 418182.
- Nichols JL (August 1979). "N6-methyladenosine in maize poly(A)-containing RNA". Plant Science Letters. 15 (4): 357–361. doi:10.1016/0304-4211(79)90141-X.
- Kennedy TD, Lane BG (June 1979). "Wheat embryo ribonucleates. XIII. Methyl-substituted nucleoside constituents and 5'-terminal dinucleotide sequences in bulk poly(AR)-rich RNA from imbibing wheat embryos". Canadian Journal of Biochemistry. 57 (6): 927–31. doi:10.1139/o79-112. PMID 476526.
- Zhong S, Li H, Bodi Z, Button J, Vespa L, Herzog M, Fray RG (May 2008). "MTA is an Arabidopsis messenger RNA adenosine methylase and interacts with a homolog of a sex-specific splicing factor". The Plant Cell. 20 (5): 1278–88. doi:10.1105/tpc.108.058883. PMC 2438467. PMID 18505803.
- Clancy MJ, Shambaugh ME, Timpte CS, Bokar JA (October 2002). "Induction of sporulation in Saccharomyces cerevisiae leads to the formation of N6-methyladenosine in mRNA: a potential mechanism for the activity of the IME4 gene". Nucleic Acids Research. 30 (20): 4509–18. doi:10.1093/nar/gkf573. PMC 137137. PMID 12384598.
- Bodi Z, Button JD, Grierson D, Fray RG (September 2010). "Yeast targets for mRNA methylation". Nucleic Acids Research. 38 (16): 5327–35. doi:10.1093/nar/gkq266. PMC 2938207. PMID 20421205.
- Dominissini D, Moshitch-Moshkovitz S, Schwartz S, Salmon-Divon M, Ungar L, Osenberg S, Cesarkas K, Jacob-Hirsch J, Amariglio N, Kupiec M, Sorek R, Rechavi G (April 2012). "Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq". Nature. 485 (7397): 201–6. Bibcode:2012Natur.485..201D. doi:10.1038/nature11112. PMID 22575960.
- Bokar JA, Shambaugh ME, Polayes D, Matera AG, Rottman FM (November 1997). "Purification and cDNA cloning of the AdoMet-binding subunit of the human mRNA (N6-adenosine)-methyltransferase". RNA. 3 (11): 1233–47. PMC 1369564. PMID 9409616.
- Harper JE, Miceli SM, Roberts RJ, Manley JL (October 1990). "Sequence specificity of the human mRNA N6-adenosine methylase in vitro". Nucleic Acids Research. 18 (19): 5735–41. doi:10.1093/nar/18.19.5735. PMC 332308. PMID 2216767.
- Kane SE, Beemon K (September 1985). "Precise localization of m6A in Rous sarcoma virus RNA reveals clustering of methylation sites: implications for RNA processing". Molecular and Cellular Biology. 5 (9): 2298–306. doi:10.1128/mcb.5.9.2298. PMC 366956. PMID 3016525.
- Horowitz S, Horowitz A, Nilsen TW, Munns TW, Rottman FM (September 1984). "Mapping of N6-methyladenosine residues in bovine prolactin mRNA". Proceedings of the National Academy of Sciences of the United States of America. 81 (18): 5667–71. Bibcode:1984PNAS...81.5667H. doi:10.1073/pnas.81.18.5667. PMC 391771. PMID 6592581.
- Bodi Z, Zhong S, Mehra S, Song J, Graham N, Li H, May S, Fray RG (2012). "Adenosine Methylation in Arabidopsis mRNA is Associated with the 3' End and Reduced Levels Cause Developmental Defects". Frontiers in Plant Science. 3: 48. doi:10.3389/fpls.2012.00048. PMC 3355605. PMID 22639649.
- Mauer J, Luo X, Blanjoie A, Jiao X, Grozhik AV, Patil DP, Linder B, Pickering BF, Vasseur JJ, Chen Q, Gross SS, Elemento O, Debart F, Kiledjian M, Jaffrey SR (January 2017). "m in the 5' cap controls mRNA stability". Nature. 541 (7637): 371–375. Bibcode:2017Natur.541..371M. doi:10.1038/nature21022. PMC 5513158. PMID 28002401.
- Fu Y, Jia G, Pang X, Wang RN, Wang X, Li CJ, et al. (2013). "FTO-mediated formation of N6-hydroxymethyladenosine and N6-formyladenosine in mammalian RNA". Nature Communications. 4: 1798. doi:10.1038/ncomms2822. PMC 3658177. PMID 23653210.
- Frayling TM, Timpson NJ, Weedon MN, Zeggini E, Freathy RM, Lindgren CM, et al. (May 2007). "A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity". Science. 316 (5826): 889–94. Bibcode:2007Sci...316..889F. doi:10.1126/science.1141634. PMC 2646098. PMID 17434869.
- Sandholt CH, Hansen T, Pedersen O (July 2012). "Beyond the fourth wave of genome-wide obesity association studies". Nutrition & Diabetes. 2 (7): e37. doi:10.1038/nutd.2012.9. PMC 3408643. PMID 23168490.
- Dina C, Meyre D, Gallina S, Durand E, Körner A, Jacobson P, Carlsson LM, Kiess W, Vatin V, Lecoeur C, Delplanque J, Vaillant E, Pattou F, Ruiz J, Weill J, Levy-Marchal C, Horber F, Potoczna N, Hercberg S, Le Stunff C, Bougnères P, Kovacs P, Marre M, Balkau B, Cauchi S, Chèvre JC, Froguel P (June 2007). "Variation in FTO contributes to childhood obesity and severe adult obesity". Nature Genetics. 39 (6): 724–6. doi:10.1038/ng2048. PMID 17496892.
- Chu X, Erdman R, Susek M, Gerst H, Derr K, Al-Agha M, Wood GC, Hartman C, Yeager S, Blosky MA, Krum W, Stewart WF, Carey D, Benotti P, Still CD, Gerhard GS (March 2008). "Association of morbid obesity with FTO and INSIG2 allelic variants". Archives of Surgery. 143 (3): 235–40, discussion 241. doi:10.1001/archsurg.2007.77. PMID 18347269.
- Thorleifsson G, Walters GB, Gudbjartsson DF, Steinthorsdottir V, Sulem P, Helgadottir A, Styrkarsdottir U, Gretarsdottir S, Thorlacius S, Jonsdottir I, Jonsdottir T, Olafsdottir EJ, Olafsdottir GH, Jonsson T, Jonsson F, Borch-Johnsen K, Hansen T, Andersen G, Jorgensen T, Lauritzen T, Aben KK, Verbeek AL, Roeleveld N, Kampman E, Yanek LR, Becker LC, Tryggvadottir L, Rafnar T, Becker DM, Gulcher J, Kiemeney LA, Pedersen O, Kong A, Thorsteinsdottir U, Stefansson K (January 2009). "Genome-wide association yields new sequence variants at seven loci that associate with measures of obesity". Nature Genetics. 41 (1): 18–24. doi:10.1038/ng.274. PMID 19079260.
- Willer CJ, Speliotes EK, Loos RJ, Li S, Lindgren CM, Heid IM, et al. (January 2009). "Six new loci associated with body mass index highlight a neuronal influence on body weight regulation". Nature Genetics. 41 (1): 25–34. doi:10.1038/ng.287. PMC 2695662. PMID 19079261.
- Speakman JR, Rance KA, Johnstone AM (August 2008). "Polymorphisms of the FTO gene are associated with variation in energy intake, but not energy expenditure". Obesity. 16 (8): 1961–5. doi:10.1038/oby.2008.318. PMID 18551109.
- Wardle J, Carnell S, Haworth CM, Farooqi IS, O'Rahilly S, Plomin R (September 2008). "Obesity associated genetic variation in FTO is associated with diminished satiety". The Journal of Clinical Endocrinology and Metabolism. 93 (9): 3640–3. doi:10.1210/jc.2008-0472. PMID 18583465.
- Timpson NJ, Emmett PM, Frayling TM, Rogers I, Hattersley AT, McCarthy MI, Davey Smith G (October 2008). "The fat mass- and obesity-associated locus and dietary intake in children". The American Journal of Clinical Nutrition. 88 (4): 971–8. doi:10.1093/ajcn/88.4.971. PMC 4773885. PMID 18842783.
- Haupt A, Thamer C, Staiger H, Tschritter O, Kirchhoff K, Machicao F, Häring HU, Stefan N, Fritsche A (April 2009). "Variation in the FTO gene influences food intake but not energy expenditure". Experimental and Clinical Endocrinology & Diabetes. 117 (4): 194–7. doi:10.1055/s-0028-1087176. PMID 19053021.
- Wardle J, Llewellyn C, Sanderson S, Plomin R (January 2009). "The FTO gene and measured food intake in children". International Journal of Obesity. 33 (1): 42–5. doi:10.1038/ijo.2008.174. PMID 18838977.
- Cecil JE, Tavendale R, Watt P, Hetherington MM, Palmer CN (December 2008). "An obesity-associated FTO gene variant and increased energy intake in children". The New England Journal of Medicine. 359 (24): 2558–66. doi:10.1056/NEJMoa0803839. PMID 19073975.
- Do R, Bailey SD, Desbiens K, Belisle A, Montpetit A, Bouchard C, Pérusse L, Vohl MC, Engert JC (April 2008). "Genetic variants of FTO influence adiposity, insulin sensitivity, leptin levels, and resting metabolic rate in the Quebec Family Study". Diabetes. 57 (4): 1147–50. doi:10.2337/db07-1267. PMID 18316358.
- Smemo S, Tena JJ, Kim KH, Gamazon ER, Sakabe NJ, Gómez-Marín C, Aneas I, Credidio FL, Sobreira DR, Wasserman NF, Lee JH, Puviindran V, Tam D, Shen M, Son JE, Vakili NA, Sung HK, Naranjo S, Acemel RD, Manzanares M, Nagy A, Cox NJ, Hui CC, Gomez-Skarmeta JL, Nóbrega MA (March 2014). "Obesity-associated variants within FTO form long-range functional connections with IRX3". Nature. 507 (7492): 371–5. Bibcode:2014Natur.507..371S. doi:10.1038/nature13138. PMC 4113484. PMID 24646999.
- Harrison P (19 August 2015). "Obesity Genetics: New Insights Might Mean New Therapies". Medscape.
- Claussnitzer M, Dankel SN, Kim KH, Quon G, Meuleman W, Haugen C, Glunk V, Sousa IS, Beaudry JL, Puviindran V, Abdennur NA, Liu J, Svensson PA, Hsu YH, Drucker DJ, Mellgren G, Hui CC, Hauner H, Kellis M (September 2015). "FTO Obesity Variant Circuitry and Adipocyte Browning in Humans". The New England Journal of Medicine. 373 (10): 895–907. doi:10.1056/NEJMoa1502214. PMC 4959911. PMID 26287746.
- Ho AJ, Stein JL, Hua X, Lee S, Hibar DP, Leow AD, et al. (May 2010). "A commonly carried allele of the obesity-related FTO gene is associated with reduced brain volume in the healthy elderly". Proceedings of the National Academy of Sciences of the United States of America. 107 (18): 8404–9. Bibcode:2010PNAS..107.8404H. doi:10.1073/pnas.0910878107. PMC 2889537. PMID 20404173.
- Benedict C, Jacobsson JA, Rönnemaa E, Sällman-Almén M, Brooks S, Schultes B, Fredriksson R, Lannfelt L, Kilander L, Schiöth HB (June 2011). "The fat mass and obesity gene is linked to reduced verbal fluency in overweight and obese elderly men". Neurobiology of Aging. 32 (6): 1159.e1–5. doi:10.1016/j.neurobiolaging.2011.02.006. PMID 21458110.
- Keller L, Xu W, Wang HX, Winblad B, Fratiglioni L, Graff C (2011). "The obesity related gene, FTO, interacts with APOE, and is associated with Alzheimer's disease risk: a prospective cohort study". Journal of Alzheimer's Disease. 23 (3): 461–9. doi:10.3233/JAD-2010-101068. PMID 21098976.
- Freathy RM, Timpson NJ, Lawlor DA, Pouta A, Ben-Shlomo Y, Ruokonen A, Ebrahim S, Shields B, Zeggini E, Weedon MN, Lindgren CM, Lango H, Melzer D, Ferrucci L, Paolisso G, Neville MJ, Karpe F, Palmer CN, Morris AD, Elliott P, Jarvelin MR, Smith GD, McCarthy MI, Hattersley AT, Frayling TM (May 2008). "Common variation in the FTO gene alters diabetes-related metabolic traits to the extent expected given its effect on BMI". Diabetes. 57 (5): 1419–26. doi:10.2337/db07-1466. PMC 3073395. PMID 18346983.
- Lao L, Zhong G, Li X, Liu Z (February 2014). "A preliminary association study of fat mass and obesity associated gene polymorphisms and degenerative disc disease in a Chinese Han population". The Journal of International Medical Research. 42 (1): 205–12. doi:10.1177/0300060513503761. PMID 24304927.
- Fischer J, Koch L, Emmerling C, Vierkotten J, Peters T, Brüning JC, Rüther U (April 2009). "Inactivation of the Fto gene protects from obesity". Nature. 458 (7240): 894–8. Bibcode:2009Natur.458..894F. doi:10.1038/nature07848. PMID 19234441.
- "Radiography data for Fto". Wellcome Trust Sanger Institute.
- "Salmonella infection data for Fto". Wellcome Trust Sanger Institute.
- "Citrobacter infection data for Fto". Wellcome Trust Sanger Institute.
- Gerdin AK (2010). "The Sanger Mouse Genetics Programme: High throughput characterisation of knockout mice". Acta Ophthalmologica. 88: 925–7. doi:10.1111/j.1755-3768.2010.4142.x.
- Mouse Resources Portal, Wellcome Trust Sanger Institute.
- "International Knockout Mouse Consortium".
- "Mouse Genome Informatics".
- Skarnes WC, Rosen B, West AP, Koutsourakis M, Bushell W, Iyer V, Mujica AO, Thomas M, Harrow J, Cox T, Jackson D, Severin J, Biggs P, Fu J, Nefedov M, de Jong PJ, Stewart AF, Bradley A (June 2011). "A conditional knockout resource for the genome-wide study of mouse gene function". Nature. 474 (7351): 337–42. doi:10.1038/nature10163. PMC 3572410. PMID 21677750.
- Dolgin E (June 2011). "Mouse library set to be knockout". Nature. 474 (7351): 262–3. doi:10.1038/474262a. PMID 21677718.
- Collins FS, Rossant J, Wurst W (January 2007). "A mouse for all reasons". Cell. 128 (1): 9–13. doi:10.1016/j.cell.2006.12.018. PMID 17218247.
- van der Weyden L, White JK, Adams DJ, Logan DW (June 2011). "The mouse genetics toolkit: revealing function and mechanism". Genome Biology. 12 (6): 224. doi:10.1186/gb-2011-12-6-224. PMC 3218837. PMID 21722353.
- Peters T, Ausmeier K, Rüther U (October 1999). "Cloning of Fatso (Fto), a novel gene deleted by the Fused toes (Ft) mouse mutation". Mammalian Genome. 10 (10): 983–6. doi:10.1007/s003359901144. PMID 10501967.
- Kim B, Kim Y, Cooke PS, Rüther U, Jorgensen JS (May 2011). "The fused toes locus is essential for somatic-germ cell interactions that foster germ cell maturation in developing gonads in mice". Biology of Reproduction. 84 (5): 1024–32. doi:10.1095/biolreprod.110.088559. PMID 21293032.
External links
- FTO protein, human at the US National Library of Medicine Medical Subject Headings (MeSH)
- Catharine Paddock (2007-04-13). "Obesity Gene Discovered". Medical News Today. Retrieved 2008-03-22.
- Overview of all the structural information available in the PDB for UniProt: Q9C0B1 (Alpha-ketoglutarate-dependent dioxygenase FTO) at the PDBe-KB.