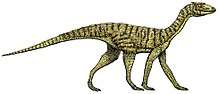
Proterosuchus
Proterosuchus is an extinct genus of archosauriform reptiles that lived during the Early Triassic. It contains three valid species: the type species P. fergusi and the referred species P. alexanderi and P. goweri. All three species lived in what is now South Africa. The genus was named in 1903 by the South African paleontologist Robert Broom. The well-known genus Chasmatosaurus is a junior synonym of Proterosuchus.
| Proterosuchus | |
|---|---|
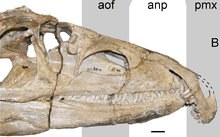 | |
| Skull of P. fergusi | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Clade: | Archosauriformes |
| Family: | †Proterosuchidae |
| Genus: | †Proterosuchus Broom, 1903 |
| Type species | |
| †Proterosuchus fergusi Broom, 1903 | |
| Species | |
| |
| Synonyms | |
| |
Proterosuchus was a mid-sized quadrupedal reptile with a sprawling stance that could reach a length of up to 3.5 meters (11 ft). It had a large head and distinctively hooked snout. It was a predator, which may have hunted prey such as Lystrosaurus. The lifestyle of Proterosuchus remains debated; it may have been terrestrial or it may have been a semiaquatic ambush predator similar to modern crocodiles.
Proterosuchus is one of the earliest members of the clade Archosauriformes, which also includes crocodilians, pterosaurs, and dinosaurs, including birds. It lived in the aftermath of the Permian–Triassic extinction event, the largest known mass extinction in Earth's history.
Description

Proterosuchus was a quadrupedal reptile with a sprawling stance.[1] It could reach a total length of up to 3.5 meters (11 ft).[2] Like most reptiles, Proterosuchus had scaly skin.[3]
Proterosuchus had a proportionally large head and long neck compared to its body.[4][5] The most distinctive characteristic of its head was its strongly hooked snout, formed by a downturned premaxilla.[6] The premaxilla contained up to nine teeth in adults, and the teeth in the snout tip were splayed out to the sides.[6] The jaws of Proterosuchus contained numerous teeth, with up to 9 premaxillary, 31 maxillary, and 28 dentary teeth in each side.[7] The teeth of Proterosuchus were recurved, labiolingually compressed, and serrated, as in most archosauriforms. They were isodont, or all equal in size and shape, in adult individuals, but in juveniles, the teeth were less strongly curved in the back of the jaw.[7]
The skull of Proterosuchus exhibits many features characteristic of its position as a basal archosauriform. It bears a prominent antorbital fenestra, like most archosauriforms. In some specimens, the jugal and quadratojugal contact to complete the ventral margin of the lower temporal fenestra, as in other archosauriforms, but in other specimens, there is a narrow gap between the bones so that the lower temporal bar is incomplete as in non-archosauriform archosauromorphs.[2] The lower jaw bears a small external mandibular fenestra, another characteristic of archosauriforms and their closest relatives.[8]
Paleobiology
The lifestyle of Proterosuchus is debated. It has conventionally been depicted as a semiaquatic ambush predator similar to modern crocodiles. However, it lived in an arid environment and many aspects of its anatomy conflict with a semiaquatic lifestyle.[9] In particular, its limbs are well-ossified, as in terrestrial animals, and the nostrils are laterally-positioned on the snout, not dorsally-positioned. The histology of its bones is reminiscent of terrestrial animals, not semiaquatic ones.[10] However, support for a semiaquatic lifestyle comes from its brain anatomy, which resembles semiaquatic predators such as crocodiles more closely than terrestrial reptiles. The orientation of its ear canals suggests its neutral head posture had the snout angled upward, which would have raised the nostrils high enough for the animal to breathe while largely submerged.[11] Proterosuchus was a predator, but the specifics of its diet are not known. It has been suggested to have eaten fish or the abundant contemporary dicynodont Lystrosaurus.[9]
Snout function
The function of the hooked snout in Proterosuchus is not fully known. The most likely use was in sexual or social signaling, similar to the hooked snout of male salmon. As the snout does not appear to have been sexually dimorphic, it may be an example of mutual sexual selection. The snout may have been used in a specialized method of predation, as it exhibits high resistance to dorsoventral bending.[11] However, what this method may have been is unclear. The premaxillary teeth do not show wear facets and did not occlude with the teeth of the lower jaw, indicating that they were not used in any abrasive activities and could not have been used to grip prey. The snout tip did not have the pressure receptors present in crocodilians and Spinosaurus.[6]
Senses
Proterosuchus had mesopic vision, indicating that it was adapted to see well in both bright and dim light. Mesopic vision is characteristic of cathemeral animals, which are active in both night and day, and crepuscular animals, which are active in twilight. Adaptations to see in dim light may have been ancestral to archosaurs, and Proterosuchus may have been an early example of this trend.[12] However, Proterosuchus lived near the Antarctic Circle, so its mesopic vision may have instead been an adaptation to the highly seasonal day lengths it experienced.[13] The hearing of Proterosuchus was likely adapted for lower frequencies, as in modern crocodiles. Due to its low-sensitivity hearing, Proterosuchus probably did not rely heavily on vocal communication and may have been relatively solitary. Based on the size of its olfactory bulbs, Proterosuchus had a strong sense of smell, similar to that of modern crocodiles. However, its olfactory bulbs were not as large as those of its relative Tasmaniosaurus, suggesting different habits and potentially a more aquatic ecology in Proterosuchus.[11]
Metabolism
The metabolism of Proterosuchus is disputed. Like other crocopod archosauromorphs, Proterosuchus had a higher metabolic rate than extant ectotherms.[14] Furthermore, Proterosuchus possessed fibrolamellar bone, indicative of a high growth rate and corresponding high metabolism.[10][15] However, studies conflict on whether the metabolism of Proterosuchus was within the range of extant endotherms.[15] Its metabolic rate was lower than most other crocopods, except for the ectothermic phytosaurs and crocodilians, which may have been an adaptation to a crocodile-like predatory strategy.[14]
Ontogeny
Proterosuchus grew quickly. It probably reached sexual maturity within a year, at roughly two-thirds of its maximum adult body size.[7] Rapid growth rates were typical of Early Triassic archosauromorphs, and may have been an adaptation to surviving the hostile environment of the Early Triassic.[10] Juvenile Proterosuchus may have hunted different prey from adults.[7]
Paleoecology
Proterosuchus fossils are found in the Lystrosaurus Assemblage Zone of the Beaufort Group in South Africa. Proterosuchus was the first new species to arrive in the Karoo environment after the Permian–Triassic extinction. Proterosuchus and the therocephalian Moschorhinus were the largest carnivores in the ecosystem at the time, and soon after the extinction Moschorhinus declined and went extinct while Proterosuchus thrived.[16] The most common tetrapod in Proterosuchus's environment was the herbivorous dicynodont Lystrosaurus. The environment was hot, semi-arid and experienced droughts.[17]
Classification
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Phylogenetic position of Proterosuchus within Crocopoda[18][19] |
Proterosuchus is an early member of Archosauriformes, which also contains crocodilians, pterosaurs, and dinosaurs, including birds. It is the type genus of Proterosuchidae, which also contains the genus Archosaurus. Proterosuchidae is, by definition, the most basal clade of archosauriforms, as Archosauriformes is defined based on their phylogenetic position.[20] Under pre-cladistic taxonomy, Proterosuchus was classified in the order Thecodontia and suborder Proterosuchia. Both taxa are now recognized as paraphyletic groups of basal archosauriforms.
Species
Valid Species
Proterosuchus currently contains three valid species, all from the Lower Triassic of South Africa.
- Proterosuchus fergusi is the type species of Proterosuchus. It was named in 1903 by Robert Broom based on a specimen from Tarkastad donated by John Fergus, for whom the species was named.[21] It is known from several specimens, and the species Chasmatosaurus vanhoepeni and Elaphrosuchus rubidgei are junior synonyms of it. The holotype is poorly preserved and indeterminate, and a neotype has been suggested. It is distinguished from other species of Proterosuchus by its more strongly curved quadrate.[2]
- Proterosuchus alexanderi was named by A. Hoffmann in 1965 based on a subadult specimen.[22][7] It is currently only known from one specimen.[2] It is distinguished from other species of Proterosuchus by its longer snout.[2]
- Proterosuchus goweri was named by Martín D. Ezcurra and Richard J. Butler in 2015, based on a specimen that had originally been described as a specimen of Chasmatosaurus vanhoepeni.[2] It is currently only known from one specimen.[2] It is distinguished from other species of Proterosuchus by a deep horizontal process of the maxilla, a sinusoidal ventral margin of the maxilla, and a gap in the tooth row between the premaxilla and maxilla.[2]
Other Species

Several species have been assigned to Proterosuchus or its junior synonym Chasmatosaurus in the past that are either no longer valid or no longer assigned to Proterosuchus.
- Ankistrodon indicus was named in 1865 by Thomas Henry Huxley, based on a specimen from the Induan-age Panchet Formation of India.[23][18] Ankistrodon has been regarded as a synonym of Proterosuchus or Chasmatosaurus in the past, but if this synonymy were correct, Ankistrodon would have priority over the other names.[24] It is now considered a nomen dubium.[24][18]
- Chasmatosaurus vanhoepeni is the type species of Chasmatosaurus. It was named in 1924 by Haughton.[25] The species name honors E. C. N. van Hoepen, who collected and prepared the holotype.[25] It is now considered a junior synonym of Proterosuchus fergusi.[2] Like all P. fergusi specimens, it is from the Lystrosaurus Assemblage Zone of the Beaufort Group of South Africa.
- Chasmatosaurus yuani was named by C. C. Young in 1936, based on specimens from the Induan-age Jiucaiyuan Formation of China.[26] It is considered a valid species of proterosuchid, but is not formally assigned to Proterosuchus.[27][18] It is considered to be in need of taxonomic revision.[27] It is more closely related to Proterosuchus goweri than to other species of Proterosuchus.[18]
- Elaphrosaurus rubidgei was named by Robert Broom in 1946.[28] It is now considered a junior synonym of Proterosuchus fergusi, with the holotype being a juvenile specimen thereof.[2] Like all P. fergusi specimens, it is from the Lystrosaurus Assemblage Zone of the Beaufort Group of South Africa.
- Chasmatosaurus ultimus was named by C. C. Young in 1964, based on a specimen from the Anisian-age Ermaying Formation of China.[29][18] It was long believed to be the geologically youngest species of proterosuchid, as it would be the only one from the Middle Triassic.[24][30] However, it is no longer considered to be a proterosuchid and is now considered to be a suchian archosaur.[18][30] It is now considered a nomen dubium.[30]
References
- Sullivan, Corwin (2015). "Evolution of hind limb posture in Triassic archosauriforms". In Dial, Kenneth P.; Shubin, Neil; Brainerd, Elizabeth L. (eds.). Great Transformations in Vertebrate Evolution. University of Chicago Press.
- Ezcurra, Martín D.; Butler, Richard J. (2015). "Taxonomy of the proterosuchid archosauriforms (Diapsida: Archosauromorpha) from the earliest Triassic of South Africa, and implications for the early archosauriform radiation". Palaeontology. 58 (1): 141–170. doi:10.1111/pala.12130.
- Thornley, A. L. (1970). "Epidermal remnants of Proterosuchus vanhoepeni". Palaeontologia Africana. 13: 57–60.
- Butler, Richard J.; Sennikov, Andrey G.; Dunne, Emma M.; Ezcurra, Martin D.; Hedrick, Brandon P.; Maidment, Susannah C. R.; Meade, Luke E.; Raven, Thomas J.; Gower, David J. (2019). "Cranial anatomy and taxonomy of the erythrosuchid archosauriform 'Vjushkovia triplicostata' Huene, 1960, from the Early Triassic of European Russia". Royal Society Open Science. 6 (11): 191289. doi:10.1098/rsos.191289. PMC 6894557. PMID 31827861.
- Charig, Alan J.; Sues, Hans-Dieter; Charig, Alan J.; Krebs, Bernard; Sues, Hans-Dieter; Westphal, Frank (1976). "Suborder Proterosuchia Broom 1906b". In Oskar Kuhn (ed.). Handbuch der Paläoherpetologie: Teil 13: Thecodontia. Handbuch der Paläoherpetologie (1st ed.). Stuttgart: Gustav Fischer Verlag. pp. 11–39. ISBN 3-437-30184-5.
- Ezcurra, Martín D. (2017). "Can social and sexual selection explain the bizarre snout of proterosuchid archosauriforms?". Historical Biology. 29 (3): 348–358. doi:10.1080/08912963.2016.1161033.
- Ezcurra, Martín D.; Butler, Richard J. (2015). "Post-hatchling cranial ontogeny in the Early Triassic diapsid reptile Proterosuchus fergusi". Journal of Anatomy. 226 (5): 387–402. doi:10.1111/joa.12300. PMC 4450940. PMID 25913624.
- Welman, J. (1998). "The taxonomy of the South African proterosuchids (Reptilia, Archosauromorpha)". Journal of Vertebrate Paleontology. 18 (2): 340–347. doi:10.1080/02724634.1998.10011062.
- Ezcurra, Martín D.; Butler, Richard J.; Gower, David J. (2013). "'Proterosuchia': the origin and early history of Archosauriformes". Geological Society, London, Special Publications. 379 (1): 9–33. doi:10.1144/SP379.11.
- Botha-Brink, Jennifer; Smith, Roger M. H. (2011). "Osteohistology of the Triassic archosauromorphs Prolacerta, Proterosuchus, Euparkeria, and Erythrosuchus from the Karoo Basin of South Africa". Journal of Vertebrate Paleontology. 31 (6): 1238–1254. doi:10.1080/02724634.2011.621797. JSTOR 41407682.
- Brown, Emily E.; Butler, Richard J.; Ezcurra, Martín D.; Bhullar, Bhart-Anjan S.; Lautenschlager, Stephan (2019). "Endocranial anatomy and life habits of the Early Triassic archosauriform Proterosuchus fergusi". Palaeontology. 63 (2): 255–282. doi:10.1111/pala.12454.
- Chang, Belinda S. W.; Jönsson, Karolina; Kazmi, Manija A.; Donoghue, Michael J.; Sakmar, Thomas P. (2002). "Recreating a Functional Ancestral Archosaur Visual Pigment". Molecular Biology and Evolution. 19 (9): 1483–1489. doi:10.1093/oxfordjournals.molbev.a004211. PMID 12200476.
- Schmitz, L.; Motani, R. (2011). "Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology". Science. 332 (6030): 705–8. doi:10.1126/science.1200043. PMID 21493820.
- Legendre, Lucas J.; Guénard, Guillaume; Botha-Brink, Jennifer; Cubo, Jorge (2016). "Paleohistological evidence for ancestral high metabolic rate in archosaurs". Systematic Biology. 65 (6): 989–986. doi:10.1093/sysbio/syw033. PMID 27073251.
- Cubo, Jorge; Jalil, Nour-Eddine (2019). "Bone histology of Azendohsaurus laaroussii: Implications for the evolution of thermometabolism in Archosauromorpha". Paleobiology. 45 (2): 317–330. doi:10.1017/pab.2019.13.
- Smith, Roger; Botha, Jennifer (2005). "The recovery of terrestrial vertebrate diversity in the South African Karoo Basin after the end-Permian extinction". Comptes Rendus Palevol. 4 (6–7): 623–636. doi:10.1016/j.crpv.2005.07.005.
- Smith, Roger; Rubidge, Bruce; van der Walt, Merrill (2012). "Therapsid biodiversity patterns and paleoenvironments of the Karoo Basin, South Africa". In Anusuya Chinsamy-Turan (ed.). Forerunners of Mammals: Radiation, Histology, Biology. Bloomington: Indiana University Press. pp. 31–62. ISBN 978-0-253-35697-0.
- Ezcurra, Martín D. (2016). "The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms". PeerJ. 4: e1778. doi:10.7717/peerj.1778. PMC 4860341. PMID 27162705.
- Pinheiro, Felipe L.; de Simão-Oliveira, Daniel; Butler, Richard J. (2019). "Osteology of the archosauromorph Teyujagua paradoxa and the early evolution of the archosauriform skull". Zoological Journal of the Linnean Society. 189: 378–417. doi:10.1093/zoolinnean/zlz093.
- Nesbitt, Sterling J. (2011). "The early evolution of archosaurs: relationships and the origin of major clades". Bulletin of the American Museum of Natural History. 352: 1–292. doi:10.1206/352.1.
- Broom, R. (1903). "On a new reptile (Proterosuchus fergusi) from the Karroo beds of Tarkastad, South Africa". Annals of the South African Museum. 4: 159–164.
- Hoffmann, A.C. (1965). "On the discovery of a new thecodont from the Middle Beaufort Beds". Researches of the National Museum, Bloemfontein. 2: 33–40.
- Huxley, Thomas H. (1865). "On a collection of vertebrate fossils from the Panchet Rocks, Ranigunj, Bengal". Memoirs of the Geological Survey of India.
- Charig, A. J.; Reig, O. A. (1970). "The classification of the Proterosuchia". Biological Journal of the Linnean Society. 2 (2): 125–171. doi:10.1111/j.1095-8312.1970.tb01708.x.
- Haughton, S.H. (1924). "On a new type of thecodont from the Middle Beaufort Beds". Annals of the Transvaal Museum. 11: 93–97.
- Young, C. C. (1936). "On a new Chasmatosaurus from Sinkiang". Bulletin of the Geological Society of China. 15 (3): 291–311. doi:10.1111/j.1755-6724.1936.mp15003003.x.
- Nesbitt, Sterling J.; Flynn, John J.; Pritchard, Adam C.; Parrish, J. Michael; Ranivoharimanana, Lovasoa; Wyss, André R. (2015). "Postcranial osteology of Azendohsaurus madagaskarensis (?Middle to Upper Triassic, Isalo Group, Madagascar) and its systematic position among stem archosaur reptiles". Bulletin of the American Museum of Natural History. 398: 1–126. doi:10.1206/amnb-899-00-1-126.1.
- Broom, R. (1946). "A new primitive proterosuchid reptile". Annals of the Transvaal Museum. 20: 343–346.
- Young, C.-C. (1964). "The pseudosuchians in China". Palaeontologia Sincia, New Series C. 151 (19): 1–205.
- Liu, Jun; Butler, Richard; Sullivan, Corwin; Ezcurra, Martin (2015). "'Chasmatosaurus ultimus,' a putative proterosuchid archosauriform from the Middle Triassic, is an indeterminate crown archosaur". Journal of Vertebrate Paleontology. 35 (5): –965779. doi:10.1080/02724634.2015.965779.