Haplogroup E-V68
Haplogroup E-V68, also known as E1b1b1a, is a major human Y-chromosome DNA haplogroup found in North Africa, the Horn of Africa, Western Asia and Europe. It is a subclade of the larger and older haplogroup, known as E1b1b or E-M215 (also roughly equivalent to E-M35). The E1b1b1a lineage is identified by the presence of a single nucleotide polymorphism (SNP) mutation on the Y chromosome, which is known as V68. It is a subject of discussion and study in genetics as well as genetic genealogy, archaeology, and historical linguistics.
| Haplogroup E-V68 | |
|---|---|
| Possible time of origin | c. 24,000 years BP[1] |
| Coalescence age | c. 19,900 years BP[1] |
| Possible place of origin | Egypt/Libya[2] or southern Egypt/northern Sudan[3] |
| Ancestor | E-M215-M35[4] |
| Descendants | E-M78[4], E-V1039 |
| Defining mutations | V68, L539, PF2203[4] |
E-V68 is dominated by its longer-known subclade E-M78. In various publications, both E-V68 and E-M78 have been referred to by other names, especially phylogenetic nomenclature such as "E3b1a" which are designed to show their place on the family tree of all human males. These various names change as new discoveries are made and are discussed below.
Origins
E-M78, like its parent clade E-V68, is thought to have an African origin. Based on genetic STR variance data, Cruciani et al. (2007) suggests that this subclade originated in "Northeastern Africa", which in the study refers specifically to the region of Egypt and Libya.[5]
Prior to Cruciani et al. (2007), Semino et al. (2004) had proposed a place of origin for E-M78 further south in East Africa. This was because of the high frequency and diversity of E-M78 lineages in the region of Ethiopia. However, Cruciani et al. (2007) were able to study more data, and concluded that the E-M78 lineages in the Horn of Africa were dominated by relatively recent branches (see E-V32 below). They concluded that the region of Egypt was the likely place of origin of E-M78 based on "the peripheral geographic distribution of the most derived subhaplogroups with respect to northeastern Africa, as well as the results of quantitative analysis of UEP and microsatellite diversity".
Cruciani et al. (2007) also note this as evidence for "a corridor for bidirectional migrations" between Northeast Africa (Egypt and Libya in their data) on the one hand and East Africa on the other. Because Cruciani et al. (2007) also proposed that E-M35, the parent clade of E-M78, originated in East Africa during the paleolithic and subsequently spread to the region of Egypt. E-M78 in East Africa, is therefore the result of a back migration. The authors believe there were "at least 2 episodes between 23.9–17.3 ky and 18.0–5.9 ky ago".
Another probable migration to the south from Egypt was noted by Hassan et al. (2008) based upon their survey of Sudan. Specifically E-V12 and E-V22, "might have been brought to Sudan from North Africa after the progressive desertification of the Sahara around 6,000-8,000 years ago".
Northwards from Egypt and Libya, E-M78 migrated into the Middle East, but additionally Trombetta et al. (2011) proposed that the earlier E-V68 carrying population may have migrated by sea directly from Africa to southwestern Europe, because they observed cases of E-V68* (without the M78 mutation) only in Sardinia, and not in the Middle Eastern samples. Concerning E-M78, like other forms of E-V68 there is evidence of multiple routes of expansion out of an African homeland.
On the other hand, while there were apparently direct migrations from North Africa to Iberia and Southern Italy (of people carrying E-V68*, E-V12, E-V22, and E-V65), the majority of E-M78 lineages found in Europe belong to the E-V13 subclade which appears to have entered Europe at some time undetermined from the Near East, where it apparently originated, via the Balkans.
Coming to similar conclusions as the Cruciani and Trombetta team, Battaglia et al. (2008), writing prior to the discovery of E-V68, describe Egypt as "a hub for the distribution of the various geographically localized M78-related sub-clades" and, based on archaeological data, they propose that the point of origin of E-M78 (as opposed to later dispersals from Egypt) may have been in a refugium which "existed on the border of present-day Sudan and Egypt, near Lake Nubia, until the onset of a humid phase around 8500 BC. The northward-moving rainfall belts during this period could have also spurred a rapid migration of Mesolithic foragers northwards in Africa, the Levant and ultimately onwards to Asia Minor and Europe, where they each eventually differentiated into their regionally distinctive branches".
The division of E-V68 into sub-clades such as E-V12, E-V13, etc. has largely been the work of an Italian team including Fulvio Cruciani, Beniamino Trombetta, Rosario Scozzari and others. They started on the basis of STR studies in 2004, and then in 2006 they announced the discoveries of single nucleotide polymorphism (SNP) mutations which could define most of the main branches with better clarity, which was then discussed further in 2007.[2][6][7] These articles were the basis of the updated phylogenies found in Karafet (2008), and ISOGG, which is in turn the basis of the phylogeny given below.
Loosdrecht et al. (2018) analysed genome-wide data from seven ancient Iberomaurusian individuals from the Grotte des Pigeons near Taforalt in eastern Morocco. The fossils were directly dated to between 15,100 and 13,900 calibrated years before present. The scientists found that all the male specimens with sufficient nuclear DNA preservation belonged to the E1b1b1a1 (M78) subclade, with one skeleton bearing the E1b1b1a1b1 parent lineage to E-V13.[8]
Age
Battaglia et al. (2007) estimated that E-M78 (called E1b1b1a1 in that paper) has been in Europe longer than 10,000 years. And more recently, Lacan et al. (2011) found that human remains excavated in a Spanish funeral cave dated to approximately 7000 years ago were in the E-V13 branch of E-M78.
In June 2015, the M78 mutation and the consequent beginning of the E-M78 and E-V68 family trees was dated by Trombetta et al. to approximately 20,300-14,800 years ago.[9]
Family tree
This phylogenetic tree of haplogroup subclades is based on the ISOGG 2019 tree.
| V68 |
E-V68* (E1b1b1a*) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| M78 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Distribution
So far, three individuals who are in E-V68 but not E-M78 have been reported in Sardinia, by Trombetta et al. (2010), when announcing the discovery of V68.
E-M78 is widely distributed in North Africa, Horn of Africa, West Asia (stretching as far as Southern Asia), and Europe.[2][7]
The most basal and rare E-M78* paragroup has been found at its highest frequencies in Egyptians from the Gurna Oasis (5.88%), with lower frequencies also observed in Moroccan Arabs, Sardinians, the Balkans, and Andalusians from Huelva.[2][3][10]
The highest frequencies of all the defined E-M78 sub-clades is primarily found amongst Afroasiatic-speaking populations in the large area stretching from the haplogroup's putative place of origin in Upper Egypt to the Sudan and the Horn of Africa.[6]
Outside of this core area of distribution (North Africa and the Horn of Africa), E-V68 is also observed in other parts of the continent at lower frequencies due to more recent dispersals. It is thus found today in pockets of the African Great Lakes and Southern Africa owing to early Afro-Asiatic-speaking settlers from the Horn region,[9] and as far west as Guinea-Bissau, where its presence has been tentatively attributed to trans-Saharan movements of people from North Africa.[11]
The distribution of E-V68 in Europe is dominated by its E-V13 subclade, except in Iberia. E-V13 has a frequency peak centered in parts of the Balkans (approximately 20% in southern areas; up to almost 50% is some particular places and populations[12][13]) and Italy. It today has lower frequencies toward the western, central and northeastern areas, though E-V13 has been found in a Neolithic burial in Catalonia. This is discussed in more detail below.
| Region | Population | n | E-M78 | E-M78* | E-V12* | E-V13 | E-V22 | E-V32 | E-V65 | Study |
|---|---|---|---|---|---|---|---|---|---|---|
| Europe | Albanians | 55 | 25.46% = (14/55) | 1.82% = (1/55) | 23.64% = (13/55) | [14] | ||||
| Europe | Macedonian Albanians | 64 | 35.94% = (23/64) | 1.56% = (1/64) | 34.38% = (22/64) | [14] | ||||
| Europe | Albanians+ Macedonian Albanians | 55+ 64= 119 | 31.09% = (37/119) | 1.68% = (2/119) | 29.41% = (35/119) | [14] | ||||
| Europe | Kosovar Albanians | 114 | 45.61% = (52/114) | 1.75% = (2/114) | 43.86% = (50/114) | Peričic et al. (2005) | ||||
| Europe | Albanians | 96 | 32.29% = (31/96) | 32.29% = (31/96) | Cruciani et al. (2007) | |||||
| Europe | Kosovar Albanians+ Macedonian Albanians+ Albanians | 119+ 114+ 96= 329 | 36.47% = (120/329) | 1.22% = (4/329) | 35.26% = (116/329) | [14] Peričic et al. (2005) Cruciani et al. (2007) | ||||
| Europe | Macedonian Arumanians | 57 | 29.82 | 29.82 | Peričic et al. (2005) | |||||
| Europe | Serbians | 113 | 20.35 | 1.77 | 18.58 | Peričic et al. (2005) | ||||
| Europe | Croatians | 108 | 5.60 | 5.60 | Peričic et al. (2005) | |||||
| Europe | Crete | 193 | 6.7% = 13/193 | 6.7% = 13/193 | King et al. (2008) | |||||
| Europe | Greeks from Nea Nikomedeia | 57 | 15.8% = 9/57 | 1.8% = 1/57 | 14.0% = 8/57 | King et al. (2008) | ||||
| Europe | Greeks from Sesklo/Dimini | 57 | 38.6% = 22/57 | 3.5% = 2/57 | 35.1% = 20/57 | King et al. (2008) | ||||
| Europe | Greeks from Lerna/Franchthi | 57 | 35.1% = 20/57 | 35.1% = 20/57 | King et al. (2008) | |||||
| Europe | Greeks from Crete+ Greeks from Nea Nikomedeia Greeks from Sesklo/Dimini from Lerna/Franchthi | 193+ 57+ 57+ 57= 364 | 17.58% = 64/364 | 0.82% = 3/364 | 16.76% = 61/364 | King et al. (2008) | ||||
| Europe | Continental Greeks | 147 | 19.05% = 28/147 | 17.69% = 26/147 | 0.68% = 1/147 | 0.68% = 1/147 | Cruciani et al. (2007) | |||
| Europe | Greeks from Crete | 215 | 6.51% = 14/215 | 0.93% = 2/215 | 5.58% = 12/215 | Cruciani et al. (2007) | ||||
| Europe | Greeks from Aegean Islands | 71 | 16.9% = 12/71 | 15.49% = 11/71 | 1.41% = 1/71 | Cruciani et al. (2007) | ||||
| Europe | Continental Greeks Greeks from Crete Greeks from Aegean Islands | 147+ 215+ 71= 433 | 12.47% = 54/433 | 0.46% = 2/433 | 11.32% = 49/433 | 0.46% = 2/433 | 0.23% = 1/433 | Cruciani et al. (2007) | ||
| Europe | Greeks from Crete+ Greeks from Nea Nikomedeia Greeks from Sesklo/Dimini from Lerna/Franchthi Continental Greeks Greeks from Crete Greeks from Aegean Islands | 364+ 433= 797 | 14.81% = 118/797 | 0.38% = 3/797 | 0.25% = 2/797 | 13.8% = 110/797 | 0.25% = 2/797 | 0.13% = 1/797 | King et al. (2008) Cruciani et al. (2007) | |
| Europe | Sicilians | 236 | 11.43 | 1.27 | 5.93 | 3.81 | 0.42 | Di Gaetano et al. (2008) | ||
| Europe | Huelva Andalusians | 167 | 6.59 | 1.20 | 4.19 | 0.60 | 0.60 | Ambrosio et al. (2010) | ||
| Europe | Macedonians | 99 | 18.18 | 17.17 | 1.01 | Cruciani et al. (2007) | ||||
| Europe | Bulgarians | 204 | 16.67 | 0.49 | 16.18 | Cruciani et al. (2007) | ||||
| Europe | Sicilians | 153 | 13.07 | 0.65 | 7.19 | 4.58 | 0.65 | Cruciani et al. (2007) | ||
| Europe | Northern Italians | 94 | 7.45 | 5.32 | 2.13 | Cruciani et al. (2007) | ||||
| Europe | Central Italians | 356 | 7.87 | 0.28 | 5.34 | 1.97 | 0.28 | Cruciani et al. (2007) | ||
| Europe | Southern Italians | 141 | 10.64 | 0.71 | 8.51 | 1.42 | Cruciani et al. (2007) | |||
| Europe | Sardinians | 374 | 3.48 | 0.27 | 0.27 | 1.07 | 0.8 | 1.07 | Cruciani et al. (2007) | |
| Europe | Northern Portuguese | 50 | 4 | 4 | Cruciani et al. (2007) | |||||
| Europe | Southern Portuguese | 49 | 4.08 | 4.08 | Cruciani et al. (2007) | |||||
| Europe | Pasiegos from Cantabria | 56 | Cruciani et al. (2007) | |||||||
| Europe | Asturians | 90 | 10 | 5.56 | 4.44 | Cruciani et al. (2007) | ||||
| Europe | Southern Spaniards | 62 | 3.23 | 3.23 | Cruciani et al. (2007) | |||||
| Europe | Spanish Basques | 55 | Cruciani et al. (2007) | |||||||
| Europe | French Basques | 16 | 6.25 | 6.25 | Cruciani et al. (2007) | |||||
| Europe | French | 225 | 4.44 | 0.44 | 4 | Cruciani et al. (2007) | ||||
| Europe | English | 28 | Cruciani et al. (2007) | |||||||
| Europe | Danish | 35 | 2.86 | 2.86 | Cruciani et al. (2007) | |||||
| Europe | Germans | 77 | 3.9 | 3.9 | Cruciani et al. (2007) | |||||
| Europe | Polish | 40 | 2.5 | 2.5 | Cruciani et al. (2007) | |||||
| Europe | Czechs | 268 | 4.85 | 4.85 | Cruciani et al. (2007) | |||||
| Europe | Slovaks | 24 | 8.33 | 8.33 | Cruciani et al. (2007) | |||||
| Europe | Slovenians | 104 | 2.88 | 2.88 | Cruciani et al. (2007) | |||||
| Europe | Estonians | 74 | 4.05 | 4.05 | Cruciani et al. (2007) | |||||
| Europe | Belarusians | 40 | Cruciani et al. (2007) | |||||||
| Europe | Northern Russians | 82 | 3.66 | 3.66 | Cruciani et al. (2007) | |||||
| Europe | Southern Russians | 92 | 2.17 | 2.17 | Cruciani et al. (2007) | |||||
| Europe | Ukrainians | 11 | 9.09 | 9.09 | Cruciani et al. (2007) | |||||
| Europe | Moldovians | 77 | 7.79 | 7.79 | Cruciani et al. (2007) | |||||
| Europe | Hungarians | 106 | 9.43 | 9.43 | Cruciani et al. (2007) | |||||
| Europe | Rumanians | 265 | 7.55 | 7.17 | 0.38 | Cruciani et al. (2007) | ||||
| Northwestern Africa | Moroccan Arabs | 55 | 40 | 3.64 | 7.27 | 29.09 | Cruciani et al. (2007) | |||
| Northwestern Africa | Asni Berbers | 54 | 3.7 | 3.7 | Cruciani et al. (2007) | |||||
| Northwestern Africa | Bouhria Berbers | 67 | 1.49 | 1.49 | Cruciani et al. (2007) | |||||
| Northwestern Africa | Moyen Atlas Berbers | 69 | 10.14 | 10.14 | Cruciani et al. (2007) | |||||
| Northwestern Africa | Marrakech Berbers | 29 | 6.9 | 3.45 | 3.45 | Cruciani et al. (2007) | ||||
| Northwestern Africa | Moroccan Jews | 50 | 12 | 2 | 2 | 8 | Cruciani et al. (2007) | |||
| Northwestern Africa | Mozabite Berbers | 20 | Cruciani et al. (2007) | |||||||
| Northeastern Africa | Libyan Jews | 25 | 8 | 4 | 4 | Cruciani et al. (2007) | ||||
| Northeastern Africa | Libyan Arabs | 10 | 20 | 20 | Cruciani et al. (2007) | |||||
| Northeastern Africa | Northern Egyptians (Delta) | 72 | 23.61 | 5.56 | 1.39 | 13.89 | 2.78 | Cruciani et al. (2007) | ||
| Northeastern Africa | Egyptian Berbers | 93 | 6.45 | 2.15 | 4.3 | Cruciani et al. (2007) | ||||
| Northeastern Africa | Egyptians from Bahari | 41 | 41.46 | 14.63 | 2.44 | 21.95 | 2.44 | Cruciani et al. (2007) | ||
| Northeastern Africa | Egyptians from Gurna Oasis | 34 | 17.65 | 5.88 | 8.82 | 2.94 | Cruciani et al. (2007) | |||
| Northeastern Africa | Egyptians | 70 | 79 | 79 | Trombetta (2015) | |||||
| Northeastern Africa | Southern Egyptians | 79 | 50.63 | 44.3 | 1.27 | 3.8 | 1.27 | Cruciani et al. (2007) | ||
| Eastern Africa | Dinka | 26 | 15.38 | 3.85 | 11.54 | Hassan et al. (2008) | ||||
| Eastern Africa | Shilluk | 15 | 13.33 | 13.33 | Hassan et al. (2008) | |||||
| Eastern Africa | Nuer | 12 | 16.67 | 16.67 | Hassan et al. (2008) | |||||
| Eastern Africa | Borgu | 26 | 15.38 | 3.85 | 11.54 | Hassan et al. (2008) | ||||
| Eastern Africa | Nuba | 28 | 25 | 3.57 | 3.57 | 7.14 | 10.71 | Hassan et al. (2008) | ||
| Eastern Africa | Masalit | 32 | 71.88 | 3.13 | 15.63 | 53.13 | Hassan et al. (2008) | |||
| Eastern Africa | Fur | 32 | 59.38 | 18.75 | 40.63 | Hassan et al. (2008) | ||||
| Eastern Africa | Nubians | 39 | 15.38 | 12.82 | 2.56 | Hassan et al. (2008) | ||||
| Eastern Africa | Fulani from Sudan | 26 | 34.62 | 30.77 | 3.85 | Hassan et al. (2008) | ||||
| Eastern Africa | Hausa from Sudan | 32 | 3.13 | 3.13 | Hassan et al. (2008) | |||||
| Eastern Africa | Egyptian Copts from Sudan | 33 | 15.15 | 15.15 | Hassan et al. (2008) | |||||
| Eastern Africa | Beja | 42 | 35.71 | 4.76 | 30.95 | Hassan et al. (2008) | ||||
| Eastern Africa | Gaalien | 50 | 18.00 | 6.00 | 6.00 | 6.00 | Hassan et al. (2008) | |||
| Eastern Africa | Meseria | 28 | 14.29 | 3.57 | 10.71 | Hassan et al. (2008) | ||||
| Eastern Africa | Arakien | 24 | 16.67 | 8.33 | 4.17 | 4.17 | Hassan et al. (2008) | |||
| Eastern Africa | Amhara | 34 | 8.82 | 8.82 | Cruciani et al. (2007) | |||||
| Eastern Africa | Ethiopian Jews | 22 | 9.09 | 9.09 | Cruciani et al. (2007) | |||||
| Eastern Africa | Mixed Ethiopians | 12 | 33.33 | 25 | 8.33 | Cruciani et al. (2007) | ||||
| Eastern Africa | Borana/Oromo (Kenya/Ethiopia) | 32 | 40.63 | 40.63 | Cruciani et al. (2007) | |||||
| Eastern Africa | Wolayta | 12 | 16.67 | 8.33 | 8.33 | Cruciani et al. (2007) | ||||
| Eastern Africa | Saho from Eritrea | 94 | 88.3 | 88.3 | Trombetta (2015) | |||||
| Eastern Africa | Somali from Ethiopia | 12 | 33.3 | 8.3 | 25 | Trombetta (2015) | ||||
| Eastern Africa | Somali from Somalia | 5 | 80 | 80 | Trombetta (2015) | |||||
| Eastern Africa | Somali from Kenya | 6 | 80 | 80 | Trombetta (2015) | |||||
| Eastern Africa | Nilotic from Kenya | 18 | 11.11 | 11.11 | Cruciani et al. (2007) | |||||
| Eastern Africa | Bantu from Kenya | 28 | 3.57 | 3.57 | Cruciani et al. (2007) | |||||
| Eastern Africa | Western Africa | 123 | 0.81 | 0.81 | Cruciani et al. (2007) | |||||
| Eastern Africa | Central Africa | 150 | 0.67 | 0.67 | Cruciani et al. (2007) | |||||
| Eastern Africa | Southern Afric | 105 | Cruciani et al. (2007) | |||||||
| Western Asia | Istanbul Turkish | 35 | 8.57 | 2.86 | 5.71 | Cruciani et al. (2007) | ||||
| Western Asia | Southwestern Turkish | 40 | 2.5 | 2.5 | Cruciani et al. (2007) | |||||
| Western Asia | Northeastern Turkish | 41 | Cruciani et al. (2007) | |||||||
| Western Asia | Southeastern Turkish | 24 | 4.17 | 4.17 | Cruciani et al. (2007) | |||||
| Western Asia | Erzurum Turkish | 25 | 4 | 4 | Cruciani et al. (2007) | |||||
| Western Asia | Central Anatolian | 61 | 6.56 | 1.64 | 4.92 | Cruciani et al. (2007) | ||||
| Western Asia | Turkish Cypriots | 46 | 13.04 | 10.87 | 2.17 | Cruciani et al. (2007) | ||||
| Western Asia | Sephardi Turkish | 19 | Cruciani et al. (2007) | |||||||
| Western Asia | Palestinians | 29 | 10.34 | 3.45 | 6.9 | Cruciani et al. (2007) | ||||
| Western Asia | Druze Arabs | 28 | 10.71 | 10.71 | Cruciani et al. (2007) | |||||
| Western Asia | Bedouin | 28 | 3.57 | 3.57 | Cruciani et al. (2007) | |||||
| Western Asia | Syrians | 100 | 2 | 2 | Cruciani et al. (2007) | |||||
| Western Asia | Kurds from Iraq | 20 | Cruciani et al. (2007) | |||||||
| Western Asia | Arabs from United Arab Emirates | 40 | 2.5 | 2.5 | Cruciani et al. (2007) | |||||
| Western Asia | Omanite | 106 | 0.94 | 0.94 | Cruciani et al. (2007) | |||||
| Western Asia | Adygei | 18 | Cruciani et al. (2007) | |||||||
| Western Asia | Azeri | 97 | 2.06 | 2.06 | Cruciani et al. (2007) | |||||
Subclades of M78
Listed here are the main subclades of M78 as of June 2015. Within the E-M78 subclade, Trombetta et al. 2015 allocated most of the former E-M78* chromosomes to three new distinct branches: E-V1083*, E-V1477 and E-V259. The first is a paragroup sister to clades E-V22 and E-V13. The mutation V1477 defines a new basal branch observed only in one northern African sample. Finally, a sister clade of E-V12, defined by V264, includes E-V65 and a new central African lineage defined by V259.[9] The rare M78 subhaplogroup E1b1b1a1-PF2186 has been found at highest frequencies among the Toubou population inhabiting Chad (21%).[15]
- E-M78 (E1b1b1a1) North Africa, Horn of Africa, West Asia, Europe (formerly E1b1b1a).
- E-M78*
- E-V1477 Found in Tunisian Jews.
- E-V1083
- PF2186 Found among Toubou in Lake Chad area.
- E-V1083* Found only in Eritrea (1.1%) and Sardinia (0.3%).
- E-V13 (E1b1b1a1b)
- E-V22
- E-V1129
- E-V12
- E-V12*
- E-V32
- E-V264
- E-V259 Found in Chadic (Afro-Asiatic) speakers from Northern Cameroon.
- E-V65
- E-V12
E-V12
This subclade of E-M78 is the one which appears to have split from the others first (it arose c. 13.7-15.2 kya[16]). According to Cruciani et al. (2007), the E-V12 sublineage likely originated in North Africa.
Undifferentiated E-V12* lineages
Undifferentiated E-V12* lineages (not E-V32 or E-M224, so therefore named "E-V12*") peak in frequency among Southern Egyptians (up to 74.5%).[17] The subclades are also scattered widely in small amounts in both Northern Africa and Europe, but with very little sign in Western Asia, apart from Turkey.[2] These E-V12* lineages were formerly included (along with many E-V22* lineages[Note 1]) in Cruciani et al.'s original (2004) "delta cluster", which he had defined using Y-STR profiles. With the discovery of the defining SNP, Cruciani et al. (2007) reported that V12* was found in its highest concentrations in Egypt, especially Southern Egypt. Hassan et al. (2008) report a significant presence of E-V12* in neighboring Sudan, including 5/33 Copts and 5/39 Nubians. E-V12* made up approximately 20% of the Sudanese E-M78. They propose that the E-V12 and E-V22 sub-clades of E-M78 might have been brought to Sudan from their place of origin in North Africa after the progressive desertification of the Sahara around 6,000–8,000 years ago. Sudden climate change might have forced several Neolithic cultures/people to migrate northward to the Mediterranean and southward to the Sahel and the Nile Valley.[18] The E-V12* paragroup is also observed in Europe (e.g. amongst French Basques) and Eastern Anatolia (e.g. Erzurum Turks).[2]
The non-basal subhaplogroup E1b1b-V12/E3b1a1 has been found at highest frequencies among various Afroasiatic-speaking populations in eastern Africa, including Garreh (74.1%), Gabra (58.6%), Wata (55.6%), Borana (50.0%), Sanye (41.7%), Beja (33.3%) and Rendille (29.0%).[19]
Sub-clades of E-V12
E-M224
E-M224 has been found in Israel among Yemeni population (5%) and appears to be a minor subclade.
Its discovery was announced in Underhill et al. (2001) and Shen et al. (2004) found 1 out of their 20 Yemeni Israelis they tested. Cruciani et al. (2006) called M224 "rare and rather uninformative" and they found no exemplars.
E-V32
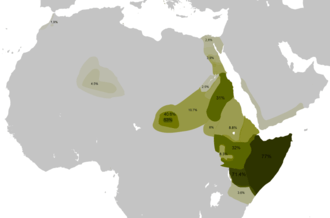
Cruciani et al. (2007) suggest that this subclade of E-V12 originated in North Africa, and then subsequently expanded further south into the Horn of Africa, where it is now prevalent.[Note 2] Before the discovery of V32, Cruciani et al. (2004) referred to the same lineages as the "gamma cluster", which was estimated to have arisen about 8,500 years ago. They stated that "the highest frequencies in the three Cushitic-speaking groups: the Borana from Kenya (71.4%), the Oromo from Ethiopia (32.0%), and the Somali (52.2%). Outside of eastern Africa, it was found only in two subjects from Egypt (3.6%) and in one Arab from Morocco". Sanchez et al. (2005) found it extremely prominent in Somali men and stated that "the male Somali population is a branch of the Horn African population – closely related to the Oromos in Ethiopia and North Kenya (Boranas)" and that their gamma cluster lineages "probably were introduced into the Somali population 4000–5000 years ago". More recently, Tillmar et al. (2009) typed 147 males from Somalia for 12 Y-STR loci, and observed that 77% (113/147) had typical E-V32 haplotypes. This is currently the highest frequency of E-V32 found in any single sample population. Similarly, Hassan et al. (2008) in their study observed this to be the most common of the sub-clades of E-M78 found in Sudan, especially among the Beja, Masalit and Fur. The Beja, like Somalis and Oromos, speak an Afro-Asiatic language and live along the "corridor" from the Horn of Africa to Egypt. Hassan et al. (2008) interpret this as reinforcing the "strong correlation between linguistic and genetic diversity" and signs of relatedness between the Beja and the peoples of the Horn of Africa such as the Amhara and Oromo. On the other hand, the Masalit and Fur live in Darfur and speak a Nilo-Saharan language. The authors observed in their study that "the Masalit possesses by far the highest frequency of the E-M78 and of the E-V32 haplogroup", which they believe suggests "either a recent bottleneck in the population or a proximity to the origin of the haplogroup." However, More recently, Tillmar et al. (2009) typed 147 males from Somalia for 12 Y-STR loci, and observed that 77% (113/147) had typical E-V32 haplotypes. This is the highest frequency of E-V32 found in any single sample population.
The STR data from Cruciani et al. (2007) concerning E-V12 can be summarized as follows.
| Haplotype | description | YCAIIa | YCAIIb | DYS413a | DYS413b | DYS19 | DYS391 | DYS393 | DYS439 | DYS460 | DYS461 | A10 |
| E-V12* | modal | 19 | 22 | 22 | 22 | 13 | 10 | 13 | 11 | 11 | 9 | 13 |
| min | 18 | 21 | 20 | 21 | 11 | 10 | 12 | 11 | 8 | 8 | 11 | |
| max | 19 | 22 | 22 | 23 | 15 | 12 | 14 | 13 | 12 | 10 | 14 | |
| number | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | 40 | |
| E-V32 | modal | 19 | 21 | 22 | 23 | 11 | 10 | 13 | 12 | 10 | 10 | 13 |
| min | 19 | 19 | 20 | 21 | 11 | 9 | 12 | 11 | 9 | 10 | 11 | |
| max | 20 | 22 | 22 | 24 | 11 | 11 | 13 | 13 | 12 | 11 | 14 | |
| number | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | 35 | |
| All E-V12 | modal | 19 | 22 | 22 | 23 | 11 | 10 | 13 | 11 | 11 | 10 | 13 |
| min | 18 | 19 | 20 | 21 | 11 | 9 | 12 | 11 | 8 | 8 | 11 | |
| max | 20 | 22 | 22 | 24 | 15 | 12 | 14 | 13 | 12 | 11 | 14 | |
| number | 75 | 75 | 75 | 75 | 75 | 75 | 75 | 75 | 75 | 75 | 75 | |
E-V13
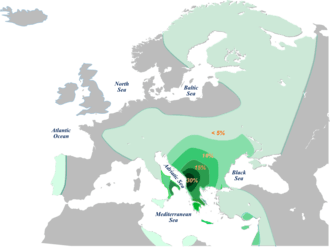
The E-V13 clade is equivalent to the "alpha cluster" of E-M78 reported in Cruciani et al. (2004), and was first defined by the SNP V13 in Cruciani et al. (2006). Another SNP is known for this clade, V36, reported in Cruciani et al. (2007). All known positive tests for V13 are also positive for V36. So E-V13 is currently considered "phylogenetically equivalent" to E-V36.
Haplogroup E-V13 is the only lineage that reaches the highest frequencies out of Africa. In fact, it represents about 85% of the European E-M78 chromosomes with a clinal pattern of frequency distribution from the southern Balkan peninsula (19.6%) to western Europe (2.5%). The same haplogroup is also present at lower frequencies in Anatolia (3.8%), the Near East (2.0%), and the Caucasus (1.8%). In Africa, haplogroup E-V13 is rare, being observed only in northern Africa at a low frequency (0.9%).
According to some authors E-V13 appears to have originated in Greece or the southern Balkans and its presence in the rest of the Mediterranean is likely a consequence of Greek colonization.[20][21][22] Within Europe, E-V13 is especially common in the Balkans and some parts of Italy. In different studies, particularly high frequencies have been observed in Kosovo Albanians (45.6%[23]), Macedonian Albanians (34.4%[14]), Albanians (32.29%Cruciani et al. (2007)) , and in some parts of Greece (ca. 35%[24]).[25] More generally, high frequencies have also been found in other areas of Greece, and amongst Bulgarians, Romanians, Macedonians and Serbs.[6][13][26][27]
Within Italy, frequencies tend to be higher in Southern Italy,[2] with particularly high results sometimes seen in particular areas; for example, in Santa Ninfa and Piazza Armerina in Sicily.[28] High frequencies appear to exist also in some northern areas[Note 3] for example around Venice,[Note 4] Genoa[29] and Rimini,[30] as well as on the island of Corsica [31] and the region of Provence in south France,[22] and is also found in scattered and small amounts in Libyan Jews and Egypt, but this is most likely a result of migration from Europe or the Near East.[2]
Among ancient specimens, Loosdrecht et al. (2018) found one E-M78-carrying fossil at the Grotte des Pigeons near Taforalt in eastern Morocco. The skeleton has been directly dated to between 15,100 and 13,900 calibrated years before present.[8]
E-V13 and ancient migrations
The apparent movement of E-M78 lineages from the Near East to Europe, and their subsequent rapid expansion, make its E-V13 subclade a particularly interesting subject for speculation about ancient human migrations.
It was concluded that northeastern Africa, rather than eastern Africa, was where the E-M78 chromosomes began dispersing to other regions.[32] The most plausible scenario is that E-V13 originated in Western Asia.[33] A hypothesis is that E-M78 carriers devoid of V13 mutation left Africa and that the coalescene occurred later in the Near East/Anatolia.[33] Data suggests that Western Asian carriers of V13 expanded in Europe at earliest 5300 years ago.[33] The TMRCA of European V13 is 4700–4000 years ago.[33] Phylogenetic analysis suggest that the European v13 spread through Europe from the Balkans in a "rapid demographic expansion".[33]
Before then, the SNP mutation, V13 apparently first arose in West Asia around 10 thousand years ago, and although not widespread there, it is for example found in high levels (>10% of the male population) in Turkish Cypriot and Druze Arab lineages.[2] The Druze are considered a genetically isolated community, and are therefore of particular interest.[34] The STR DNA signature of some of the E-V13 men amongst them was actually originally classified in the delta cluster in Cruciani et al. (2004). This means that Druze E-V13 clustered together with most E-V12 and E-V22, and not with European E-V13, which was mostly in the alpha cluster.
| haplotype | description | YCAIIa | YCAIIb | DYS413a | DYS413b | DYS19 | DYS391 | DYS393 | DYS439 | DYS460 | DYS461 | A10 |
| All E-V13 | modal | 19 | 21 | 23 | 24 | 13 | 10 | 13 | 12 | 9 | 10 | 13 |
| Druze V13 | 1 | 19 | 21 | 23 | 23 | 13 | 10 | 13 | 13 | 11 | 9 | 12 |
| Druze V13 | 2 | 19 | 21 | 23 | 23 | 13 | 10 | 13 | 13 | 11 | 9 | 13 |
| All E-V22 | modal | 19 | 22 | 22 | 23 | 14 | 10 | 13 | 12 | 11 | 10 | 12 |
| All E-V12* | modal | 19 | 22 | 22 | 22 | 13 | 10 | 13 | 11 | 11 | 9 | 13 |
Early migration from the Middle East to Europe
The distribution and diversity of V13 are often thought to represent the introduction of early farming technologies, during the Neolithic expansion, into Europe by way of the Balkans.[12] The haplogroup J2b (J-M12) has also frequently been discussed in connection with V13, as a haplogroup with a seemingly very similar distribution and pre-history.[3][6][12] (There is no consensus regarding the circumstances or timing of its evolution.)
Cruciani et al. (2007) says there were at least four major demographic events which have been envisioned for this geographic area:
- The "post-Last Glacial Maximum expansion (about 20 kya)"
- The "Younger Dryas-Holocene reexpansion (about 12 kya)"
- The "population growth associated with the introduction of agricultural practices (about 8 kya)"
- The "development of Bronze technology (about 5kya)"
The last two seem within the timespan possible for V13 given its STR age of arise putatively in the Middle East. In favor of the agricultural connection, human remains excavated in a Spanish funeral cave dating from approximately 7000 years ago were shown to be in this haplogroup.[35]
However, earlier entry into Europe is also possible. Battaglia et al. (2008), for example, propose that the E-M78* lineage ancestral to all modern E-V13 men moved rapidly out of a Southern Egyptian homeland, in the wetter conditions of the early Holocene; arrived in the Balkans with only Mesolithic technologies and then only subsequently integrated with Neolithic cultures which arrived later in the Balkans.
E-V13 is in any case often described in population genetics as one of the components of the European genetic composition which shows a relatively recent link of populations from the Middle East, entering Europe and presumably associated with bringing new technologies.[36][37][38] As such, it is also sometimes remarked that it is a relatively recent genetic movement out of Africa into Eurasia, and has been described as "a signal for a separate late-Pleistocene migration from Africa to Europe over the Sinai ... which is not manifested in mtDNA haplogroup distributions".[39]
After its initial entry in Europe, there was then a dispersal from the Balkans into the rest of Europe. Also for this movement, a wide range of possibilities exists. Battaglia et al. (2008) suggest that the E-V13 subclade of E-M78 originated in situ in Europe, and propose that the first major dispersal of E-V13 from the Balkans may have been in the direction of the Adriatic Sea with the Neolithic Impressed Ware culture often referred to as Impressa or Cardial. The above-mentioned find of archaic E-V13 in Spain supports this suggestion.
In contrast, Cruciani et al. (2007) suggest that the movement out of the Balkans may have been more recent than 5300 years ago. The authors suggest that for the most part, modern E-V13 descends from a population which remained in the Balkans until the Balkan Bronze Age. They consider that "the dispersion of the E-V13 and J-M12 haplogroups seems to have mainly followed the river waterways connecting the southern Balkans to north-central Europe". Peričic et al. (2005) propose the Vardar-Morava-Danube rivers as a possible route of Neolithic dispersal into central Europe. Bird (2007) proposes a still more recent dispersal out of the Balkans, around the time of the Roman empire.
According to Lacan et al. (2011), Neolithic skeletons (~7,000 years old) that were excavated from the Avellaner cave in Catalonia, northeastern Spain included a male specimen, which carried haplogroup E1b1b. This fossil belonged to the E1b1b1a1b (V13) subclade, and possessed identical haplotypes as found in modern European individuals (five Albanians, two Provence French, two Corsicans, two Bosnians, one Italian, one Sicilian, and one Greek). The presence of this haplogroup in Neolithic Spain suggests that it is associated with the Neolithic agricultural package. The ancient farmer also bore the U5 mtDNA clade, an early European maternal haplogroup. His autosomal STR markers were likewise most typical of Europe. Additionally, the specimen was homozygous C/C for the LP-13910-C/T lactase persistence SNP, indicating that he was lactose intolerant.
Greek soldiers in Pakistan
Both E-M78 and J-M12 have also been used in studies seeking to find evidence of a remaining Greek presence in Afghanistan and Pakistan, going back to the time of Alexander the Great.
An extensive analysis of Y diversity within Greeks and three Pakistani populations – the Burusho, Kalash and Pathan – who claim descent from Greek soldiers allowed us to compare Y lineages within these populations and re-evaluate their suggested Greek origins. This study as a whole seems to exclude a large Greek contribution to any Pakistani population, confirming previous observations. However, it provides strong evidence in support of the Greek origins for a small proportion of Pathans, as demonstrated by the clade E network and the low pairwise genetic distances between these two populations.
This study however tested only for M78, and not V13, the typical type of M78 from the Balkans. More recent and detailed analyses of E-V13 in this region have however concluded that this hypothesis is incorrect, and that the variants found there are not the types typical of the Balkans.[40] Instead "Afghanistan's lineages are correlated with Middle Easterners and Iranians but not with populations from the Balkans"[41]
Ancient Britain
Significant frequencies of E-V13 have also been observed in towns in Wales, around Chester (ancient Deva Victrix) in England, and Scotland. The old trading town of Abergele on the northern coast of Wales in particular showed 7 out of 18 local people tested were in this lineage (approximately 40%), as reported in Weale et al. (2002).
Some scholars (e.g. Steven Bird (2007) have attributed the presence of E-V13 in Great Britain, especially in areas of high frequency, to Roman settlement during the 1st through 4th centuries CE. The Roman Army including men of Balkan ancestry, including Thracians, Illyrians and Dacians. In particular, Steven Bird proposes a connection to a modern region encompassing Kosovo, southern Serbia, northern Macedonia, and extreme northwestern Bulgaria – a region corresponding to the Roman province of Moesia Superior, which was identified by Peričic et al. (2005) as harboring the highest frequency worldwide of this subclade.[Note 5]
It is also notable that E-V13 appears to be absent in modern central England, especially the West Midlands and South Midlands.[Note 6] Bird (2007) notes that the collective genetic profile of the English Midlands is similar to that of the Dutch province of Friesland, which was not colonised by Rome, but was, like England, subject to Anglo-Saxon settlement. The so-called "E3b hole" in Central England, according to Steven Bird, may reflect a population replacement – of Romano-British people by Anglo-Saxons.[Note 7] Thomas et al. (2006) raises the possibility of "apartheid"-type, elite dominance social structures in Anglo-Saxon England. Bird (2007) concurs: "The 'E3b hole' suggests that either (a) a massive displacement of the ... Romano-British population by invasion or, (b) the substantial genetic replacement of Romano-British Y-DNA through an elite dominance ("apartheid") model... Regardless of the mechanism, the Central England region ... with its lack of E3b haplotypes, is the area having the most "striking similarity in the distribution of Y-chromosomes" with Friesland."
Sub-clades of E-V13
Although most E-V13 individuals do not show any known downstream SNP mutations, and are therefore categorized as E-V13* there are several recognized sub-clades, all of which may be very small. These are one of two cases where Karafet et al. (2008) remarked that at the time of that article, it was not certain that the two clades were truly separate ("the positions of these mutations have not been resolved because of a lack of a DNA sample containing the derived state at V27").
- E-V27. Defined by V27. Cruciani et al. (2007) found one case in Sicily.
- E-P65. Defined by P65.
- E-L17. Defined by L17.
- E-L143. Defined by L143.
- E-M35.2. Defined by M35.2.
- E-L241. Defined by L241.
- E-L250. Defined by L250, L251, and L252.
E-V22
This clade comprises most of those classified in the "delta cluster" of Cruciani et al. (2004). Cruciani et al. (2006) later noted that "E-V22 and E-V12* chromosomes are intermingled and not clearly differentiated by their microsatellite haplotypes".
This subclade of E-M78 is "relatively common"[2] in the Horn of Africa and Egypt, with higher microsatellite variance (0.35 vs. 0.46, respectively) in Egypt. In the article announcing this first information, Cruciani et al. (2007) described it as uncommon in Western Asia and they proposed Northeast Africa (Libya/Egypt) as this subclade's likely place of origin.
The highest frequency of E-V22 has thus far been observed in the Cushitic-speaking Saho population of central Eritrea at a rate of 88%.[9] The Saho are known to be organized in strict patrilineal and patrilocal clans. It has been hypothesized that this kind of social structure can explain patterns of variability characterized by low Y-chromosome diversity within groups and large difference between groups.[42]
Hassan et al. (2008) also reported a significant presence in neighboring Sudan, making up about 30% of the diverse range of the country's E-M78 lineages in their study, including 8 out of 26 Fulani (about 31%), a widely-dispersed pastoral people.[Note 8] E-V22 was also present in much smaller frequencies amongst the Shilluk (2 of 15 samples, 13%) and Dinka (3 of 26, 11.5%) Nilotes of Southern Sudan. Hassan et al. suggest that E-V22, like E-V12, might have entered Sudan from North Africa "after the progressive desertification of the Sahara around 6,000–8,000 years ago". They add that the gene flow to Sudan "is not only recent (Holocene onward) but also largely of focal nature", and that "most speakers of Nilo-Saharan languages, the major linguistic family spoken in the country, show very little evidence of gene flow and demonstrate low migration rate, with exception of the Nubians, who appear to have sustained considerable gene flow from Asia and Europe together with the Beja."
Other frequencies reported by Cruciani et al. (2007) include Asturians (4.44% out of 90 people), Sicilians (4.58% out of 153 people), Moroccan Arabs (7.27%, 55 people), Moroccan Jews (8%, 50 people), Istanbul Turkish (5.71% out of 35 people), and Palestinians (6.9% out of 29 people). Cadenas et al. (2007) found a 6.7% presence in the UAE.
Sub-clades of E-V22
There are two recognized sub-clades, which are apparently separate, although Karafet (2008) remarked that at the time of that article, "the positions of these mutations have not been resolved because of a lack of a DNA sample containing the derived state at [...] V19".
- E-M148 Defined by M148. Underhill et al. (2000) found 1 example in the Indian subcontinent. Cruciani et al. (2006) calls M148 "rare and rather uninformative".
- E-V19 Defined by V19. Cruciani et al. (2007) found 2 exemplars in Sardinia.
E-V65
This subclade, equivalent to the previously classified "beta cluster", is found in high levels in the Maghreb regions of far northern Africa. Cruciani et al. (2007) report levels of about 20% amongst Libyan Arab lineages, and about 30% amongst Moroccan Arabs. It appears to be less common amongst Berbers, but still present in levels of >10%. The authors suggest a North African origin for this lineage. In Europe, only a few individuals were found in Italy and Greece. The results from the article can be summarized as follows...
| E-V65 | YCAIIa | YCAIIb | DYS413a | DYS413b | DYS19 | DYS391 | DYS393 | DYS439 | DYS460 | DYS461 | A10 |
| modal | 19 | 21 | 21 | 23 | 13 | 10 | 13 | 10 | 10 | 11 | 13 |
| min | 19 | 20 | 20 | 22 | 11 | 10 | 13 | 10 | 9 | 9 | 12 |
| max | 21 | 21 | 22 | 23 | 14 | 11 | 14 | 11 | 11 | 12 | 13 |
| number | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 | 38 |
Capelli et al. (2009) studied the beta cluster in Europe. They found small amounts in Southern Italy, but also traces in Cantabria, Portugal and Galicia, with Cantabria having the highest level in Europe in their study, at 3.1% (5 out of 161 people).
E-M521
This subclade's discovery was announced in Battaglia et al. (2008) They found 2 out of 92 Greeks to have this mutation.
Phylogenetics
Phylogenetic history
Prior to 2002, there were in academic literature at least seven naming systems for the Y-Chromosome Phylogenetic tree. This led to considerable confusion. In 2002, the major research groups came together and formed the Y-Chromosome Consortium (YCC). They published a joint paper that created a single new tree that all agreed to use. Later, a group of citizen scientists with an interest in population genetics and genetic genealogy formed a working group to create an amateur tree aiming at being above all timely. The table below brings together all of these works at the point of the landmark 2002 YCC Tree. This allows a researcher reviewing older published literature to quickly move between nomenclatures.
| YCC 2002/2008 (Shorthand) | (α) | (β) | (γ) | (δ) | (ε) | (ζ) | (η) | YCC 2002 (Longhand) | YCC 2005 (Longhand) | YCC 2008 (Longhand) | YCC 2010r (Longhand) | ISOGG 2006 | ISOGG 2007 | ISOGG 2008 | ISOGG 2009 | ISOGG 2010 | ISOGG 2011 | ISOGG 2012 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E-P29 | 21 | III | 3A | 13 | Eu3 | H2 | B | E* | E | E | E | E | E | E | E | E | E | E |
| E-M33 | 21 | III | 3A | 13 | Eu3 | H2 | B | E1* | E1 | E1a | E1a | E1 | E1 | E1a | E1a | E1a | E1a | E1a |
| E-M44 | 21 | III | 3A | 13 | Eu3 | H2 | B | E1a | E1a | E1a1 | E1a1 | E1a | E1a | E1a1 | E1a1 | E1a1 | E1a1 | E1a1 |
| E-M75 | 21 | III | 3A | 13 | Eu3 | H2 | B | E2a | E2 | E2 | E2 | E2 | E2 | E2 | E2 | E2 | E2 | E2 |
| E-M54 | 21 | III | 3A | 13 | Eu3 | H2 | B | E2b | E2b | E2b | E2b1 | - | - | - | - | - | - | - |
| E-P2 | 25 | III | 4 | 14 | Eu3 | H2 | B | E3* | E3 | E1b | E1b1 | E3 | E3 | E1b1 | E1b1 | E1b1 | E1b1 | E1b1 |
| E-M2 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a* | E3a | E1b1 | E1b1a | E3a | E3a | E1b1a | E1b1a | E1b1a | E1b1a1 | E1b1a1 |
| E-M58 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a1 | E3a1 | E1b1a1 | E1b1a1 | E3a1 | E3a1 | E1b1a1 | E1b1a1 | E1b1a1 | E1b1a1a1a | E1b1a1a1a |
| E-M116.2 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a2 | E3a2 | E1b1a2 | E1b1a2 | E3a2 | E3a2 | E1b1a2 | E1b1a2 | E1ba12 | removed | removed |
| E-M149 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a3 | E3a3 | E1b1a3 | E1b1a3 | E3a3 | E3a3 | E1b1a3 | E1b1a3 | E1b1a3 | E1b1a1a1c | E1b1a1a1c |
| E-M154 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a4 | E3a4 | E1b1a4 | E1b1a4 | E3a4 | E3a4 | E1b1a4 | E1b1a4 | E1b1a4 | E1b1a1a1g1c | E1b1a1a1g1c |
| E-M155 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a5 | E3a5 | E1b1a5 | E1b1a5 | E3a5 | E3a5 | E1b1a5 | E1b1a5 | E1b1a5 | E1b1a1a1d | E1b1a1a1d |
| E-M10 | 8 | III | 5 | 15 | Eu2 | H2 | B | E3a6 | E3a6 | E1b1a6 | E1b1a6 | E3a6 | E3a6 | E1b1a6 | E1b1a6 | E1b1a6 | E1b1a1a1e | E1b1a1a1e |
| E-M35 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b* | E3b | E1b1b1 | E1b1b1 | E3b1 | E3b1 | E1b1b1 | E1b1b1 | E1b1b1 | removed | removed |
| E-M78 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b1* | E3b1 | E1b1b1a | E1b1b1a1 | E3b1a | E3b1a | E1b1b1a | E1b1b1a | E1b1b1a | E1b1b1a1 | E1b1b1a1 |
| E-M148 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b1a | E3b1a | E1b1b1a3a | E1b1b1a1c1 | E3b1a3a | E3b1a3a | E1b1b1a3a | E1b1b1a3a | E1b1b1a3a | E1b1b1a1c1 | E1b1b1a1c1 |
| E-M81 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b2* | E3b2 | E1b1b1b | E1b1b1b1 | E3b1b | E3b1b | E1b1b1b | E1b1b1b | E1b1b1b | E1b1b1b1 | E1b1b1b1a |
| E-M107 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b2a | E3b2a | E1b1b1b1 | E1b1b1b1a | E3b1b1 | E3b1b1 | E1b1b1b1 | E1b1b1b1 | E1b1b1b1 | E1b1b1b1a | E1b1b1b1a1 |
| E-M165 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b2b | E3b2b | E1b1b1b2 | E1b1b1b1b1 | E3b1b2 | E3b1b2 | E1b1b1b2a | E1b1b1b2a | E1b1b1b2a | E1b1b1b2a | E1b1b1b1a2a |
| E-M123 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b3* | E3b3 | E1b1b1c | E1b1b1c | E3b1c | E3b1c | E1b1b1c | E1b1b1c | E1b1b1c | E1b1b1c | E1b1b1b2a |
| E-M34 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3b3a* | E3b3a | E1b1b1c1 | E1b1b1c1 | E3b1c1 | E3b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1c1 | E1b1b1b2a1 |
| E-M136 | 25 | III | 4 | 14 | Eu4 | H2 | B | E3ba1 | E3b3a1 | E1b1b1c1a | E1b1b1c1a1 | E3b1c1a | E3b1c1a | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1c1a1 | E1b1b1b2a1a1 |
Research publications
The following research teams per their publications were represented in the creation of the YCC tree.
See also
| Wikiquote has quotations related to: Haplogroup E-V68 |
Genetics
- African admixture in Europe
- Genetic genealogy
- Haplogroup D (Y-DNA)
- Haplogroup DE (Y-DNA)
- Haplogroup
- Haplotype
- Human Y-chromosome DNA haplogroup
- Molecular phylogenetics
- Paragroup
- Subclade
- Y-chromosome haplogroups in populations of the world
- Y-DNA haplogroups by ethnic group
- Y-DNA haplogroups in populations of Sub-Saharan Africa
Y-DNA E subclades
- Haplogroup E-L485 (Y-DNA)
- Haplogroup E-M180 (Y-DNA)
- Haplogroup E-M33 (Y-DNA)
- Haplogroup E-M96 (Y-DNA)
- Haplogroup E-P147 (Y-DNA)
- Haplogroup E-P177 (Y-DNA)
- Haplogroup E-P2 (Y-DNA)
- Haplogroup E-V12 (Y-DNA)
- Haplogroup E-V13 (Y-DNA)
- Haplogroup E-V22 (Y-DNA)
- Haplogroup E-V65 (Y-DNA)
- Haplogroup E-V38 (Y-DNA)
- Haplogroup E-M215 (Y-DNA)
- Haplogroup E-M123 (Y-DNA)
- Haplogroup E-M75 (Y-DNA)
- Haplogroup E-V68 (Y-DNA)
- Haplogroup E-Z820 (Y-DNA)
- Haplogroup E-Z827 (Y-DNA)
- Haplogroup E-M521 (Y-DNA)
Y-DNA backbone tree
| Phylogenetic tree of human Y-chromosome DNA haplogroups [χ 1][χ 2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-chromosomal Adam" | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| G | HIJK | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | H | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| I | J | LT [χ 5] | K2 [χ 6] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| L | T | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| K-M2313 [χ 10] | K2b1 [χ 11] | P [χ 12] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| NO | S [χ 13] | M [χ 14] | P1 | P2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| N | O | Q | R | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Notes
- Cruciani et al. (2004): "E-V22 and E-V12* chromosomes are intermingled and not clearly differentiated by their microsatellite haplotypes". In Cruciani et al. (2007) the same authors show that a branch of E-V13 found amongst the Druze Arabs is also in the delta cluster. (Contrast the data tables of Cruciani et al. (2007) and Cruciani et al. (2004).)
- Cruciani et al. (2007): Fig. 2/C
- Genetic surveys do not all test the same markers.
- Scozzari et al. 2001. See clade 25.1. The same data set was later used in Cruciani et al. (2004) and Cruciani et al. (2007).
- Doubts about this line of reasoning have been expressed because: (a.) new data appearing in King et al. (2008) indicates that there were also high concentrations of E-V13 in Greece and (b.) the data in Peričic et al. (2005) show that the area with the highest frequency does not have the highest diversity, implying that V13 arrived there more recently than in Greece.
- Bird uses three sources: Weale et al. (2002), Capelli et al. (2003) and Sykes (2006). Neither Capelli nor Weale have data from the area in the English Midlands where Bird suggests that there is a lack of E1b1b [editor E-M243]. In 2006 Bird mentioned that there were 193 Central English haplotypes in Sykes.
- However, in the E3b distribution maps published in Bird's own paper – the Norfolk area is shown as having a high percentage of E3b. Norfolk is part of the epicentre of the supposed Anglian invasion.
- Rosa et al. (2007) in a study of Guinea Bissau, showed that the Fulani there are about 10% E-M78. Note that this study did not test specifically for V12 or V22, so the E-M78 may have a different exact breakdown of diversity as well as a lower frequency.
- "E-L539 YTree".
- Cruciani et al. (2007)
- Battaglia et al. (2008)
- ISOGG, Copyright 2016 by. "ISOGG 2017 Y-DNA Haplogroup E". isogg.org. Retrieved 2019-01-07.
- Cruciani et al. (2007) Table 1
- Cruciani et al. (2004)
- Cruciani et al. (2006)
- Loosdrecht; et al. (2018). "Pleistocene North African genomes link Near Eastern and sub-Saharan African human populations". Science. 360: eaar8380. Bibcode:2018Sci...360..548V. doi:10.1126/science.aar8380. PMID 29545507.
- Trombetta (2015)
- Ambrosio et al. (2010)
- Rosa et al. (2007)
- Semino et al. (2004)
- Peričic (2005)
- Battaglia et al. 2008.
- Haber, Marc; et al. (2016). "Chad Genetic Diversity Reveals an African History Marked by Multiple Holocene Eurasian Migrations". American Journal of Human Genetics. 99 (6): 1316–1324. doi:10.1016/j.ajhg.2016.10.012. PMC 5142112. PMID 27889059. - Y-chromosomal haplogroup frequencies on Table S.4
- See Figure 1.
- Beniamino Trombetta (2015). "Phylogeographic refinement and large scale genotyping of human Y chromosome haplogroup E provide new insights into the dispersal of early pastoralists in the African continent". Genome Biology and Evolution. 7 (7): 1940–1950. doi:10.1093/gbe/evv118. PMC 4524485. PMID 26108492.
- Hassan et al. (2008)
- Hirbo, Jibril Boru. "Complex Genetic History of East African Human Populations" (PDF). University of Maryland, College Park. Retrieved 13 July 2017.
- Sarno, Stefania; et al. (2014). "An Ancient Mediterranean Melting Pot: Investigating the Uniparental Genetic Structure and Population History of Sicily and Southern Italy". PLOS ONE. 9 (4): e96074. doi:10.1371/journal.pone.0096074. PMC 4005757. PMID 24788788.
- Primorac, Dragan; et al. (2011). "Croatian genetic heritage: Y-chromosome story". Croat Med J. 52 (3): 225–234. doi:10.3325/cmj.2011.52.225. PMC 3118711. PMID 21674820.
- King, Roy J.; et al. (2011). "The coming of the Greeks to Provence and Corsica: Y-chromosome models of archaic Greek colonization of the western Mediterranean". BMC Evolutionary Biology. 11: 69. doi:10.1186/1471-2148-11-69. PMC 3068964. PMID 21401952.
- Peričic et al. 2005.
- King et al. 2008.
- Semino et al. (2004) suggest that there might be levels of E-M78 in the Peloponnese above 40%. They found 17 out of 36 there (47%), but justified drawing conclusions from this small sample by referring also to Di Giacomo et al. (2003).
- Rosser et al. 2000
- King et al. (2008)
- Di Gaetano et al. (2008)
- Di Giacomo et al. (2003)
- Pelotti et al. 2007
- Francalacci et al. 2003
- Cruciani et al. 2007, "Locating the Origin of Haplogroup E-M78".
- Cruciani et al. 2007, "The Haplogroup E-V13: Migrations and Demographic Expansions in Western Eurasia".
- Shlush et al. (2008)
- Lacan et al. (2011)
- Semino et al. (2000)
- King and Underhill (2002)
- Underhill (2002)
- Underhill and Kivisild (2007)
- Lacau et al. (2012)
- Haber et al. (2012)
- Iacovacci, Giuseppe; et al. (2017). "Forensic data and microvariant sequence characterization of 27 Y-STR loci analyzed in four Eastern African countries". Forensic Science International: Genetics. 27: 123–131. doi:10.1016/j.fsigen.2016.12.015. PMID 28068531. Retrieved 23 September 2018.
References
- Ambrosio, B; Dugoujon, JM; Hernández, C; De La Fuente, D; González-Martín, A; Fortes-Lima, CA; Novelletto, A; Rodríguez, JN; Calderón, R; et al. (2010), "The Andalusian population from Huelva reveals a high diversification of Y-DNA paternal lineages from haplogroup E: Identifying human male movements within the Mediterranean space", Annals of Human Biology, 37 (1): 86–107, doi:10.3109/03014460903229155, PMID 19939195
- Adams, Susan M; Bosch, Elena; Balaresque, Patricia L.; Ballereau, Stéphane J.; Lee, Andrew C.; Arroyo, Eduardo; López-Parra, Ana M.; Aler, Mercedes; et al. (2008), "The Genetic Legacy of Religious Diversity and Intolerance: Paternal Lineages of Christians, Jews, and Muslims in the Iberian Peninsula", The American Journal of Human Genetics, 83 (6): 725–36, doi:10.1016/j.ajhg.2008.11.007, PMC 2668061, PMID 19061982
- Alvarez; Santos, Cristina; Montiel, Rafael; Caeiro, Blazquez; Baali, Abdellatif; Dugoujon, Jean-Michel; Aluja, Maria Pilar (2009), "Y-chromosome variation in South Iberia: Insights into the North African contribution", American Journal of Human Biology, 21 (3): 407–409, doi:10.1002/ajhb.20888, PMID 19213004
- Arredi, B; Poloni, E; Paracchini, S; Zerjal, T; Fathallah, D; Makrelouf, M; Pascali, V; Novelletto, A; Tylersmith, C (2004), "A Predominantly Neolithic Origin for Y-Chromosomal DNA Variation in North Africa", American Journal of Human Genetics, 75 (2): 338–345, doi:10.1086/423147, PMC 1216069, PMID 15202071
- Battaglia, Vincenza; Fornarino, Simona; Al-Zahery, Nadia; Olivieri, Anna; Pala, Maria; Myres, Natalie M; King, Roy J; Rootsi, Siiri; et al. (2008), "Y-chromosomal evidence of the cultural diffusion of agriculture in southeast Europe", European Journal of Human Genetics, 17 (6): 820–830, doi:10.1038/ejhg.2008.249, PMC 2947100, PMID 19107149
- Behar, Doron M.; Thomas, Mark G.; Skorecki, Karl; Hammer, Michael F.; Bulygina, Ekaterina; Rosengarten, Dror; Jones, Abigail L.; Held, Karen; et al. (October 2003), "Multiple Origins of Ashkenazi Levites: Y Chromosome Evidence for Both Near Eastern and European Ancestries", Am. J. Hum. Genet., 73 (4), pp. 768–779, doi:10.1086/378506, PMC 1180600, PMID 13680527. Also at http://www.ucl.ac.uk/tcga/tcgapdf/Behar-AJHG-03.pdf and https://web.archive.org/web/20090304100321/http://www.familytreedna.com/pdf/400971.pdf
- Behar; Garrigan; Kaplan; Mobasher; Rosengarten (November 2004), "Contrasting patterns of Y chromosome variation in Ashkenazi Jewish and host non-Jewish European populations" (PDF), Hum. Genet., 114 (4), pp. 354–365, doi:10.1007/s00439-003-1073-7, PMID 14740294, archived from the original (PDF) on 2011-11-10, retrieved 2012-01-14
- Beleza, Sandra; Gusmao, Leonor; Lopes, Alexandra; Alves, Cintia; Gomes, Iva; Giouzeli, Maria; Calafell, Francesc; Carracedo, Angel; Amorim, Antonio (2006), "Micro-Phylogeographic and Demographic History of Portuguese Male Lineages", Annals of Human Genetics, 70 (2): 181–194, doi:10.1111/j.1529-8817.2005.00221.x, PMID 16626329
- Bird, Steven (2007), "Haplogroup E3b1a2 as a Possible Indicator of Settlement in Roman Britain by Soldiers of Balkan Origin", Journal of Genetic Genealogy, 3 (2)
- Bortolini; Thomas, Mark G.; Chikhi, Lourdes; Aguilar, Juan A.; Castro-De-Guerra, Dinorah; Salzano, Francisco M.; Ruiz-Linares, Andres (2004), "Ribeiro's typology, genomes, and Spanish colonialism, as viewed from Gran Canaria and Colombia" (PDF), Genetics and Molecular Biology, 27 (1): 1–8, doi:10.1590/S1415-47572004000100001
- Bosch, Elena; Calafell, Francesc; Comas, David; Oefner, Peter J.; Underhill, Peter A.; Bertranpetit, Jaume (2001), "High-resolution analysis of human Y-chromosome variation shows a sharp discontinuity and limited gene flow between north-western Africa and the Iberian Peninsula", Am J Hum Genet, 68 (4): 1019–1029, doi:10.1086/319521, PMC 1275654, PMID 11254456
- Bosch, E.; Calafell, F.; Gonzalez-Neira, A.; Flaiz, C.; Mateu, E.; Scheil, H.-G.; Huckenbeck, W.; Efremovska, L.; et al. (2006), "Paternal and maternal lineages in the Balkans show a homogeneous landscape over linguistic barriers, except for the isolated Aromuns", Annals of Human Genetics, 70 (4): 459–487, doi:10.1111/j.1469-1809.2005.00251.x, PMID 16759179, archived from the original on 2012-12-10
- Cadenas; Zhivotovsky, Lev A; Cavalli-Sforza, Luca L; Underhill, Peter A; Herrera, Rene J (2007), "Y-chromosome diversity characterizes the Gulf of Oman", European Journal of Human Genetics, 16 (3): 1–13, doi:10.1038/sj.ejhg.5201934, PMID 17928816
- Capelli, Cristian; Redhead, Nicola; Abernethy, Julia K.; Gratrix, Fiona; Wilson, James F.; Moen, Torolf; Hervig, Tor; Richards, Martin; et al. (2003), "A Y Chromosome Census of the British Isles", Current Biology, 13 (11): 979–84, doi:10.1016/S0960-9822(03)00373-7, PMID 12781138 also at
- Caratti; Gino, S.; Torre, C.; Robino, C. (2009), "Subtyping of Y-chromosomal haplogroup E-M78 (E1b1b1a) by SNP assay and its forensic application", International Journal of Legal Medicine, 123 (4): 357–360, doi:10.1007/s00414-009-0350-y, PMID 19430804
- Capelli, Cristian; Onofri, Valerio; Brisighelli, Francesca; Boschi, Ilaria; Scarnicci, Francesca; Masullo, Mara; Ferri, Gianmarco; Tofanelli, Sergio; et al. (2009), "Moors and Saracens in Europe: estimating the medieval North African male legacy in southern Europe", European Journal of Human Genetics, 17 (6): 848–852, doi:10.1038/ejhg.2008.258, PMC 2947089, PMID 19156170
- Cinnioğlu, Cengiz; King, Roy; Kivisild, Toomas; Kalfoglu, Ersi; Atasoy, Sevil; Cavalleri, Gianpiero L.; Lillie, Anita S.; Roseman, Charles C.; et al. (2004), "Excavating Y-chromosome haplotype strata in Anatolia", Hum Genet, 114 (2): 127–48, doi:10.1007/s00439-003-1031-4, PMID 14586639
- Contu, Daniela; Morelli, Daniela; Santoni, Federico; Foster, Jamie W.; Francalacci, Paolo; Cucca, Francesco (2008), "Y-Chromosome Based Evidence for Pre-Neolithic Origin of the Genetically Homogeneous but Diverse Sardinian Population: Inference for Association Scans" (PDF), PLOS ONE, 3 (1): e1430, doi:10.1371/journal.pone.0001430, PMC 2174525, PMID 18183308, archived from the original (PDF) on 2016-03-04, retrieved 2012-01-14
- Cruciani, Fulvio; Santolamazza, Piero; Shen, Peidong; MacAulay, Vincent; Moral, Pedro; Olckers, Antonel; Modiano, David; Holmes, Susan (2002), "A Back Migration from Asia to Sub-Saharan Africa Is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes", American Journal of Human Genetics, 70 (5): 1197–1214, doi:10.1086/340257, PMC 447595, PMID 11910562
- Cruciani; La Fratta; Santolamazza; Sellitto (May 2004), "Phylogeographic Analysis of Haplogroup E3b (E-M215) Y Chromosomes Reveals Multiple Migratory Events Within and Out Of Africa" (PDF), American Journal of Human Genetics, 74 (5): 1014–1022, doi:10.1086/386294, PMC 1181964, PMID 15042509, archived from the original (PDF) on 2008-06-26, retrieved 2009-07-15
- Cruciani; La Fratta; Torroni; Underhill; Scozzari (2006), "Molecular Dissection of the Y Chromosome Haplogroup E-M78 (E3b1a): A Posteriori Evaluation of a Microsatellite-Network-Based Approach Through Six New Biallelic Markers", Human Mutation, 27 (8): 831–2, doi:10.1002/humu.9445, PMID 16835895
- Cruciani, F.; La Fratta, R.; Trombetta, B.; Santolamazza, P.; Sellitto, D.; Colomb, E. B.; Dugoujon, J.-M.; Crivellaro, F.; et al. (2007), "Tracing Past Human Male Movements in Northern/Eastern Africa and Western Eurasia: New Clues from Y-Chromosomal Haplogroups E-M78 and J-M12", Molecular Biology and Evolution, 24 (6): 1300–1311, doi:10.1093/molbev/msm049, PMID 17351267 Also see Supplementary Data.
- Di Gaetano; Cerutti, Francesca; Crobu, Carlo; Robino (2009), "Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome", European Journal of Human Genetics, 17 (1): 91–99, doi:10.1038/ejhg.2008.120, PMC 2985948, PMID 18685561
- Ehret, C.; Keita, SO; Newman, P (2004), "The Origins of Afroasiatic", Science, 306 (5702): 1680, doi:10.1126/science.306.5702.1680c, PMID 15576591
- El-Sibai, Mirvat; Platt, Daniel E.; Haber, Marc; Xue, Yali; Youhanna, Sonia C.; Wells, R. Spencer; Izaabel, Hassan; Sanyoura, May F.; et al. (2009), "Geographical Structure of the Y-chromosomal Genetic Landscape of the Levant: A coastal-inland contrast", Annals of Human Genetics, 73 (6): 568–581, doi:10.1111/j.1469-1809.2009.00538.x, PMC 3312577, PMID 19686289, archived from the original on 2013-01-05
- Firasat; Khaliq, Shagufta; Mohyuddin, Aisha; Papaioannou, Myrto; Tyler-Smith, Chris; Underhill, Peter A; Ayub, Qasim (2006), "Y-chromosomal evidence for a limited Greek contribution to the Pathan population of Pakistan", European Journal of Human Genetics, 15 (1): 121–126, doi:10.1038/sj.ejhg.5201726, PMC 2588664, PMID 17047675
- Flores, Carlos; Maca-Meyer, Nicole; González, Ana M; Oefner, Peter J; Shen, Peidong; Pérez, Jose A; Rojas, Antonio; Larruga, Jose M; Underhill, Peter A (2004), "Reduced genetic structure of the Iberian peninsula revealed by Y-chromosome analysis: implications for population demography" (PDF), European Journal of Human Genetics, 12 (10): 855–863, doi:10.1038/sj.ejhg.5201225, PMID 15280900, archived from the original (PDF) on 2008-04-06
- Flores; Maca-Meyer, Nicole; Larruga, Jose M.; Cabrera, Vicente M.; Karadsheh, Naif; Gonzalez, Ana M. (2005), "Isolates in a corridor of migrations: a high-resolution analysis of Y-chromosome variation in Jordan", J Hum Genet, 50 (9): 435–441, doi:10.1007/s10038-005-0274-4, PMID 16142507
- Francalacci, P.; Morelli, L.; Underhill, P.A.; Lillie, A.S.; Passarino, G.; Useli, A.; Madeddu, R.; Paoli, G.; et al. (2003), "Peopling of Three Mediterranean Islands (Corsica, Sardinia, and Sicily) Inferred by Y-Chromosome Biallelic Variability", American Journal of Physical Anthropology, 121 (3): 270–279, doi:10.1002/ajpa.10265, PMID 12772214
- Fregel, Rosa; Gomes, Verónica; Gusmão, Leonor; González, Ana M; Cabrera, Vicente M; Amorim, António; Larruga, Jose M (2009), "Demographic history of Canary Islands male gene-pool: replacement of native lineages by European", BMC Evolutionary Biology, 9: 181, doi:10.1186/1471-2148-9-181, PMC 2728732, PMID 19650893
- Gérard; Berriche, S; Aouizérate, A; Diéterlen, F; Lucotte, G (2006), "North African Berber and Arab influences in the western Mediterranean revealed by Y-chromosome DNA haplotypes", Human Biology, 78 (3): 307–316, doi:10.1353/hub.2006.0045, PMID 17216803
- Gonçalves, R; Freitas, A; Branco, M; Rosa, A; Fernandes, AT; Zhivotovsky, LA; Underhill, PA; Kivisild, T; Brehm, A (2005), "Y-chromosome Lineages from Portugal, Madeira and Açores Record Elements of Sephardim and Berber Ancestry", Annals of Human Genetics, 69 (Pt 4): 443–454, doi:10.1111/j.1529-8817.2005.00161.x, PMID 15996172
- Haber, Marc; Platt, Daniel E.; Ashrafian Bonab, Maziar; Youhanna, Sonia C.; Soria-Hernanz, David F.; Martínez-Cruz, Begoña; Douaihy, Bouchra; Ghassibe-Sabbagh, Michella; Rafatpanah, Hoshang; Ghanbari, Mohsen; Whale, John; Balanovsky, Oleg; Wells, R. Spencer; Comas, David; Tyler-Smith, Chris; Zalloua, Pierre A. (2012), "Afghanistan's Ethnic Groups Share a Y-Chromosomal Heritage Structured by Historical Events", PLOS ONE, 7 (3): e34288, doi:10.1371/journal.pone.0034288, PMC 3314501, PMID 22470552
- Hammer (2003), "Human population structure and its effects on sampling Y chromosome sequence variation", Genetics, 164 (4): 1495–1509, PMC 1462677, PMID 12930755
- Hassan, Hisham Y.; Underhill, Peter A.; Cavalli-Sforza, Luca L.; Ibrahim, Muntaser E. (2008), "Y-Chromosome Variation Among Sudanese: Restricted Gene Flow, Concordance With Language, Geography, and History" (PDF), American Journal of Physical Anthropology, 137 (3): 316–23, doi:10.1002/ajpa.20876, PMID 18618658, archived from the original (PDF) on 2009-03-04
- Henn, B. M.; Gignoux, C.; Lin, Alice A; Oefner, Peter J.; Shen, P.; Scozzari, R.; Cruciani, F.; Tishkoff, S. A.; Mountain, J. L.; Underhill, P. A. (2008), "Y-chromosomal evidence of a pastoralist migration through Tanzania to southern Africa", PNAS, 105 (31): 10693–8, doi:10.1073/pnas.0801184105, PMC 2504844, PMID 18678889. See comment on Dienekes blog, comment on the Spitoon blog and public release.
- ISOGG (2013), Y-DNA Haplogroup E and its Subclades - 2013, International Society of Genetic Genealogists "ISOGG"
- Jobling, M.A.; Tyler-Smith, C. (2000), "New uses for new haplotypes the human Y chromosome, disease and selection", Trends Genet., 16 (8): 356–362, doi:10.1016/S0168-9525(00)02057-6, PMID 10904265
- Karafet, T. M.; Mendez, F. L.; Meilerman, M. B.; Underhill, P. A.; Zegura, S. L.; Hammer, M. F. (May 2008), "New Binary Polymorphisms Reshape and Increase Resolution of the Human Y-Chromosomal Haplogroup Tree", Genome Research, 18 (5): 830–8, doi:10.1101/gr.7172008, PMC 2336805, PMID 18385274. Published online April 2, 2008. See also Supplementary Material.
- Keita, Shomarka (2008), "Geography, selected Afro-Asiatic families, and Y chromosome lineage variation", In Hot Pursuit of Language in Prehistory: Essays in the Four Fields of Anthropology : In Honor of Harold Crane Fleming, ISBN 978-90-272-3252-6
- Keita, S. O. Y.; Boyce, A. J. (Anthony J.) (2005), "Genetics, Egypt, and History: Interpreting Geographical Patterns of Y Chromosome Variation", History in Africa, 32: 221–246, doi:10.1353/hia.2005.0013
- King, R. J.; Özcan, S. S.; Carter, T.; Kalfoğlu, E.; Atasoy, S.; Triantaphyllidis, C.; Kouvatsi, A.; Lin, A. A.; et al. (2008), "Differential Y-chromosome Anatolian Influences on the Greek and Cretan Neolithic" (PDF), Annals of Human Genetics, 72 (2): 205–214, doi:10.1111/j.1469-1809.2007.00414.x, PMID 18269686, archived from the original (PDF) on 2009-03-05
- King; Underhill (2002), "Congruent distribution of Neolithic painted pottery and ceramic figurines with Y-chromosome lineages", Antiquity, 76 (293): 707–14, doi:10.1017/S0003598X00091158
- Kujanova; Pereira; Fernandes; Pereira; Cerný (2009), "Near Eastern Neolithic Genetic Input in a Small Oasis of the Egyptian Western Desert", American Journal of Physical Anthropology, 140 (2): 336–346, doi:10.1002/ajpa.21078, PMID 19425100
- Lacan, Marie; Keyser, Christine; Ricaut, François-Xavier; Brucato, Nicolas; Tarrús, Josep; Bosch, Angel; Guilaine, Jean; Crubézy, Eric; Ludes, Bertrand (2011), "Ancient DNA suggests the leading role played by men in the Neolithic dissemination", PNAS, 108 (45): 18255–9, doi:10.1073/pnas.1113061108, PMC 3215063, PMID 22042855
- Lacau, Harlette; Gayden, Tenzin; Regueiro, Maria; Chennakrishnaiah, Shilpa; Bukhari, Areej; Underhill, Peter; Garcia-Bertrand, Ralph; Herrera, Rene (2012), "Afghanistan from a Y-chromosome perspective", European Journal of Human Genetics, 20 (10): 1063–70, doi:10.1038/ejhg.2012.59, PMC 3449065, PMID 22510847
- Lancaster, Andrew (2009), "Y Haplogroups, Archaeological Cultures and Language Families: a Review of the Multidisciplinary Comparisons using the case of E-M243" (PDF), Journal of Genetic Genealogy, 5 (1)
- Luis, J; Rowold, D; Regueiro, M; Caeiro, B; Cinnioglu, C; Roseman, C; Underhill, P; Cavallisforza, L; Herrera, R (2004), "The Levant versus the Horn of Africa: Evidence for Bidirectional Corridors of Human Migrations" (PDF), American Journal of Human Genetics, 74 (3): 532–544, doi:10.1086/382286, PMC 1182266, PMID 14973781, archived from the original (PDF) on 2012-02-16. (Also see Errata)
- Maca-Meyer, N.; Sánchez-Velasco, P.; Flores, C.; Larruga, JM; González, AM; Oterino, A; Leyva-Cobián, F; et al. (2003), "Y Chromosome and Mitochondrial DNA Characterization of Pasiegos, a Human Isolate from Cantabria (Spain)", Annals of Human Genetics, 67 (Pt 4): 329–339, CiteSeerX 10.1.1.584.4253, doi:10.1046/j.1469-1809.2003.00045.x, PMID 12914567.
- Martinez, Laisel; Underhill, Peter A; Zhivotovsky, Lev A; Gayden, Tenzin; Moschonas, Nicholas K; Chow, Cheryl-Emiliane T; Conti, Simon; Mamolini, Elisabetta; Cavalli-Sforza, L Luca; Herrera, Rene (April 1, 2007), "Paleolithic Y-haplogroup heritage predominates in a Cretan highland plateau", European Journal of Human Genetics, 15 (4): 485–493, doi:10.1038/sj.ejhg.5201769, ISSN 1018-4813, PMID 17264870
- Mendizabal, Isabel; Sandoval, Karla; Berniell-Lee, Gemma; Calafell, Francesc; Salas, Antonio; Martinez-Fuentes, Antonio; Comas, David (2008), "Genetic origin, admixture, and asymmetry in maternal and paternal human lineages in Cuba", BMC Evol. Biol., 8: 213, doi:10.1186/1471-2148-8-213, PMC 2492877, PMID 18644108
- Nebel; Filon, D; Brinkmann, B; Majumder, P; Faerman, M; Oppenheim, A (2001), "The Y Chromosome Pool of Jews as Part of the Genetic Landscape of the Middle East", American Journal of Human Genetics, 69 (5): 1095–1112, doi:10.1086/324070, PMC 1274378, PMID 11573163
- Onofri, Valerio; Alessandrini, Federica; Turchi, Chiara; Pesaresi, Mauro; Buscemi, Loredana; Tagliabracci, Adriano (2006), "Development of multiplex PCRs for evolutionary and forensic applications of 37 human Y chromosome SNPs" (PDF), Forensic Science International, 157 (1): 23–35, doi:10.1016/j.forsciint.2005.03.014, PMID 15896936
- Paracchini; Pearce, CL; Kolonel, LN; Altshuler, D; Henderson, BE; Tyler-Smith, C (2003), "A Y chromosomal influence on prostate cancer risk: the multi-ethnic cohort study", J Med Genet, 40 (11): 815–819, doi:10.1136/jmg.40.11.815, PMC 1735314, PMID 14627670
- Pelotti; Ceccardi, S; Lugaresi, F; Trane, R; Falconi, M; Bini, C; Willuweit, S; Roewer, L (2007), "Microgeographic genetic variation of Y chromosome in a population sample of Ravenna's area in the Emilia-Romagna region (North of Italy)", Forensic Science International: Genetics Supplement Series, 1 (1): 242–243, doi:10.1016/j.fsigss.2007.10.025
- Pereira, Luísa; Černý, Viktor; Cerezo, María; Silva, Nuno M; Hájek, Martin; Vašíková, Alžběta; Kujanová, Martina; Brdička, Radim; Salas, Antonio (2010), "Linking the sub-Saharan and West Eurasian gene pools: maternal and paternal heritage of the Tuareg nomads from the African Sahel" (PDF), European Journal of Human Genetics, 18 (8): 915–923, doi:10.1038/ejhg.2010.21, PMC 2987384, PMID 20234393, archived from the original (PDF) on 2013-05-28
- Peričic, M.; Lauc, LB; Klarić, IM; Rootsi, S; Janićijevic, B; Rudan, I; Terzić, R; Colak, I; et al. (2005), "High-resolution phylogenetic analysis of southeastern Europe traces major episodes of paternal gene flow among Slavic populations", Mol. Biol. Evol., 22 (10), pp. 1964–75, doi:10.1093/molbev/msi185, PMID 15944443.
- Ramos-Luisa, E.; Blanco-Verea, A.; Brión, M.; Van Huffel, V.; Carracedo, A.; Sánchez-Diz, P. (2009), "Phylogeography of French male lineages (unpublished data 23rd International ISFG Congress)", Forensic Science International, 2: 439–441, doi:10.1016/j.fsigss.2009.09.026
- Regueiro, M.; Cadenas, A.M.; Gayden, T.; Underhill, P.A.; Herrera, R.J. (2006), "Iran: Tricontinental Nexus for Y-Chromosome Driven Migration", Hum Hered, 61 (3): 132–143, doi:10.1159/000093774, PMID 16770078
- Robino, C.; Crobu, F.; Gaetano, C.; Bekada, A.; Benhamamouch, S.; Cerutti, N.; Piazza, A.; Inturri, S.; Torre, C. (2008), "Analysis of Y-chromosomal SNP haplogroups and STR haplotypes in an Algerian population sample", Journal International Journal of Legal Medicine, 122 (3): 251–5, doi:10.1007/s00414-007-0203-5, PMID 17909833
- Rosa, Alexandra; Ornelas, Carolina; Jobling, Mark A; Brehm, António; Villems, Richard (2007), "Y-chromosomal diversity in the population of Guinea-Bissau: a multiethnic perspective" (PDF), BMC Evolutionary Biology, 7: 124, doi:10.1186/1471-2148-7-124, PMC 1976131, PMID 17662131
- Rosser, Z; Zerjal, T; Hurles, M; Adojaan, M; Alavantic, D; Amorim, A; Amos, W; Armenteros, M; et al. (2000), "Y-Chromosomal Diversity in Europe Is Clinal and Influenced Primarily by Geography, Rather than by Language", American Journal of Human Genetics, 67 (6): 1526–1543, doi:10.1086/316890, PMC 1287948, PMID 11078479, archived from the original on 2008-05-06
- Sanchez, Juan J; Hallenberg, Charlotte; Børsting, Claus; Hernandez, Alexis; Gorlin, RJ (2005), "High frequencies of Y chromosome lineages characterized by E3b1, DYS19-11, DYS392-12 in Somali males", European Journal of Human Genetics, 13 (7): 856–866, doi:10.1038/sj.ejhg.5201390, PMID 15756297. Published online 9 March 2005
- Scozzari, Rosaria; Cruciani, F; Pangrazio, A; Santolamazza, P; Vona, G; Moral, P; Latini, V; Varesi, L; et al. (2001), "Human Y-Chromosome Variation in the Western Mediterranean Area: Implications for the Peopling of the Region" (PDF), Human Immunology, 62 (9): 871–884, CiteSeerX 10.1.1.408.4857, doi:10.1016/S0198-8859(01)00286-5, PMID 11543889
- Semino, O.; Passarino, G; Oefner, PJ; Lin, AA; Arbuzova, S; Beckman, LE; De Benedictis, G; Francalacci, P; et al. (2000), "The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans: A Y Chromosome Perspective" (PDF), Science, 290 (5494), pp. 1155–59, doi:10.1126/science.290.5494.1155, PMID 11073453, archived from the original (PDF) on 2003-11-25.
- Semino; Santachiara-Benerecetti, A. Silvana; Falaschi, Francesco; Cavalli-Sforza, L. Luca; Underhill, Peter A. (2002), "Ethiopians and Khoisan share the deepest clades of the human Y-chromosome phylogeny" (PDF), Am J Hum Genet, 70 (1), pp. 265–268, doi:10.1086/338306, PMC 384897, PMID 11719903, archived from the original (PDF) on 2006-03-15
- Semino, Ornella; Magri, Chiara; Benuzzi, Giorgia; Lin, Alice A.; Al-Zahery, Nadia; Battaglia, Vincenza; MacCioni, Liliana; Triantaphyllidis, Costas; et al. (2004), "Origin, Diffusion, and Differentiation of Y-Chromosome Haplogroups E and J: Inferences on the Neolithization of Europe and Later Migratory Events in the Mediterranean Area", American Journal of Human Genetics, 74 (5), pp. 1023–1034, doi:10.1086/386295, PMC 1181965, PMID 15069642
- Shen, Peidong; Lavi, Tal; Kivisild, Toomas; Chou, Vivian; Sengun, Deniz; Gefel, Dov; Shpirer, Issac; Woolf, Eilon; et al. (2004), "Reconstruction of Patrilineages and Matrilineages of Samaritans and Other Israeli Populations From Y-Chromosome and Mitochondrial DNA Sequence Variation" (PDF), Human Mutation, 24 (3), pp. 248–60, doi:10.1002/humu.20077, PMID 15300852, archived from the original (PDF) on 2009-03-05, retrieved 2009-07-15
- Silva; Carvalho, Elizeu; Costa, Guilherme; Tavares, Lígia; Amorim, António; Gusmão, Leonor (2006), "Y-chromosome genetic variation in Rio De Janeiro population", American Journal of Human Biology, 18 (6): 829–837, doi:10.1002/ajhb.20567, PMID 17039481, archived from the original on 2012-10-18
- Shlush; Behar, Doron M.; Yudkovsky, Guennady; Templeton, Alan; Hadid, Yarin; Basis, Fuad; Hammer, Michael; Itzkovitz, Shalev; Skorecki, Karl (2008), Gemmell, Neil John (ed.), "The Druze: A Population Genetic Refugium of the Near East", PLOS ONE, 3 (5): e2105, doi:10.1371/journal.pone.0002105, PMC 2324201, PMID 18461126
- Sykes, Bryan (2006), Blood of the Isles: Exploring the Genetic Roots of Our Tribal History, Bantam, ISBN 978-0-593-05652-3
- Thomas; Stumpf, M. P.H; Harke, H. (2006), "Evidence for an apartheid-like social structure in early Anglo-Saxon England" (PDF), Proceedings of the Royal Society B, 273 (1601): 2651–2657, doi:10.1098/rspb.2006.3627, PMC 1635457, PMID 17002951, archived from the original (PDF) on 2009-03-05
- Trombetta, Beniamino; Cruciani, Fulvio; Sellitto, Daniele; Scozzari, Rosaria (2011), MacAulay, Vincent (ed.), "A New Topology of the Human Y Chromosome Haplogroup E1b1 (E-P2) Revealed through the Use of Newly Characterized Binary Polymorphisms", PLOS ONE, 6 (1): e16073, doi:10.1371/journal.pone.0016073, PMC 3017091, PMID 21253605
- Trombetta (2015), "Phylogeographic refinement and large scale genotyping of human Y chromosome haplogroup E provide new insights into the dispersal of early pastoralists in the African continent", Genome Biology and Evolution, 7 (7): 1940–1950, doi:10.1093/gbe/evv118, PMC 4524485, PMID 26108492
- Underhill, Peter A.; Shen, Peidong; Lin, Alice A.; Jin, Li; Passarino, Giuseppe; Yang, Wei H.; Kauffman, Erin; Bonné-Tamir, Batsheva; et al. (2000), "Y chromosome sequence variation and the history of human populations", Nat Genet, 26 (3), pp. 358–361, doi:10.1038/81685, PMID 11062480
- Underhill; Passarino, G.; Lin, A. A.; Shen, P.; Mirazon Lahr, M.; Foley, R. A.; Oefner, P. J.; Cavalli-Sforza, L. L. (2001), "The phylogeography of Y chromosome binary haplotypes and the origins of modern human populations" (PDF), Ann Hum Genet, 65 (Pt 1), pp. 43–62, doi:10.1046/j.1469-1809.2001.6510043.x, PMID 11415522, archived from the original (PDF) on 2011-07-21
- Underhill (2002), Bellwood and Renfrew (ed.), Inference of Neolithic Population Histories using Y-chromosome Haplotypes, Cambridge: McDonald Institute for Archaeological Research, ISBN 978-1-902937-20-5
- Underhill, Peter A.; Kivisild, Toomas (2007), "Use of Y Chromosome and Mitochondrial DNA Population Structure in Tracing Human Migrations", Annu. Rev. Genet., 41: 539–64, doi:10.1146/annurev.genet.41.110306.130407, PMID 18076332
- Weale, M. E.; Weiss, D. A.; Jager, R. F.; Bradman, N.; Thomas, M. G. (2002), "Y Chromosome Evidence for Anglo-Saxon Mass Migration" (PDF), Mol. Biol. Evol., 19 (7), pp. 1008–1021, doi:10.1093/oxfordjournals.molbev.a004160, PMID 12082121.
- Weale; Shah, T; Jones, AL; Greenhalgh, J; Wilson, JF; Nymadawa, P; Zeitlin, D; Connell, BA; et al. (September 1, 2003), "Rare Deep-Rooting Y Chromosome Lineages in Humans: Lessons for Phylogeography", Genetics, 165 (1), pp. 229–234, PMC 1462739, PMID 14504230
- Y Chromosome Consortium "YCC" (2002), "A Nomenclature System for the Tree of Human Y-Chromosomal Binary Haplogroups", Genome Research, 12 (2), pp. 339–348, doi:10.1101/gr.217602, PMC 155271, PMID 11827954
- Zalloua, Pierre A.; Xue, Yali; Khalife, Jade; Makhoul, Nadine; Debiane, Labib; Platt, Daniel E.; Royyuru, Ajay K.; Herrera, Rene J.; et al. (2008), "Y-Chromosomal Diversity in Lebanon Is Structured by Recent Historical Events", American Journal of Human Genetics, 82 (4): 873–882, doi:10.1016/j.ajhg.2008.01.020, PMC 2427286, PMID 18374297
- Zalloua, Pierre A.; Platt, Daniel E.; El Sibai, Mirvat; Khalife, Jade; Makhoul, Nadine; Haber, Marc; Xue, Yali; Izaabel, Hassan; et al. (2008), "Identifying Genetic Traces of Historical Expansions: Phoenician Footprints in the Mediterranean", The American Journal of Human Genetics, 83 (5): 633–642, doi:10.1016/j.ajhg.2008.10.012, PMC 2668035, PMID 18976729
- Zerjal (1999), The use of Y-chromosomal DNA variation to investigate population history; in Papiha SS, Deka R, Chakraborty R (eds): Genomic diversity: applications in human population genetics, Kluwer Academic/Plenum Publishers, pp. 91–101
- Zhao; Khan, Faisal; Borkar, Minal; Herrera, Rene; Agrawal, Suraksha (2009), "Presence of three different paternal lineages among North Indians: A study of 560 Y chromosomes", Annals of Human Biology, 36 (1): 1–14, doi:10.1080/03014460802558522, PMC 2755252, PMID 19058044