Choristodera
Choristodera is an extinct order of semiaquatic diapsid reptiles that ranged from the Middle Jurassic, or possibly Late Triassic, to at least the early Miocene. It was named by Edward Drinker Cope in 1884.[1][2] Choristoderes have been found in North America, Asia, and Europe, and possibly also North Africa[3] and East Timor.[4] The most common fossils are typically found from the Late Cretaceous to the lower Eocene. Cladists have placed them between basal diapsids and basal archosauromorphs, but the phylogenetic position of Choristodera is still uncertain. It has also been proposed that they represent basal lepidosauromorphs. Most recently, workers have tentatively placed Choristodera within Archosauromorpha, with their ear bone anatomy more closely correlating to those of archosaurs and turtles than to those of lepidosaurs.[5]

| Choristodera | |
|---|---|
 | |
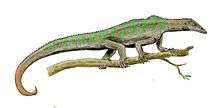
| Life restoration of a Hyphalosaurus | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Sauria |
| Order: | †Choristodera Cope, 1884 |
| Subgroups | |
| |
Description and phylogeny

Champsosauridae is the best-known family of the Choristodera and typifies the group. Champsosaurus was first described from Late Cretaceous strata of Montana by Cope in 1876. Champsosaurs resembled modern gharials (gavials) or false gharials. The skull of these animals have a long, thin snout filled with small, sharp conical teeth. This is due to champsosaurs and gharials occupying similar Ecological niches: Hunting small aquatic prey in rivers and swamps. This is a classic example of convergent evolution. More primitive choristoderes have shorter, broader snouts.
There are major differences seen between choristoderes and gharials and other crocodilians, however, particularly in the skull. The orbits are found well forward on the skull, and the rear of the skull is bulbous, hugely expanded and consists of complex bony arches surrounding empty space. These spaces probably contained massive jaw muscles. Other hypotheses for the spaces, such as an otic sensory organ housing, have been tossed around with little support. The external nares are found on the tip of the rostrum. This indicates that champsosaurs breathed while submerged by extending their rostrum through the water surface while their body rested on the bed of the lake or stream. Crocodylians and phytosaurs have their nares located dorsally on their rostrum or skull respectively. This position allows them to rest submerged just below the surface.
Champsosaur skulls are actually very similar to lizard skulls, though heavily modified. This has led some researchers to consider champsosaurids as lepidosauromorphs. However, champsosaurs lack the complex quadrate of lepidosaurians. With features of both diapsid groups, the phylogenetic position of Choristodera is highly confused.
Other features found in choristoderes include heavily ossified gastralia and modified distal limbs, not just the manus and pes, used as paddles. In addition, champsosaur ribs are short and massive, as in other aquatic reptiles. The thorax is dorso-ventrally flattened, and the tail is laterally compressed to aid in swimming. Skin impressions found with champsosaur fossils show non-overlapping scales of very small size and the skin containing no scutes like those found in crocodilians (see crocodile exoskeleton).
Most choristoderes have rather simple teeth, but neochoristoderes have teeth completely enveloped in striated enamel with an enamel infolding at the base, labiolingually compressed and hooked, the exception being Ikechosaurus which has still rather simple teeth aside from the start of an enamel infolding. There is some tooth differentiation, with the anterior teeth being larger than the posterior ones. Choristoderes have retained palatal teeth, indicating food manipulation in the mouth.[6]
As a whole, choristoderes were exceptionally well adapted to life in the water. In forms like Champsosaurus, only adult females could crawl ashore to lay eggs on land, with males and juveniles appearing to be incapable of doing so,[7] while Hyphalosaurus and Monjurosuchus are known to have been ovoviviparous.
It appears that, at least on Champsosaurus, the ear bones were internal and located near the base of the skull. This study also finds nasal conchae in all neochoristoderes aside from Ikechosaurus, suggesting this animals were capable of regulating their body temperature in their cold aquatic environments.[5]
Champsosaurid remains are known from as far as East Timor.[4]
Classification
Strict consensus of 10 most parsimonious trees from the analysis of Matsumoto et al. (2019):[8]
| order Choristodera |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Fossil record


Long considered to be a morphologically conservative group, recent phylogenetic analyses and descriptions of new taxa have revolutionized understanding of this taxon.
The order Choristodera comprises two monophyletic groups and three basal taxa. Primitive choristoderes are characterized by small body size, a large, dorsally directed orbit and closed lower temporal fenestrae.
Cteniogenys is the most basal choristodere. Like Monjurosuchus, it is a small-bodied, lizard-like animal. The webbed feet of Monjurosuchus reflect its aquatic lifestyle. Cteniogenys is known from North America and Europe, while Monjurosuchus is known from China.
Lazarussuchus was considered the most basal choristoderan when first described, and hence a late-surviving basal choristodere. However, Matsumoto et al. (2013) recovered Lazarussuchus in a clade along with hyphalosaurids and monjurosuchids, as did the cladistic analysis of Coeruleodraco by Matsumoto et al. (2019). This taxon is known from Cenozoic deposits in France and the Czech Republic. One species, L. dvoraki, persisted into the early Miocene in the Czech Republic.[9]
The two small-bodied choristoderes, Shokawa and Hyphalosaurus form the clade Hyphalosauridae within Choristodera. Their elongate necks and tails represent a unique shared derived condition. They resemble small plesiosaurs or nothosaurs. Shokawa is known from Japan, and Hyphalosaurus is known from China.
The named clade Neochoristodera includes Champsosaurus, Tchoiria, Simoedosaurus, and Ikechosaurus. This group of large-bodied reptiles is characterized by elongate snouts and relatively small orbits. Although it reflects our early understanding of the Choristodera as a whole, it in fact represents a highly derived condition.
The neochoristoderan taxa Champsosauridae and Simoedosauridae are found from the Late Cretaceous to the middle Eocene, indicating that the group survived the Cretaceous–Paleogene extinction event. Champsosaurus teeth, vertebrae, and other bones are common fossils in Cretaceous deposits such as the Dinosaur Park Formation of Alberta and the Lance Formation of Wyoming. The presence of champsosaurs as far north as the North Slope of Alaska implies warmer temperatures during the Cretaceous. The Paleocene species, Champsosaurus gigas, is the largest known champsosaur. The most complete skeleton was found at the Wannagan Creek site in North Dakota, but is found throughout North American Paleocene strata in the Dakotas, Wyoming, and Montana. C. gigas is unusual among Paleocene reptiles in that it is far larger than known Mesozoic ancestors, at about 3 meters in length (10 ft) versus 1.5 meters (5 ft) for the largest Cretaceous champsosaurs.
Simoedosaurus is known from Europe, western Asia and North America, Tchoria is known from Mongolia and Ikechosaurus is known from China. Champsosaurus is known from North America, where its distinctive, hourglass-shaped vertebral centra are important biostratigraphic indicators.
Pachystropheus rhaeticus may or may not be a choristodere. If a member of Choristodera, it extends the fossil record of the choristoderes from the Middle Jurassic back 45 mya and implies a significant ghost lineage. However, Pachystropheus lacks any cranial material, and all of the postcranial adaptations that link it to choristoderes may merely reflect convergent adaptations to an aquatic lifestyle. All unambiguous choristoderes were freshwater animals, while Pachystropheus was recovered from European marine transgressive sequences. Pachystropheus may be a choristodere, or it may be a type of thalattosaur, a large bodied, long necked group of marine reptiles. More complete fossil material is needed in order to resolve the placement of Pachystropheus.
The ultimate extinction of choristoderes may have been due to a number of possible causes, such as falling temperatures and habitat loss. Competition with crocodilians has occasionally been cited as a possible source of decline, but the co-existence of large choristoderes with aquatic crocodilians, as well as a fairly broad range of morphology in both clades, render this unresolved, if not unlikely.[10]
In 2007, Buffetaut et al. reported the discovery of a two-headed fossil of a juvenile Hyphalosaurus, apparently the first known case of a two-headed fossil tetrapod.[11]
Notes
- Cope, E.D. (1876). "On some extinct reptiles and Batrachia from the Judith River and Fox Hills beds of Montana". Proceedings of the Academy of Natural Sciences of Philadelphia. 1876: 340–359.
- Cope, E.D. (1884). "The Choristodera". American Naturalist. 18: 815–817.
- Haddoumi, Hamid; Allain, Ronan; Meslouh, Said; Metais, Grégoire; Monbaron, Michel; Pons, Denise; Rage, Jean-Claude; Vullo, Romain; Zouhri, Samir; Gheerbrant, Emmanuel (2016). "Guelb el Ahmar (Bathonian, Anoual Syncline, eastern Morocco): First continental flora and fauna including mammals from the Middle Jurassic of Africa" (PDF). Gondwana Research. 29 (1): 290–319. Bibcode:2016GondR..29..290H. doi:10.1016/j.gr.2014.12.004.
- J. H. F. Umbgrove, Structural History of the East Indies
- Dudgeon, Thomas William (2019). The internal cranial anatomy of Champsosaurus lindoei and its functional implications (PDF). Earth Sciences (M.Sci.). Ottawa, Ontario: Carleton University.
- Matsumoto, Ryoko; Evans, Susan E. (2016). "Morphology and function of the palatal dentition in Choristodera". Journal of Anatomy. 228 (3): 414–429. doi:10.1111/joa.12414. PMC 5341546. PMID 26573112.
- Katsura, Yoshihiro (2007). "Fusion of sacrals and anatomy in Champsosaurus(Diapsida, Choristodera)". Historical Biology. 19 (3): 263–271. doi:10.1080/08912960701374659.
- Matsumoto, Ryoko; Dong, Liping; Wang, Yuan; Evans, Susan E. (2019). "The first record of a nearly complete choristodere (Reptilia: Diapsida) from the Upper Jurassic of Hebei province, People's Republic of China". Journal of Systematic Palaeontology. 17 (12): 1031–1048. doi:10.1080/14772019.2018.1494220.
- Evans, Susan E.; Klembara, Jozef (2005). "A choristoderan reptile (Reptilia: Diapsida) from the Lower Miocene of northwest Bohemia (Czech Republic)". Journal of Vertebrate Paleontology. 25 (1): 171–184. doi:10.1671/0272-4634(2005)025[0171:ACRRDF]2.0.CO;2. ISSN 0272-4634.
- Matsumoto, R.; Evans, S.E. (2010). "Choristoderes and the freshwater assemblages of Laurasia". Journal of Iberian Geology. 36 (2): 253–274. doi:10.5209/rev_jige.2010.v36.n2.11.
- Buffetaut, Eric; Li, Jianjun; Tong, Haiyan; Zhang, He (2007). "A two-headed reptile from the Cretaceous of China". Biology Letters. 3 (1): 80–81. doi:10.1098/rsbl.2006.0580. PMC 2373827. PMID 17443971.
References
- de Braga, M.; Rieppel, O. (1997). "Reptile phylogeny and the interrelationships of turtles". Zoology – J. Linnean Soc. 120 (3): 281–354. doi:10.1111/j.1096-3642.1997.tb01280.x.
- Erickson, B.R. (1972). The Lepidosaurian Reptile Champsosaurus in North America. Paleontology, Monograph. 1. Science Museum of Minnesota.
- Evans, S.E.; Hecht, M.K. (1993). "A history of an extinct reptilian clade, the Choristodera: Longevity, Lazarus taxa, and the fossil record". Evolutionary Biology. 27: 323–338.
- Gao, K.; Fox, R.C. (1998). "New choristoderes (Reptilia: Diapsida) from the Upper Cretaceous and Palaeocene, Alberta and Saskatchewan, Canada, and phylogenetic relationships of Choristodera". Zoology – J. Linnean Soc. 124 (4): 303–353. doi:10.1111/j.1096-3642.1998.tb00580.x.
- Gao, K.-Q.; Ksepka, D.T. (2008). "Osteology and taxonomic revision of Hyphalosaurus (Diapsida: Choristodera) from the Lower Cretaceous of Liaoning, China". Journal of Anatomy. 212 (6): 747–760. doi:10.1111/j.1469-7580.2008.00907.x. PMC 2423398. PMID 18510504.
- Matsumoto, R.; Suzuki, S.; Tsogtbaatar, K.; Evans, S.E. (2009). "New material of the enigmatic reptile Khurendukhosaurus (Diapsida: Choristodera) from Mongolia". Naturwissenschaften. 96 (2): 233–242. Bibcode:2009NW.....96..233M. doi:10.1007/s00114-008-0469-6. PMID 19034405.
- Ksepka, D.; Gao, K.; Norell, M.A. (2005). "A new choristodere from the Cretaceous of Mongolia". American Museum Novitates. 3468: 1–22. doi:10.1206/0003-0082(2005)468<0001:ancftc>2.0.co;2. hdl:2246/2778.
- Storrs, G.W.; Gower, D.J. (1993). "The earliest possible choristodere (Diapsida) and gaps in the fossil record of semi-aquatic reptiles". Journal of the Geological Society [of America]. 150 (6): 1103–1107. Bibcode:1993JGSoc.150.1103S. doi:10.1144/gsjgs.150.6.1103.
External links
- "Choristodera page". Palæos. Archived from the original on 16 April 2007.
- "Choristodera-champsosaurs". Mikko's Phylogeny Archive.
- "Archosauria: Systematics". U.C. Berkeley.
- "Two-Headed Reptile Fossil Found". BBC News.
- "Ghost lineages". Archived from the original on 18 April 2008. — brief article on the fossil record of choristoderes



| |||||
| Basal choristoderes |  | ||||
| Hyphalosauridae | |||||
| Monjurosuchidae | |||||
| Neochoristodera |
| ||||
| Related categories |
| ||||