Vitamin K2
Vitamin K2 or menaquinone (/ˌmɛnəˈkwɪnoʊn/) is one of three types of vitamin K, the other two being vitamin K1 (phylloquinone) and K3 (menadione). K2 is both a tissue and bacterial product (derived from vitamin K1 in both cases) and is usually found in animal products or fermented foods.[1]
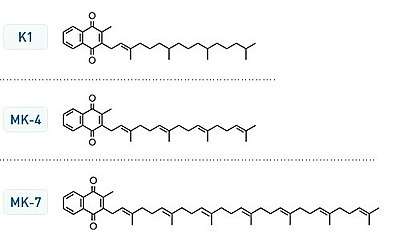
There are nine chemical variants of vitamin K2, determined by number of isoprenyl units in their side chains. The most common in the human diet is the short-chain, water-soluble menatetrenone (MK-4), which is usually produced by tissue and/or bacterial conversion of vitamin K1, and is commonly found in animal products. It is known that production of MK-4 from dietary plant vitamin K1 can be accomplished by animal tissues alone, as it proceeds in germ-free rodents.
Long-chain menaquinones (longer than MK-4) include MK-7, MK-8 and MK-9 and are more predominant in fermented foods such as natto. Longer-chain menaquinones (MK-10 to MK-13) are produced by anaerobic bacteria in the colon, but they are not well absorbed at this level and have little physiological impact.[1]
When there are no isoprenyl side chain units, the remaining molecule is vitamin K3. This can only be manufactured synthetically, and is used in animal feed. It was formerly given to premature infants, but due to inadvertent toxicity in the form of hemolytic anemia and jaundice, it is no longer used for this purpose.[1]
Description
Vitamin K2, the main storage form in animals, has several subtypes, which differ in isoprenoid chain length. These vitamin K2 homologues are called menaquinones, and are characterized by the number of isoprenoid residues in their side chains. Menaquinones are abbreviated MK-n, where M stands for menaquinone, the K stands for vitamin K, and the n represents the number of isoprenoid side chain residues. For example, menaquinone-4 (abbreviated MK-4) has four isoprene residues in its side chain. Menaquinone-4 (also known as menatetrenone from its four isoprene residues) is the most common type of vitamin K2 in animal products since MK-4 is normally synthesized from vitamin K1 in certain animal tissues (arterial walls, pancreas, and testes) by replacement of the phytyl tail with an unsaturated geranylgeranyl tail containing four isoprene units, thus yielding menaquinone-4 which is Water Soluble in nature. This homolog of vitamin K2 may have enzyme functions distinct from those of vitamin K1.
MK-7 and other long-chain menaquinones are different from MK-4 in that they are not produced by human tissue. MK-7 may be converted from phylloquinone (K1) in the colon by Escherichia coli bacteria.[2] However, these menaquinones synthesized by bacteria in the gut appear to contribute minimally to overall vitamin K status.[3][4] MK-4 and MK-7 are both found in the United States in dietary supplements for bone health.
The U.S. Food and Drug Administration (FDA) has not approved any form of vitamin K for the prevention or treatment of osteoporosis. MK-4 (45 mg daily) has been approved by the Ministry of Health in Japan since 1995 for the prevention and treatment of osteoporosis.[5]
All K vitamins are similar in structure: they share a "quinone" ring, but differ in the length and degree of saturation of the carbon tail and the number of repeating isoprene units in the "side chain".[6] The number of repeating units is indicated in the name of the particular menaquinone (e.g., MK-4 means that four isoprene units are repeated in the carbon tail). The chain length influences lipid solubility and thus transport to different target tissues.
Mechanism of action
The mechanism of action of vitamin K2 is similar to vitamin K1. K vitamins were first recognized as a factor required for coagulation, but the functions performed by this vitamin group were revealed to be much more complex. K vitamins play an essential role as cofactor for the enzyme γ-glutamyl carboxylase, which is involved in vitamin K-dependent carboxylation of the gla domain in "Gla proteins" (i.e., in conversion of peptide-bound glutamic acid (Glu) to γ-carboxy glutamic acid (Gla) in these proteins).
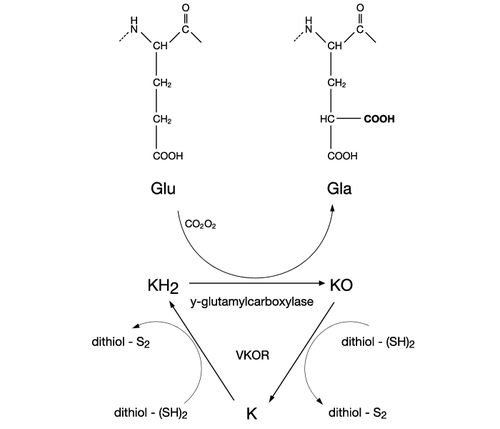
Carboxylation of these vitamin K-dependent Gla-proteins, besides being essential for the function of the protein, is also an important vitamin recovery mechanism since it serves as a recycling pathway to recover vitamin K from its epoxide metabolite (KO) for reuse in carboxylation.
Several human Gla-containing proteins synthesized in several different types of tissues have been discovered:
- Coagulation factors (II, VII, IX, X), as well as anticoagulation proteins (C, S, Z). These Gla-proteins are synthesized in the liver and play an important role in blood homeostasis.
- Osteocalcin. This non-collagenous protein is secreted by osteoblasts and plays an essential role in the formation of mineral in bone.
- Matrix gla protein (MGP). This calcification inhibitory protein is found in numerous body tissues, but its role is most pronounced in cartilage and in arterial vessel walls.
- Growth arrest-specific protein 6 (GAS6). GAS6 is secreted by leucocytes and endothelial cells in response to injury and helps in cell survival, proliferation, migration, and adhesion.
- Proline-rich Gla-proteins (PRGP), transmembrane Gla-proteins (TMG), Gla-rich protein (GRP) and periostin. Their precise functions are still unknown.
Health effects
MK-4 or MK-7 has a protective effect on bone mineral density and reduced risk of hip, vertebral and non-vertebral fractures.[7] These effects appear to be accentuated when combined with vitamin D and in the setting of osteoporosis.[1] The possible health benefits suggested for further investigation relate to bone strength and arterial health (reducing calcification or even decalcifying, with a possible reduction in blood pressure).[8]
Absorption profile
Vitamin K is absorbed along with dietary fat from the small intestine and transported by chylomicrons in the circulation. Most of vitamin K1 is carried by triacylglycerol-rich lipoproteins (TRL) and rapidly cleared by the liver; only a small amount is released into the circulation and carried by LDL and HDL. MK-4 is carried by the same lipoproteins (TRL, LDL, and HDL) and cleared fast as well. The long-chain menaquinones are absorbed in the same way as vitamin K1 and MK-4, but are efficiently redistributed by the liver in predominantly LDL (VLDL). Since LDL has a long half life in the circulation, these menaquinones can circulate for extended times resulting in higher bioavailability for extra-hepatic tissues as compared to vitamin K1 and MK-4. Accumulation of vitamin K in extra-hepatic tissues has direct relevance to vitamin K functions not related to hemostasis.[9]
Dietary intake in humans
The European Food Safety Authority (EU) and the US Institute of Medicine, on reviewing existing evidence, have decided there is insufficient evidence to publish a dietary reference value for vitamin K or for K2. They have, however, published an adequate intake (AI) for vitamin K, but no value specifically for K2. Evidence suggests K2 is converted from dietary K1 in mice, so they might not require a dietary intake of K2.[10]
Parts of the scientific literature, dating back to 1998, suggest that the AI values are based only on the hepatic requirements (i.e. related to the liver).[11][12] This hypothesis is supported by the fact that the majority of the Western population exhibits a substantial fraction of undercarboxylated extra-hepatic proteins. Thus, complete activation of coagulation factors is satisfied, but there does not seem to be enough vitamin K2 for the carboxylation of osteocalcin in bone and MGP in the vascular system.[13][14]
There is no known toxicity associated with high doses of menaquinones (vitamin K2). Unlike the other fat-soluble vitamins, vitamin K is not stored in any significant quantity in the liver; therefore the toxic level is not a described problem. All data available as of 2017 demonstrate that vitamin K has no adverse effects in healthy subjects. The recommendations for the daily intake of vitamin K, as issued recently by the US Institute of Medicine, also acknowledge the wide safety margin of vitamin K: "a search of the literature revealed no evidence of toxicity associated with the intake of either K1 or K2". Animal models involving rats, if generalisable to humans, show that MK-7 is well tolerated.[15]
Dietary sources
Apart from animal livers, the richest dietary source of menaquinones are fermented foods (from bacteria, not molds or yeasts); sources include cheeses consumed in Western diets (e.g., containing MK-8 and MK-9) and fermented soybean products (e.g., in traditional nattō consumed in Japan, containing MK-7). (Here and following it is noteworthy that most food assays measure only fully unsaturated menaquinones.)
MK-4 is synthesized by animal tissues and is found in meat, eggs, and dairy products.[16] Cheeses have been found to contain MK-8 at 10–20 μg per 100 g and MK-9 at 35–55 μg per 100 g.[9] In one report, no substantial differences in MK-4 levels were observed between wild game, free-range animals, and factory farm animals.[17]
In addition to its animal origins, menaquinones are synthesized by bacteria during fermentation and so, as stated, are found in most fermented cheese and soybean products.[18] As of 2001, the richest known source of natural K2 was nattō fermented using the nattō strain of Bacillus subtilis,[19] which is reportedly a good source of long-chain MK-7. In nattō, MK-4 is absent as a form of vitamin K, and in cheeses it is present among the vitamins K only in low proportions.[20] As of this date, it is unknown whether B. subtilis will produce K2 using other legumes (e.g., chickpeas, or lentils).
Food frequency questionnaire-derived estimates of relative intakes of Vitamins K in one northern European country suggest that for that population, about 90% of total vitamin K intakes are provided by K1, about 7.5% by MK-5 through MK-9 and about 2.5% by MK-4; the intense smell and strong taste of nattō appear to make this soya food a less attractive source of K2 for Western tastes.
With regard to utilisation, reports suggest that Vitamin K2 is preferred by the extrahepatic tissues (bone, cartilage, vasculature), which may be produced as MK-4 by the animal from K1, or it may be of bacterial origin (from MK-7, MK-9, and other MKs). Discussion is ongoing as to what extent K2 produced by human intestinal bacteria contributes to daily vitamin K2 needs.
Supplement companies sell nattō extract reportedly standardized with regards to K2 content, in capsule form.
| Food | Vitamin K2 (μg per 100 g) | Proportion of compounds |
|---|---|---|
| Nattō, cooked | 1,034.0[17] | 0% MK-4, 1% MK-5, 1% MK-6, 90% MK-7, 8% MK-8 |
| Goose liver pâté | 369.0[17] | 100% MK-4 |
| Australian emu oil | 360 | 100% MK-4 |
| Hard cheeses | 76.3[17] | 6% MK-4, 2% MK-5, 1% MK-6, 2% MK-7, 22% MK-8, 67% MK-9 |
| Soft cheeses | 56.5[17] | 6.5% MK-4, 0.5% MK-5, 1% MK-6, 2% MK-7, 20% MK-8, 70% MK-9 |
| Milk (4% fat, USA) |
38.1[21] | 2% MK-4, 46% MK-9, 7% MK-10, 45% MK-11 |
| Egg yolk (Netherlands) | 32.1[17] | 98% MK-4, 2% MK-6 |
| Goose leg | 31.0[17] | 100% MK-4 |
| Grass-fed ghee and butter oil | 19.6–43.1 average 29.9[22] |
|
| Curd cheeses | 24.8[17] | 2.6% MK-4, 0.4% MK-5, 1% MK-6, 1% MK-7, 20% MK-8, 75% MK-9 |
| Egg yolk (USA) | 15.5[23] | 100% MK-4 |
| Butter | 15.0[17] | 100% MK-4 |
| Chicken liver (raw) | 14.1[23] | 100% MK-4 |
| Chicken liver (pan-fried) | 12.6[23] | 100% MK-4 |
| Cheddar cheese (USA) | 10.2[23] | 6% MK-4, 94% other MK |
| Meat franks | 9.8[23] | 100% MK-4 |
| Salami | 9.0[17] | 100% MK-4 |
| Chicken breast | 8.9[17] | 100% MK-4 |
| Chicken leg | 8.5[17] | 100% MK-4 |
| Ground beef (medium fat) | 8.1[23] | 100% MK-4 |
| Luncheon meat | 7.7[17] | 100% MK-4 |
| Chicken liver (braised) | 6.7[23] | 100% MK-4 |
| Minced meat | 6.7[17] | 100% MK-4 |
| Calf's liver (pan-fried) | 6.0[23] | 100% MK-4 |
| Hot dog | 5.7[23] | 100% MK-4 |
| Bacon | 5.6[23] | 100% MK-4 |
| Whipping cream | 5.4[17] | 100% MK-4 |
| Sauerkraut | 4.8[17] | 8% MK-4, 17% MK-5, 31% MK-6, 4% MK-7, 17% MK-8, 23% MK-9 |
| Pork steak | 3.7[17] | 57% MK-4, 13% MK-7, 30% MK-8 |
| Duck breast | 3.6[17] | 100% MK-4 |
| Buttermilk | 2.5[17] | 8% MK-4, 4% MK-5, 4% MK-6, 4% MK-7, 24% MK-8, 56% MK-9 |
| Plaice | 2.2[17] | 9% MK-4, 14% MK-6, 4% MK-7, 73% MK-8 |
| Eel | 2.2[17] | 77% MK-4, 5% MK-6, 18% MK-7 |
| Fermented cod liver oil | 1.8[22] | 69% MK-4, 18% MK-6, 6% MK-8, 7% MK-9 |
| Chocolate | 1.5[17] | 100% MK-4 |
| Beef | 1.1[17] | 100% MK-4 |
| Buckwheat bread | 1.1[17] | 100% MK-7 |
| Whole milk yogurt | 0.9[17] | 67% MK-4, 11% MK-5, 22% MK-8 |
| Whole milk (Netherlands) |
0.9[17] | 89% MK-4, 11% MK-5 |
| Egg white | 0.9[17] | 100% MK-4 |
| Venison back | 0.7[17] | 100% MK-4 |
| Salmon | 0.5[17] | 100% MK-4 |
| Cow's liver (pan-fried) | 0.4[23] | 100% MK-4 |
| Mackerel | 0.4[17] | 100% MK-4 |
| Pork liver | 0.3[17] | 100% MK-4 |
| Rabbit leg | 0.1[17] | 100% MK-4 |
| Skimmed milk yogurt | 0.1[17] | 100% MK-8 |
Notes:

Prawns, herring, kale, spinach, broccoli, green peas, bananas, apples, oranges, margarine, corn oil, sunflower oil, olive oil, rye bread, wheat bread, sourdough bread, and tea contain vitamin K1 but not vitamin K2. Skimmed milk and coffee do not contain any vitamin K.[17]
Anticoagulants and vitamin K2 supplementation
Recent studies found a clear association between long-term oral (or intravenous) anticoagulant treatment (OAC) and reduced bone quality due to reduction of active osteocalcin. OAC might lead to an increased incidence of fractures, reduced bone mineral density or content, osteopenia, and increased serum levels of undercarboxylated osteocalcin.[24]
Furthermore, OAC is often linked to undesired soft-tissue calcification in both children and adults.[25][26] This process has been shown to be dependent upon the action of K vitamins. Vitamin K deficiency results in undercarboxylation of MGP. Vascular calcification was shown to appear in warfarin-treated experimental animals within two weeks.[27] Also in humans on OAC treatment, two-fold more arterial calcification was found as compared to patients not receiving vitamin K antagonists.[28][29] Among consequences of anticoagulant treatment: increased aortic wall stiffness, coronary insufficiency, ischemia, and even heart failure. Arterial calcification might also contribute to systolic hypertension and ventricular hypertrophy.[30][31] Coumarins, by interfering with vitamin K metabolism, are thought by some to lead to excessive calcification of cartilage and the tracheobronchial arteries.
Anticoagulant therapy is usually instituted to avoid life-threatening diseases, and high vitamin K intake interferes with anticoagulant effects. Patients on warfarin (Coumadin) or being treated with other vitamin K antagonists are therefore advised not to consume diets rich in K vitamins. However, the latest research proposes to combine vitamins K with OAC to stabilize the International normalized ratio (INR, a laboratory test measure of blood coagulation).
Individuals taking anticoagulant medications, such as warfarin (coumarins), should consult their doctor before taking vitamin K2.
In other organisms
Many bacteria synthesize menaquinones from chorismic acid. They use it as a part of the electron transport chain, playing a similar role as other quinones such as ubiquinone. Oxygen, heme, and menaquinones are needed for many species of lactic acid bacteria to conduct respiration.[32]
See also
References
- Myneni VD, Mezey E (November 2017). "Regulation of bone remodeling by vitamin K2". Oral Diseases. 23 (8): 1021–1028. doi:10.1111/odi.12624. PMC 5471136. PMID 27976475.
- Vermeer C, Braam L (2001). "Role of K vitamins in the regulation of tissue calcification". Journal of Bone and Mineral Metabolism. 19 (4): 201–6. doi:10.1007/s007740170021. PMID 11448011.
- Suttie JW (1995). "The importance of menaquinones in human nutrition". Annual Review of Nutrition. 15: 399–417. doi:10.1146/annurev.nu.15.070195.002151. PMID 8527227.
- Weber P (October 2001). "Vitamin K and bone health". Nutrition. 17 (10): 880–7. doi:10.1016/S0899-9007(01)00709-2. PMID 11684396.
- Iwamoto I, Kosha S, Noguchi S, Murakami M, Fujino T, Douchi T, Nagata Y (January 1999). "A longitudinal study of the effect of vitamin K2 on bone mineral density in postmenopausal women a comparative study with vitamin D3 and estrogen-progestin therapy". Maturitas. 31 (2): 161–4. doi:10.1016/S0378-5122(98)00114-5. PMID 10227010.
- Shearer, M. J. (2003). Physiology. Elsevier Sciences. pp. 6039–6045.
- Cockayne S, Adamson J, Lanham-New S, Shearer MJ, Gilbody S, Torgerson DJ (June 2006). "Vitamin K and the prevention of fractures: systematic review and meta-analysis of randomized controlled trials". Archives of Internal Medicine. 166 (12): 1256–61. doi:10.1001/archinte.166.12.1256. PMID 16801507.
- Jensen GS, Lenninger M, Ero MP, Benson KF (October 2016). "Consumption of nattokinase is associated with reduced blood pressure and von Willebrand factor, a cardiovascular risk marker: results from a randomized, double-blind, placebo-controlled, multicenter North American clinical trial". Integrated Blood Pressure Control. 9: 95–104. doi:10.2147/ibpc.s99553. PMC 5066864. PMID 27785095.
- Shearer MJ, Newman P (October 2008). "Metabolism and cell biology of vitamin K". Thrombosis and Haemostasis. 100 (4): 530–47. doi:10.1160/th08-03-0147. PMID 18841274.
- Okano T, Shimomura Y, Yamane M, Suhara Y, Kamao M, Sugiura M, Nakagawa K (April 2008). "Conversion of phylloquinone (Vitamin K1) into menaquinone-4 (Vitamin K2) in mice: two possible routes for menaquinone-4 accumulation in cerebra of mice". The Journal of Biological Chemistry. 283 (17): 11270–9. doi:10.1074/jbc.M702971200. PMID 18083713.
It is postulated that phylloquinone is converted into menaquinone-4 and accumulates in extrahepatic tissues. To clarify this, phylloquinone with a deuterium-labeled 2-methyl-1,4-naphthoquinone ring was given orally to mice, and cerebra were collected for D NMR and liquid chromatography-tandem mass spectrometry analyses.
- Booth SL, Suttie JW (May 1998). "Dietary intake and adequacy of vitamin K". The Journal of Nutrition. 128 (5): 785–8. doi:10.1093/jn/128.5.785. PMID 9566982.
- Schurgers LJ, Vermeer C (February 2002). "Differential lipoprotein transport pathways of K-vitamins in healthy subjects". Biochimica et Biophysica Acta (BBA) - General Subjects. 1570 (1): 27–32. doi:10.1016/s0304-4165(02)00147-2. PMID 11960685.
- Hofbauer LC, Brueck CC, Shanahan CM, Schoppet M, Dobnig H (March 2007). "Vascular calcification and osteoporosis--from clinical observation towards molecular understanding". Osteoporosis International. 18 (3): 251–9. doi:10.1007/s00198-006-0282-z. PMID 17151836.
- Plantalech L, Guillaumont M, Vergnaud P, Leclercq M, Delmas PD (November 1991). "Impairment of gamma carboxylation of circulating osteocalcin (bone gla protein) in elderly women". Journal of Bone and Mineral Research. 6 (11): 1211–6. doi:10.1002/jbmr.5650061111. PMID 1666807.
- Pucaj K, Rasmussen H, Møller M, Preston T (September 2011). "Safety and toxicological evaluation of a synthetic vitamin K2, menaquinone-7". Toxicology Mechanisms and Methods. 21 (7): 520–32. doi:10.3109/15376516.2011.568983. PMC 3172146. PMID 21781006.
- Elder SJ, Haytowitz DB, Howe J, Peterson JW, Booth SL (January 2006). "Vitamin K contents of meat, dairy, and fast food in the U.S. diet". Journal of Agricultural and Food Chemistry. 54 (2): 463–7. doi:10.1021/jf052400h. PMID 16417305.
- Schurgers LJ, Vermeer C (November 2000). "Determination of phylloquinone and menaquinones in food. Effect of food matrix on circulating vitamin K concentrations". Haemostasis. 30 (6): 298–307. doi:10.1159/000054147. PMID 11356998.
- Tsukamoto Y, Ichise H, Kakuda H, Yamaguchi M (2000). "Intake of fermented soybean (natto) increases circulating vitamin K2 (menaquinone-7) and gamma-carboxylated osteocalcin concentration in normal individuals". Journal of Bone and Mineral Metabolism. 18 (4): 216–22. doi:10.1007/s007740070023. PMID 10874601.
- Kaneki M, Hodges SJ, Hedges SJ, Hosoi T, Fujiwara S, Lyons A, et al. (April 2001). "Japanese fermented soybean food as the major determinant of the large geographic difference in circulating levels of vitamin K2: possible implications for hip-fracture risk". Nutrition. 17 (4): 315–21. doi:10.1016/s0899-9007(00)00554-2. PMID 11369171.
- "On the Trail of the Elusive X-Factor: Vitamin K2 Revealed".
- Fu X, Harshman SG, Shen X, Haytowitz DB, Karl JP, Wolfe BE, Booth SL (June 2017). "Multiple Vitamin K Forms Exist in Dairy Foods". Current Developments in Nutrition. 1 (6): e000638. doi:10.3945/cdn.117.000638. PMC 5998353. PMID 29955705.
- "Hook Line and Stinker" (PDF).
- Rhéaume-Bleue, Kate (August 27, 2013). Vitamin K2 and the Calcium Paradox: How a Little-Known Vitamin Could Save Your Life. Harper. pp. 66–67. ISBN 978-0062320049.
- Caraballo PJ, Gabriel SE, Castro MR, Atkinson EJ, Melton LJ (1999). "Changes in bone density after exposure to oral anticoagulants: a meta-analysis". Osteoporosis International. 9 (5): 441–8. doi:10.1007/s001980050169. PMID 10550464.
- Barnes C, Newall F, Ignjatovic V, Wong P, Cameron F, Jones G, Monagle P (April 2005). "Reduced bone density in children on long-term warfarin". Pediatric Research. 57 (4): 578–81. doi:10.1203/01.pdr.0000155943.07244.04. PMID 15695604.
- Hawkins D, Evans J (May 2005). "Minimising the risk of heparin-induced osteoporosis during pregnancy". Expert Opinion on Drug Safety. 4 (3): 583–90. doi:10.1517/14740338.4.3.583. PMID 15934862.
- Price PA, Faus SA, Williamson MK (September 1998). "Warfarin causes rapid calcification of the elastic lamellae in rat arteries and heart valves". Arteriosclerosis, Thrombosis, and Vascular Biology. 18 (9): 1400–7. doi:10.1161/01.atv.18.9.1400. PMID 9743228.
- Schurgers LJ, Aebert H, Vermeer C, Bültmann B, Janzen J (November 2004). "Oral anticoagulant treatment: friend or foe in cardiovascular disease?". Blood. 104 (10): 3231–2. doi:10.1182/blood-2004-04-1277. PMID 15265793.
- Koos R, Mahnken AH, Mühlenbruch G, Brandenburg V, Pflueger B, Wildberger JE, Kühl HP (September 2005). "Relation of oral anticoagulation to cardiac valvular and coronary calcium assessed by multislice spiral computed tomography". The American Journal of Cardiology. 96 (6): 747–9. doi:10.1016/j.amjcard.2005.05.014. PMID 16169351.
- Zieman SJ, Melenovsky V, Kass DA (May 2005). "Mechanisms, pathophysiology, and therapy of arterial stiffness". Arteriosclerosis, Thrombosis, and Vascular Biology. 25 (5): 932–43. doi:10.1161/01.atv.0000160548.78317.29. PMID 15731494.
- Raggi P, Shaw LJ, Berman DS, Callister TQ (May 2004). "Prognostic value of coronary artery calcium screening in subjects with and without diabetes". Journal of the American College of Cardiology. 43 (9): 1663–9. doi:10.1016/j.jacc.2003.09.068. PMID 15120828.
- Walther B, Karl JP, Booth SL, Boyaval P (July 2013). "Menaquinones, bacteria, and the food supply: the relevance of dairy and fermented food products to vitamin K requirements". Advances in Nutrition. 4 (4): 463–73. doi:10.3945/an.113.003855. PMC 3941825. PMID 23858094.