Transforming protein RhoA
Transforming protein RhoA, also known as Ras homolog family member A (RhoA), is a small GTPase protein in the Rho family of GTPases that in humans is encoded by the RHOA gene.[5] While the effects of RhoA activity are not all well known, it is primarily associated with cytoskeleton regulation, mostly actin stress fibers formation and actomyosin contractility. It acts upon several effectors. Among them, ROCK1 (Rho-associated, coiled-coil containing protein kinase 1) and DIAPH1 (Diaphanous Homologue 1, a.k.a. hDia1, homologue to mDia1 in mouse, diaphanous in Drosophila) are the best described. RhoA, and the other Rho GTPases, are part of a larger family of related proteins known as the Ras superfamily, a family of proteins involved in the regulation and timing of cell division. RhoA is one of the oldest Rho GTPases, with homologues present in the genomes since 1.5 billion years. As a consequence, RhoA is somehow involved in many cellular processes which emerged throughout evolution. RhoA specifically is regarded as a prominent regulatory factor in other functions such as the regulation of cytoskeletal dynamics, transcription, cell cycle progression and cell transformation.
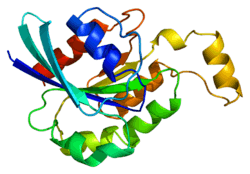


Structure
The specific gene that encodes RhoA, RHOA, is located on chromosome 3 and consists of four exons,[6] which has also been linked as a possible risk factor for atherothrombolic stroke.
Similar to other GTPases, RhoA presents a Rho insert in its primary sequence in the GTPase domain. RhoA contains also four insertion or deletion sites with an extra helical subdomain; these sites are characteristic of many GTPases in the Rho family. Most importantly, RhoA contains two switch regions, Switch I and Switch II, whose conformational states are modified following the activation or inactivation of the protein. Both of these switches have characteristic folding, correspond to specific regions on the RhoA coil and are uniformly stabilized via hydrogen bonds. The conformations of the Switch domains are modified depending on the binding of either GDP or GTP to RhoA. The nature of the bound nucleotide and the ensuing conformational modification of the Switch domains dictates the ability of RhoA to bind or not with partner proteins (see below).
The primary protein sequences of members of the Rho family are mostly identical, with the N-terminal containing most of the protein coding for GTP binding and hydrolysis. The C-terminal of RhoA is modified via prenylation, anchoring the GTPase into membranes, which is essential for its role in cell growth and cytoskeleton organization. Key amino acids that are involved in the stabilization and regulation of GTP hydrolysis are conserved in RhoA as Gly14, Thr19, Phe30 and Gln63.
Correct localization of the RhoA proteins is heavily dependent on the C-terminus; during prenylation, the anchoring of the prenyl group is essential for the stability, inhibition of and the synthesis of enzymes and proliferation. RhoA is sequestered by dissociation inhibitors (RhoGDIs) which remove the protein from the membrane while preventing its further interaction with other downstream effectors.[7]
Activation mechanism
RhoA acquires both inactive GDP-bound and active GTP-bound conformational states; these states alternate between the active and inactive states via the exchange of GDP to GTP (conducted simultaneously via guanine nucleotide exchange factors and GTPase activating factor). RhoA is activated primarily by guanine nucleotide exchange factors (GEFs) via phosphorylation; due to large network of overlapping phosphorylation, a multitude of GEFs are utilized to enable specific signaling pathways. These structural arrangements provide interaction sites that can interact with effectors and guanine factors in order to stabilize and signal the hydrolysis of GTP.[8]
Participation in cellular processes
RhoA is primarily involved in these activities: actin organization, myosin contractility, cell cycle maintenance, cellular morphological polarization, cellular development and transcriptional control.
Actin organization
RhoA is prevalent in regulating cell shape, polarity and locomotion via actin polymerization, actomyosin contractility, cell adhesion, and microtubule dynamics. In addition, RhoA is believed to act primarily at the rear (uropod) of migrating cells to promote detachment, similar to the attachment and detachment process found in the focal adhesion mechanism. Signal transduction pathways regulated via RhoA link plasma membrane receptors to focal adhesion formation and the subsequent activation of relevant actin stress fibers. RhoA directly stimulates actin polymerization through activation of diaphanous-related formins, thereby structurally changing the actin monomers to filaments. ROCK kinases induce actomyosin-based contractility and phosphorylate TAU and MAP2 involved in regulating myosins and other actin-binding proteins in order to assist in cell migration and detachment. The concerted action of ROCK and Dia is essential for the regulation of cell polarity and organization of microtubules. RhoA also regulates the integrity of the extracellular matrix and the loss of corresponding cell-cell adhesions (primarily adherens and tight junctions) required for the migration of epithelial. RhoA's role in signal transduction mediation is also attributed to the establishment of tissue polarity in epidermal structures due to its actin polymerization to coordinate vesicular motion;[9] movement within actin filaments forms webs that move in conjunction with vesicular linear motion. As a result, mutations present in the polarity genes indicate that RhoA is critical for tissue polarity and directed intracellular movement.
Cell development
RhoA is required for processes involving cell development, some of which include outgrowth, dorsal closure, bone formation, and myogenesis. Loss of RhoA function is frequently attributed to failed gastrulation and cell migration inability. In extension, RhoA has been shown to function as an intermediary switch within the overall mechanically mediated process of stem cell commitment and differentiation. For example, human mesenchymal stem cells and their differentiation into adipocytes or osteocytes are direct results of RhoA's impact on cell shape, signaling and cytoskeletal integrity. Cell shape acts as the primary mechanical cue that drives RhoA activity and downstream effector ROCK activity to control stem cell commitment and cytoskeletal maintenance.[10] Transforming growth factor (TGF)-mediated pathways that control tumor progression and identity are also frequently noted to be RhoA-dependent mechanisms. TGF-β1, a tumor suppressive growth factor, is known to regulate growth, differentiation and epithelial transformation in tumorigenesis. Instead of blocking growth, TGF-β1 directly activates RhoA in epithelial cells while blocking its downstream target, p160; as a result, activated RhoA-dependent pathways induce stress fiber formation and subsequent mesenchymal properties .[11]
Transcriptional control
Activated RhoA also participates in regulating transcriptional control over other signal transduction pathways via various cellular factors. RhoA proteins help potentiate the transcription independent of ternary complex factors when activated while simultaneously modulating subsequent extracellular signal activity. It has also been shown that RhoA mediates serum-, LPA- and AIF4-induced signaling pathways in addition to regulating the transcription of the c-fos promoter, a key component in the formation of the ternary complex producing the serum and ternary factors.[12] RhoA signaling and modulation of actin polymerization also regulates Sox9 expression via controlling transcriptional Sox9 activity. The expression and transcriptional activity of Sox9 is directly linked with the loss of RhoA activity and illustrates how RhoA participates in the transcriptional control of specific protein expression.[13]
Cell cycle maintenance
RhoA as well as several other members of the Rho family are identified as having roles in the regulation of the cytoskeleton and cell division. RhoA plays a pivotal role in G1 cell cycle progression, primarily through regulation of cyclin D1 and cyclin-dependent kinase inhibitors (p21 and p27) expression. These regulation pathways activate protein kinases, which subsequently modulate transcription factor activity. RhoA specifically suppresses p21 levels in normal and transformed cell lines via a p53-independent transcriptional mechanism while p27 levels are regulated using effector Rho-associated kinases. Cytokinesis is defined by actomyosin-based contraction. RhoA-dependent diaphanous-related formins (DRFs) localize to the cleavage furrow during cytokinesis while stimulating local actin polymerization by coordinating microtubules with actin filaments at the site of the myosin contractile ring. Differences in effector binding distinguish RhoA amongst other related Ras homologs GTPases. Integrins can modulate RhoA activity depending on the extracellular matrix composition and other relevant factors. Similarly, RhoA's stimulation of PKN2 kinase activity regulates cell-cell adhesion through apical junction formation and disassembly.[7][14] Though RhoA is most easily recognized from its unique contributions in actin-myosin contractility and stress fiber formation, new research has also identified it as a key factor in the mediation of membrane ruffling, lamellae formation and membrane blebbing. A majority of this activity occurs in the leading edge of cells during migration in coordination with membrane protrusions of breast carcinoma.[15]
RhoA pathway
Molecules act on various receptors, such as NgR1, LINGO1, p75, TROY and other unknown receptors (e.g. by CSPGs), which stimulates RhoA. RhoA activates ROCK (RhoA kinase) which stimulates LIM kinase, which then inhibits cofilin, which effectively reorganizes the actin cytoskeleton of the cell.[5] In the case of neurons, activation of this pathway results in growth cone collapse, therefore inhibits the growth and repair of neural pathways and axons. Inhibition of this pathway by its various components usually results in some level of improved re-myelination.[16][17][18][19] After global ischemia, hyperbaric oxygen (at least at 3 ATA) appears to partially suppress expression of RhoA, in addition to Nogo protein (Reticulon 4), and a subunit of its receptor Ng-R.[20] The MEMO1-RhoA-DIAPH1 signaling pathway plays an important role in ERBB2-dependent stabilization of microtubules at the cell cortex. A recent study shows that RhoA-Rho kinase signaling mediates thrombin-induced brain damage.[21]
Interactions
RHOA has been shown to interact with:
- ARHGAP1,[22][23][24][25]
- ARHGAP5,[26]
- ARHGDIA,[27][28][29][30][31]
- ARHGEF11,[32]
- ARHGEF12,[33]
- ARHGEF3,[34]
- CIT[35][36]
- DGKQ,[37]
- DIAPH1,[35]
- GEFT,[38]
- ITPR1,[35][39]
- KCNA2,[40]
- KTN1,[41][42][43]
- PKN2,[44][45][46]
- PLCG1,[47]
- Phospholipase D1,[48][49]
- Protein kinase N1,[35][45][50]
- RAP1GDS1,[51]
- RICS,[52][53][53]
- ROCK1,[54][55]
- TRIO,[56] and
- TRPC1.[39]
Clinical significance
Cancer
Given that its overexpression is found in many malignancies, RhoA activity has been linked within several cancer applications due to its significant involvement in cancer signaling cascades. Serum response factors (SRFs) are known to mediate androgen receptors in prostate cancer cells, including roles ranging from distinguishing benign from malignant prostate and identifying aggressive disease. RhoA mediates androgen-responsiveness of these SRF genes; as a result, interference with RhoA has been shown to prevent the androgen regulation of SRF genes. In application, RhoA expression is notably higher in malignant prostate cancer cells compared to benign prostate cells, with elevated RhoA expression being associated with elevated lethality and aggressive proliferation. On the other hand, silencing RhoA lessened androgen-regulated cell viability and handicapped prostate cancer cell migration.[57]
RhoA has also been found to be hyper activated in gastric cancer cells; in consequence, suppression of RhoA activity partially reversed the proliferation phenotype of gastric cancer cells via the down-regulation of the RhoA-mammalian Diaphanous 1 pathway.[58] Doxorubicin has been referred to frequently as a highly-promising anti-cancer drug that is also being utilized in chemotherapy treatments; however, as with nearly all chemotherapeutics, the issue of drug resistance remains. Minimizing or postponing this resistance would the necessary dose to eradicate the tumor, thus diminishing drug toxicity. Subsequent RhoA expression decrease has also been associated with increased sensitivity to doxorubicin and the complete reversion of doxorubicin resistance in certain cells; this shows the resilience of RhoA as a consistent indicator anti-cancer activity. In addition to promoting tumor-suppression activity, RhoA also has inherent impact upon the efficacy of drugs in relation to cancer functionality and could be applied to gene therapy protocols in future research.[59]
Protein expression of RhoA has been identified to be significantly higher in testicular tumor tissue than that in nontumor tissue; expression of protein for RhoA, ROCK-I, ROCK-II, Rac1, and Cdc42 was greater in tumors of higher stages than lower stages, coinciding with greater lymph metastasis and invasion in upper urinary tract cancer. Although both RhoA and RhoC proteins comprise a significant part of the Rho GTPases that are linked to promoting the invasive behavior of breast carcinomas, attributing specific functions to these individual members has been difficult. We have used a stable retroviral RNA interference approach to generate invasive breast carcinoma cells (SUM-159 cells) that lack either RhoA or RhoC expression. Analysis of these cells enabled us to deduce that RhoA impedes and RhoC stimulates invasion. Unexpectedly, this analysis also revealed a compensatory relationship between RhoA and RhoC at the level of both their expression and activation, and a reciprocal relationship between RhoA and Rac1 activation. Chronic Myeloid Leukemia (CML), a stem cell disorder that prevents myeloid cells from functioning correctly, has been linked to actin polymerization. Signaling proteins like RhoA, regulate polymerization of actin. Due to differences proteins exhibited between normal and affected neutrocytes, RhoA has become the key element; further experimentation has also shown that RhoA-inhibiting pathways prevent the overall growth of CML cells. As a result, RhoA has significant potential as a therapeutic target in gene therapy techniques to treating CML.[60] Therefore, RhoA's role in the proliferation of cancer cell phenotypes is a key application that can be applied to targeted cancer therapeutics and the development for pharmaceuticals.
Drug applications
In June 2012, a new drug candidate named "Rhosin" was synthesized by researchers at the Cincinnati Children's Hospital, a drug with the full intention to inhibit cancer proliferation and promote nerve cell regeneration. This inhibitor specifically targets Rho GTPases to prevent cell growth related to cancer. When tested on breast cancer cells, Rhosin inhibited growth and the growth of mammary spheres in a dose dependent manner, functioning as targets for RhoA while simultaneously maintaining the integrity of normal cellular processes and normal breast cells. These promising results indicate Rhosin's general effectiveness in preventing breast cancer proliferation via RhoA targeting.[61]
Possible target for asthma and diabetes drugs
RhoA's physiological functions have been linked to the contraction and migration of cells which are manifested as symptoms in both asthma and diabetes (i.e. airflow limitation and hyper-responsiveness, desensitization, etc.). Due to pathophysiological overlap of RhoA and Rho-kinase in asthma, both RhoA and Rho-kinase have become promising new target molecules for pharmacological research to develop alternate forms of treatment for asthma.[62] RhoA and Rho kinase mechanisms have been linked to diabetes due to the up-regulated expression of targets within type 1 and 2 diabetic animals. Inhibition of this pathway prevented and ameliorated pathologic changes in diabetic complications, indicating that RhoA pathway is a promising target for therapeutic development in diabetes treatment[63]
References
- GRCh38: Ensembl release 89: ENSG00000067560 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000007815 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Kiss C, Li J, Szeles A, Gizatullin RZ, Kashuba VI, Lushnikova T, Protopopov AI, Kelve M, Kiss H, Kholodnyuk ID, Imreh S, Klein G, Zabarovsky ER (1997). "Assignment of the ARHA and GPX1 genes to human chromosome bands 3p21.3 by in situ hybridization and with somatic cell hybrids". Cytogenet. Cell Genet. 79 (3–4): 228–230. doi:10.1159/000134729. PMID 9605859.
- Oliyarnyk, Olena; et al. (2005). "Interindividual differences of response to statin treatment cannot be explained by variations of the human gene for RhoA". Biochemical Genetics. 43 (3): 143–148. doi:10.1007/s10528-005-1507-0. PMID 15932062.
- Wheeler AP, Ridley AJ (2004). "Why three Rho proteins? RhoA, RhoB, RhoC, and cell motility". Exp. Cell Res. 301 (1): 43–9. doi:10.1016/j.yexcr.2004.08.012. PMID 15501444.
- Ihara K, Muraguchi S, Kato M, Shimizu T, Shirakawa M, Kuroda S, Kaibuchi K, Hakoshima T (1998). "Crystal structure of human RhoA in a dominantly active form complexed with a GTP analogue". Journal of Biological Chemistry. 273 (16): 9656–9666. doi:10.1074/jbc.273.16.9656. PMID 9545299.
- Strutt DI, Weber U, Mlodzik M (1997). "The role of RhoA in tissue polarity and Frizzled signalling". Nature. 387 (6630): 292–5. doi:10.1038/387292a0. PMID 9153394.
- McBeath R, Pirone DM, Nelson CM, Bhadriraju K, Chen CS (2004). "Cell shape, cytoskeletal tension, and RhoA regulate stem cell lineage commitment". Developmental Cell. 6 (4): 483–495. doi:10.1016/S1534-5807(04)00075-9. PMID 15068789.
- Bhowmick NA, Ghiassi M, Bakin A, Aakre M, Lundquist CA, Engel ME, Arteaga CL, Moses HL (2001). "Transforming growth factor-β1 mediates epithelial to mesenchymal transdifferentiation through a RhoA-dependent mechanism". Molecular Biology of the Cell. 12 (1): 27–36. doi:10.1091/mbc.12.1.27. PMC 30565. PMID 11160820.
- Hill, Caroline S., Judy Wynne, and Richard Treisman (1995). "The Rho family GTPases RhoA, Racl, and CDC42Hs regulate transcriptional activation by SRF". Cell. 81 (7): 1159–1170. doi:10.1016/S0092-8674(05)80020-0. PMID 7600583.CS1 maint: multiple names: authors list (link)
- Kumar, Deepak; Lassar, Andrew B. (2009). "The transcriptional activity of Sox9 in chondrocytes is regulated by RhoA signaling and actin polymerization". Molecular and Cellular Biology. 29 (15): 4262–4273. doi:10.1128/MCB.01779-08. PMC 2715793. PMID 19470758.
- Wheeler, Ann P.; Ann J. Ridley (2004). "Why three Rho proteins? RhoA, RhoB, RhoC, and cell motility". Experimental Cell Research. 301 (1): 43–49. doi:10.1016/j.yexcr.2004.08.012. PMID 15501444.
- O’ Connor K. and Chen M. (2013). "Dynamic functions of RhoA in tumor cell migration and invasion". Small GTPases. 4 (3): 141–147. doi:10.4161/sgtp.25131. PMC 3976970. PMID 24025634.
- Yiu G, He Z (August 2006). "Glial inhibition of CNS axon regeneration". Nat. Rev. Neurosci. 7 (8): 617–627. doi:10.1038/nrn1956. PMC 2693386. PMID 16858390.
- Bradbury EJ, McMahon SB (August 2006). "Spinal cord repair strategies: why do they work?". Nat. Rev. Neurosci. 7 (8): 644–653. doi:10.1038/nrn1964. PMID 16858392.
- Karnezis T, Mandemakers W, McQualter JL, Zheng B, Ho PP, Jordan KA, Murray BM, Barres B, Tessier-Lavigne M, Bernard CC (July 2004). "The neurite outgrowth inhibitor Nogo A is involved in autoimmune-mediated demyelination". Nat. Neurosci. 7 (7): 736–744. doi:10.1038/nn1261. PMID 15184901.
- Bregman BS, Kunkel-Bagden E, Schnell L, Dai HN, Gao D, Schwab ME (November 1995). "Recovery from spinal cord injury mediated by antibodies to neurite growth inhibitors". Nature. 378 (6556): 498–501. doi:10.1038/378498a0. PMID 7477407.
- Yiu G, He Z (September 2003). "Glial inhibition of CNS axon regeneration". Biochem. Biophys. Res. Commun. 309 (2): 368–76. doi:10.1016/j.bbrc.2003.08.006. PMID 12951059.
- Han X, Lan X, Li Q, Gao Y, Zhu W, Cheng T, Maruyama T, Wang J (2015). "Inhibition of prostaglandin E2 receptor EP3 mitigates thrombin-induced brain injury". J Cereb Blood Flow Metab. 36 (6): 1059–74. doi:10.1177/0271678X15606462. PMC 4908617. PMID 26661165.
- Rual JF, Venkatesan K, Hao T, Hirozane-Kishikawa T, Dricot A, Li N, Berriz GF, Gibbons FD, Dreze M, Ayivi-Guedehoussou N, Klitgord N, Simon C, Boxem M, Milstein S, Rosenberg J, Goldberg DS, Zhang LV, Wong SL, Franklin G, Li S, Albala JS, Lim J, Fraughton C, Llamosas E, Cevik S, Bex C, Lamesch P, Sikorski RS, Vandenhaute J, Zoghbi HY, Smolyar A, Bosak S, Sequerra R, Doucette-Stamm L, Cusick ME, Hill DE, Roth FP, Vidal M (October 2005). "Towards a proteome-scale map of the human protein-protein interaction network". Nature. 437 (7062): 1173–8. doi:10.1038/nature04209. PMID 16189514.
- Zhang B, Zheng Y (April 1998). "Regulation of RhoA GTP hydrolysis by the GTPase-activating proteins p190, p50RhoGAP, Bcr, and 3BP-1". Biochemistry. 37 (15): 5249–57. doi:10.1021/bi9718447. PMID 9548756.
- Li R, Zhang B, Zheng Y (December 1997). "Structural determinants required for the interaction between Rho GTPase and the GTPase-activating domain of p190". J. Biol. Chem. 272 (52): 32830–5. doi:10.1074/jbc.272.52.32830. PMID 9407060.
- Zhang B, Chernoff J, Zheng Y (April 1998). "Interaction of Rac1 with GTPase-activating proteins and putative effectors. A comparison with Cdc42 and RhoA". J. Biol. Chem. 273 (15): 8776–82. doi:10.1074/jbc.273.15.8776. PMID 9535855.
- Wennerberg K, Forget MA, Ellerbroek SM, Arthur WT, Burridge K, Settleman J, Der CJ, Hansen SH (July 2003). "Rnd proteins function as RhoA antagonists by activating p190 RhoGAP". Curr. Biol. 13 (13): 1106–15. doi:10.1016/s0960-9822(03)00418-4. PMC 6918695. PMID 12842009.
- Ewing RM, Chu P, Elisma F, Li H, Taylor P, Climie S, McBroom-Cerajewski L, Robinson MD, O'Connor L, Li M, Taylor R, Dharsee M, Ho Y, Heilbut A, Moore L, Zhang S, Ornatsky O, Bukhman YV, Ethier M, Sheng Y, Vasilescu J, Abu-Farha M, Lambert JP, Duewel HS, Stewart II, Kuehl B, Hogue K, Colwill K, Gladwish K, Muskat B, Kinach R, Adams SL, Moran MF, Morin GB, Topaloglou T, Figeys D (2007). "Large-scale mapping of human protein-protein interactions by mass spectrometry". Mol. Syst. Biol. 3 (1): 89. doi:10.1038/msb4100134. PMC 1847948. PMID 17353931.
- Gajate C, Mollinedo F (March 2005). "Cytoskeleton-mediated death receptor and ligand concentration in lipid rafts forms apoptosis-promoting clusters in cancer chemotherapy". J. Biol. Chem. 280 (12): 11641–7. doi:10.1074/jbc.M411781200. PMID 15659383.
- Michaelson D, Silletti J, Murphy G, D'Eustachio P, Rush M, Philips MR (January 2001). "Differential localization of Rho GTPases in live cells: regulation by hypervariable regions and RhoGDI binding". J. Cell Biol. 152 (1): 111–26. doi:10.1083/jcb.152.1.111. PMC 2193662. PMID 11149925.
- Gorvel JP, Chang TC, Boretto J, Azuma T, Chavrier P (January 1998). "Differential properties of D4/LyGDI versus RhoGDI: phosphorylation and rho GTPase selectivity". FEBS Lett. 422 (2): 269–73. doi:10.1016/s0014-5793(98)00020-9. PMID 9490022.
- Fauré J, Dagher MC (May 2001). "Interactions between Rho GTPases and Rho GDP dissociation inhibitor (Rho-GDI)". Biochimie. 83 (5): 409–14. doi:10.1016/s0300-9084(01)01263-9. PMID 11368848.
- Rümenapp U, Blomquist A, Schwörer G, Schablowski H, Psoma A, Jakobs KH (October 1999). "Rho-specific binding and guanine nucleotide exchange catalysis by KIAA0380, a dbl family member". FEBS Lett. 459 (3): 313–8. doi:10.1016/s0014-5793(99)01270-3. PMID 10526156.
- Suzuki N, Nakamura S, Mano H, Kozasa T (January 2003). "Galpha 12 activates Rho GTPase through tyrosine-phosphorylated leukemia-associated RhoGEF". Proc. Natl. Acad. Sci. U.S.A. 100 (2): 733–8. doi:10.1073/pnas.0234057100. PMC 141065. PMID 12515866.
- Arthur WT, Ellerbroek SM, Der CJ, Burridge K, Wennerberg K (November 2002). "XPLN, a guanine nucleotide exchange factor for RhoA and RhoB, but not RhoC". J. Biol. Chem. 277 (45): 42964–72. doi:10.1074/jbc.M207401200. PMID 12221096.
- Riento K, Guasch RM, Garg R, Jin B, Ridley AJ (June 2003). "RhoE binds to ROCK I and inhibits downstream signaling". Mol. Cell. Biol. 23 (12): 4219–29. doi:10.1128/mcb.23.12.4219-4229.2003. PMC 156133. PMID 12773565.
- Madaule P, Furuyashiki T, Reid T, Ishizaki T, Watanabe G, Morii N, Narumiya S (December 1995). "A novel partner for the GTP-bound forms of rho and rac". FEBS Lett. 377 (2): 243–8. doi:10.1016/0014-5793(95)01351-2. PMID 8543060.
- Houssa B, de Widt J, Kranenburg O, Moolenaar WH, van Blitterswijk WJ (March 1999). "Diacylglycerol kinase theta binds to and is negatively regulated by active RhoA". J. Biol. Chem. 274 (11): 6820–2. doi:10.1074/jbc.274.11.6820. PMID 10066731.
- Lutz S, Freichel-Blomquist A, Rümenapp U, Schmidt M, Jakobs KH, Wieland T (May 2004). "p63RhoGEF and GEFT are Rho-specific guanine nucleotide exchange factors encoded by the same gene". Naunyn Schmiedebergs Arch. Pharmacol. 369 (5): 540–6. doi:10.1007/s00210-004-0926-5. PMID 15069594.
- Mehta D, Ahmmed GU, Paria BC, Holinstat M, Voyno-Yasenetskaya T, Tiruppathi C, Minshall RD, Malik AB (August 2003). "RhoA interaction with inositol 1,4,5-trisphosphate receptor and transient receptor potential channel-1 regulates Ca2+ entry. Role in signaling increased endothelial permeability". J. Biol. Chem. 278 (35): 33492–500. doi:10.1074/jbc.M302401200. PMID 12766172.
- Cachero TG, Morielli AD, Peralta EG (June 1998). "The small GTP-binding protein RhoA regulates a delayed rectifier potassium channel". Cell. 93 (6): 1077–85. doi:10.1016/s0092-8674(00)81212-x. PMID 9635436.
- Neudauer CL, Joberty G, Macara IG (January 2001). "PIST: a novel PDZ/coiled-coil domain binding partner for the rho-family GTPase TC10". Biochem. Biophys. Res. Commun. 280 (2): 541–7. doi:10.1006/bbrc.2000.4160. PMID 11162552.
- Hotta K, Tanaka K, Mino A, Kohno H, Takai Y (August 1996). "Interaction of the Rho family small G proteins with kinectin, an anchoring protein of kinesin motor". Biochem. Biophys. Res. Commun. 225 (1): 69–74. doi:10.1006/bbrc.1996.1132. PMID 8769096.
- Vignal E, Blangy A, Martin M, Gauthier-Rouvière C, Fort P (December 2001). "Kinectin is a key effector of RhoG microtubule-dependent cellular activity". Mol. Cell. Biol. 21 (23): 8022–34. doi:10.1128/MCB.21.23.8022-8034.2001. PMC 99969. PMID 11689693.
- Quilliam LA, Lambert QT, Mickelson-Young LA, Westwick JK, Sparks AB, Kay BK, Jenkins NA, Gilbert DJ, Copeland NG, Der CJ (November 1996). "Isolation of a NCK-associated kinase, PRK2, an SH3-binding protein and potential effector of Rho protein signaling". J. Biol. Chem. 271 (46): 28772–6. doi:10.1074/jbc.271.46.28772. PMID 8910519.
- Flynn P, Mellor H, Palmer R, Panayotou G, Parker PJ (January 1998). "Multiple interactions of PRK1 with RhoA. Functional assignment of the Hr1 repeat motif". J. Biol. Chem. 273 (5): 2698–705. doi:10.1074/jbc.273.5.2698. PMID 9446575.
- Gebbink MF, Kranenburg O, Poland M, van Horck FP, Houssa B, Moolenaar WH (June 1997). "Identification of a novel, putative Rho-specific GDP/GTP exchange factor and a RhoA-binding protein: control of neuronal morphology". J. Cell Biol. 137 (7): 1603–13. doi:10.1083/jcb.137.7.1603. PMC 2137826. PMID 9199174.
- Thodeti CK, Massoumi R, Bindslev L, Sjölander A (July 2002). "Leukotriene D4 induces association of active RhoA with phospholipase C-gamma1 in intestinal epithelial cells". Biochem. J. 365 (Pt 1): 157–63. doi:10.1042/BJ20020248. PMC 1222665. PMID 12071848.
- Genth H, Schmidt M, Gerhard R, Aktories K, Just I (February 2003). "Activation of phospholipase D1 by ADP-ribosylated RhoA". Biochem. Biophys. Res. Commun. 302 (1): 127–32. doi:10.1016/s0006-291x(03)00112-8. PMID 12593858.
- Cai S, Exton JH (May 2001). "Determination of interaction sites of phospholipase D1 for RhoA". Biochem. J. 355 (Pt 3): 779–85. doi:10.1042/bj3550779. PMC 1221795. PMID 11311142.
- Alberts AS, Bouquin N, Johnston LH, Treisman R (April 1998). "Analysis of RhoA-binding proteins reveals an interaction domain conserved in heterotrimeric G protein beta subunits and the yeast response regulator protein Skn7". J. Biol. Chem. 273 (15): 8616–22. doi:10.1074/jbc.273.15.8616. PMID 9535835.
- Vikis HG, Stewart S, Guan KL (April 2002). "SmgGDS displays differential binding and exchange activity towards different Ras isoforms". Oncogene. 21 (15): 2425–32. doi:10.1038/sj.onc.1205306. PMID 11948427.
- Nakazawa T, Watabe AM, Tezuka T, Yoshida Y, Yokoyama K, Umemori H, Inoue A, Okabe S, Manabe T, Yamamoto T (July 2003). "p250GAP, a novel brain-enriched GTPase-activating protein for Rho family GTPases, is involved in the N-methyl-d-aspartate receptor signaling". Mol. Biol. Cell. 14 (7): 2921–34. doi:10.1091/mbc.E02-09-0623. PMC 165687. PMID 12857875.
- Nakamura T, Komiya M, Sone K, Hirose E, Gotoh N, Morii H, Ohta Y, Mori N (December 2002). "Grit, a GTPase-activating protein for the Rho family, regulates neurite extension through association with the TrkA receptor and N-Shc and CrkL/Crk adapter molecules". Mol. Cell. Biol. 22 (24): 8721–34. doi:10.1128/mcb.22.24.8721-8734.2002. PMC 139861. PMID 12446789.
- Leung T, Chen XQ, Manser E, Lim L (October 1996). "The p160 RhoA-binding kinase ROK alpha is a member of a kinase family and is involved in the reorganization of the cytoskeleton". Mol. Cell. Biol. 16 (10): 5313–27. doi:10.1128/mcb.16.10.5313. PMC 231530. PMID 8816443.
- Fujisawa K, Fujita A, Ishizaki T, Saito Y, Narumiya S (September 1996). "Identification of the Rho-binding domain of p160ROCK, a Rho-associated coiled-coil containing protein kinase". J. Biol. Chem. 271 (38): 23022–8. doi:10.1074/jbc.271.38.23022. PMID 8798490.
- Medley QG, Serra-Pagès C, Iannotti E, Seipel K, Tang M, O'Brien SP, Streuli M (November 2000). "The trio guanine nucleotide exchange factor is a RhoA target. Binding of RhoA to the trio immunoglobulin-like domain". J. Biol. Chem. 275 (46): 36116–23. doi:10.1074/jbc.M003775200. PMID 10948190.
- Schmidt, Lucy J. (2012). "RhoA as a mediator of clinically relevant androgen action in prostate cancer cells". Molecular Endocrinology. 26 (5): 716–735. doi:10.1210/me.2011-1130. PMC 3355556. PMID 22456196.
- Zhang S, Tang Q, Xu F, Xue Y, Zhen Z, Deng Y, Liu M, Chen J, Liu S, Qiu M, Liao Z, Li Z, Luo D, Shi F, Zheng Y, Bi F (2009). "RhoA regulates G1-S progression of gastric cancer cells by modulation of multiple INK4 family tumor suppressors". Molecular Cancer Research. 7 (4): 570–580. doi:10.1158/1541-7786.MCR-08-0248. PMID 19372585.
- Doublier, Sophie; et al. (2008). "RhoA silencing reverts the resistance to doxorubicin in human colon cancer cells". Molecular Cancer Research. 6 (10): 1607–1620. doi:10.1158/1541-7786.MCR-08-0251. PMID 18922976.
- Molli PR, Pradhan MB, Advani SH, Naik NR (2012). "RhoA: A therapeutic target for chronic myeloid leukemia". Molecular Cancer. 11 (1): 16. doi:10.1186/1476-4598-11-16. PMC 3353160. PMID 22443473.
- Shang X, Marchioni F, Sipes N, Evelyn CR, Jerabek-Willemsen M, Duhr S, Seibel W, Wortman M, Zheng Y (2012). "Rational design of small molecule inhibitors targeting RhoA subfamily Rho GTPases". Chemistry & Biology. 19 (6): 699–710. doi:10.1016/j.chembiol.2012.05.009. PMC 3383629. PMID 22726684.
- Kume H (2008). "RhoA/Rho-kinase as a therapeutic target in asthma". Curr. Med. Chem. 15 (27): 2876–85. doi:10.2174/092986708786242831. PMID 18991642.
- Zhou H, Li YJ (2010). "RhoA/Rho kinase: a novel therapeutic target in diabetic complications". Chin. Med. J. 123 (17): 2461–6. PMID 21034566.
Further reading
- Ramakers GJ (2002). "Rho proteins, mental retardation and the cellular basis of cognition". Trends Neurosci. 25 (4): 191–199. doi:10.1016/S0166-2236(00)02118-4. PMID 11998687.
- Chang ZF, Lee HH (2006). "RhoA signaling in phorbol ester-induced apoptosis". J. Biomed. Sci. 13 (2): 173–180. doi:10.1007/s11373-005-9056-4. PMID 16496227.
External links
- rhoA+Protein at the US National Library of Medicine Medical Subject Headings (MeSH)
- RHOA Info with links in the Cell Migration Gateway
- Overview of all the structural information available in the PDB for UniProt: P61586 (Human Transforming protein RhoA) at the PDBe-KB.
- Overview of all the structural information available in the PDB for UniProt: Q9QUI0 (Mouse Transforming protein RhoA) at the PDBe-KB.
PDB gallery | |
|---|---|
|