Chaetognatha
Chaetognatha (meaning bristle-jaws) is a phylum of predatory marine worms that are a major component of plankton worldwide. Commonly known as arrow worms, about 20% of the known Chaetognatha species are benthic, and can attach to algae and rocks. They are found in all marine waters, from surface tropical waters and shallow tide pools to the deep sea and polar regions. Most chaetognaths are transparent and are torpedo shaped, but some deep-sea species are orange. They range in size from 2 to 120 millimetres (0.1 to 4.7 in).
| Arrow worms | |
|---|---|
 | |
| Spadella cephaloptera | |
| Scientific classification | |
| Kingdom: | Animalia |
| Subkingdom: | Eumetazoa |
| Clade: | ParaHoxozoa |
| Clade: | Bilateria |
| Clade: | Nephrozoa |
| (unranked): | Protostomia |
| (unranked): | Spiralia |
| Clade: | Gnathifera |
| Phylum: | Chaetognatha Leuckart, 1854 |
| Class: | Sagittoidea Claus & Grobben, 1905 [2] |
| Orders | |
| |
There are more than 120 modern species assigned to over 20 genera.[3] Despite the limited diversity of species, the number of individuals is large.[4]
Arrow worms are usually considered a type of protostome that do not belong to either Ecdysozoa or Lophotrochozoa.
Anatomy
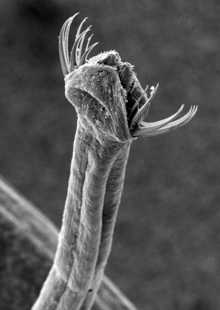
Chaetognaths are transparent or translucent dart-shaped animals covered by a cuticle. The body is divided into a distinct head, trunk, and tail. There are between four and fourteen hooked, grasping spines on each side of their head, flanking a hollow vestibule containing the mouth. The spines are used in hunting, and covered with a flexible hood arising from the neck region when the animal is swimming. All chaetognaths are carnivorous, preying on other planktonic animals.[5] Some species are also reported to be omnivores, feeding on algae and detritus[6].
The trunk bears one or two pairs of lateral fins incorporating structures superficially similar to the fin rays of fish, with which they are not homologous, however: unlike those of vertebrates, these are composed of a thickened basement membrane extending from the epidermis. An additional caudal fin covers the post-anal tail.[5] Two chaetognath species, Caecosagitta macrocephala and Eukrohnia fowleri, have bioluminescent organs on their fins.[7][8]
Chaetognaths swim in short bursts using a dorso-ventral undulating body motion, where their tail fin assists with propulsion and the body fins with stabilization and steering.[9] Some species are known to use the neurotoxin tetrodotoxin to subdue prey.[10]
The body cavity is lined by peritoneum, and therefore represents a true coelom, and is divided into one compartment on each side of the trunk, and additional compartments inside the head and tail, all separated completely by septa. Although they have a mouth with one or two rows of tiny teeth, compound eyes, and a nervous system, they have no respiratory or circulatory systems.
The mouth opens into a muscular pharynx, which contains glands to lubricate the passage of food. From here, a straight intestine runs the length of the trunk to an anus just forward of the tail. The intestine is the primary site of digestion and includes a pair of diverticula near the anterior end.[5] Materials are moved about the body cavity by cilia. Waste materials are simply excreted through the skin and anus. Eukrohniid species possess an oil vacuole closely associated with the gut. This organ contains wax esters which may assist reproduction and growth outside of the production season for Eukrohnia hamata in Arctic seas[11]. Owing to the position of the oil vacuole in the center of the tractus, the organ may also have implications for buoyancy, trim and locomotion[12].
The nervous system is reasonably simple, consisting of a ganglionated nerve ring surrounding the pharynx. The dorsal ganglion is the largest, but nerves extend from all the ganglia along the length of the body. Chaetognaths have two compound eyes, each consisting of a number of pigment-cup ocelli fused together. In addition, there are a number of sensory bristles arranged in rows along the side of the body, where they probably perform a function similar to that of the lateral line in fish. An additional, curved, band of sensory bristles lies over the head and neck.[5]
The arrow worm rhabdomeres are derived from microtubules 20 nm long and 50 nm wide, which in turn form conical bodies that contain granules and thread structures. The cone body is derived from a cilium.[13]
Reproduction
All species are hermaphroditic, carrying both eggs and sperm.[4] Each animal possesses a pair of testes within the tail, and a pair of ovaries in the posterior region of the main body cavity. Immature sperm are released from the testes to mature inside the cavity of the tail, and then swim through a short duct to a seminal vesicle where they are packaged into a spermatophore.[5](Sexually and Asexually)
During mating, each individual places a spermatophore onto the neck of its partner after rupture of the seminal vesicle. The sperm rapidly escape from the spermatophore and swim along the midline of the animal until they reach a pair of small pores just in front of the tail. These pores connect to the oviducts, into which the developed eggs have already passed from the ovaries, and it is here that fertilisation takes place.[5]
The eggs are planktonic, develop in marsupial sacs or attached to algae, and hatch into miniature versions of the adult, without a well-defined larval stage.[5][14]
Classification
Chaetognaths are traditionally classed as deuterostomes by embryologists. Lynn Margulis and K. V. Schwartz place chaetognaths in the deuterostomes in their Five Kingdom classification.[15] Molecular phylogenists, however, consider them to be protostomes. Thomas Cavalier-Smith places them in the protostomes in his Six Kingdom classification.[16] The similarities between chaetognaths and nematodes mentioned above may support the protostome thesis—in fact, chaetognaths are sometimes regarded as a basal ecdysozoan or lophotrochozoan.[17] Chaetognatha appears close to the base of the protostome tree in most studies of their molecular phylogeny.[18] This may explain their deuterostome embryonic characters. If chaetognaths branched off from the protostomes before they evolved their distinctive protostome embryonic characters, they might have retained deuterostome characters inherited from early bilaterian ancestors. Thus chaetognaths may be a useful model for the ancestral bilaterian.[19] Studies of arrow worms' nervous systems suggests they should be placed within the protostomes.[20][21] According to 2017 and 2019 papers, chaetognaths appear related to gnathiferans.[22][23]
Taxonomy
Class Sagittoidea Claus & Grobben 1905[24]
- Subclass Eukrohniones Kassatkina 2011
- Order Biphragmoeukrohniformes Kassatkina 2012
- Family Protoeukrohniidae Kassatkina 2012
- Genus Protoeukrohnia Kassatkina 2003
- Family Heterokrohniidae Сasanova 1986
- Genus Archeterokrohnia Casanova 1986
- Genus Heterokrohnia Ritter-Záhony 1911
- Genus Xenokrohnia Casanova 1993
- Family Tokiokaispadellidae Salvini-Plaven 1986
- Genus Aberrospadella Kassatkina 1971
- Genus Entokrohnia Kassatkina 2012
- Genus Tokiokaispadella Kassatkina 2008
- Genus Protoheterokrohnia Kassatkina 2016
- Family Protoeukrohniidae Kassatkina 2012
- Order Monophragmoeukrohniformes Kassatkina 2012
- Family Eukrohniidae Tokioka 1965
- Genus Eukrohnia Ritter-Záhony 1909
- Genus Paraeukrohnia Kasatkina & Stolyarova 2006
- Genus Praeukrohnia Kasatkina & Stolyarova 2006
- Genus Pseudoeukrohnia asatkina 1998
- Family Eukrohniidae Tokioka 1965
- Order Aphragmoeukrohniformes Kassatkina 2012
- Family Pterosagittidae Tokioka 1965
- Genus Pterosagitta Costa 1869
- Family Krohnittidae Tokioka 1965
- Genus Krohnitta Ritter-Záhony 1910
- Family Krohnittellidae Bieri 1989
- Genus Prakrohnitella Kassatkina 2017
- Genus Protokrohnitella Kasatkina 2003
- Genus Krohnittella Germain & Joubin 1912
- Family Pterosagittidae Tokioka 1965
- Order Biphragmoeukrohniformes Kassatkina 2012
- Subclass Spadelliones Kassatkina 2011
- Family Spadellidae Tokioka 1965
- Genus Bathyspadella Tokioka 1939
- Genus Calispadella Casanova & Moreau 2005
- Genus Hemispadella Casanova 1996
- Genus Paraspadella Salvini-Plawen 1986
- Genus Spadella Langerhans 1880
- Family Spadellidae Tokioka 1965
- Subclass Sagittiones Kassatkina 2011
- Family ?Pterokrohniidae Bieri 1991
- Genus Pterokrohnia Srinivasan 1988
- Family ?Bathybelidae Bieri 1989
- Genus Bathybelos Owre 1974
- Order Biphragmosagittiformes Kassatkina 2012
- Family Biphragmosagittidae Kassatkina 2012
- Genus Biphragmosagitta Kassatkina 2012
- Genus Biphragmofastigata Kassatkina 2012
- Family Biphragmosagittidae Kassatkina 2012
- Order Aphragmophora Tokioka 1965
- Family Sagittidae Claus & Grobben 1905
- Subfamily Flaccisagittinae Kassatkina 2007
- Genus Flaccisagitta Tokioka 1965
- Genus Pseudosagitta Germain & Joubin 1912
- Genus Accedosagitta Kassatkina 1971
- Genus Abosagitta Kassatkina 2006
- Subfamily Sagittinae Claus et Groben 1905
- Genus Aidanosagitta Tokioka 1965
- Genus Bathysagitta Kasatkina 2001
- Genus CaecosagittaTokioka, 1965
- Genus Demisagitta Bieri 1991
- Genus Eucaecosagitta Kasatkina 2003
- Genus Ferosagitta Kassatkina 1971
- Genus Leptosagitta Kassatkina 1973
- Genus Mesosagitta Tokioka 1965
- Genus Oligoradiata Kassatkina 1971
- Genus Omittosagitta Kassatkina 1971
- Genus Parasagitta Tokioka 1965
- Genus Pseudomesosagitta Kassatkina & Stolyarova 2010
- Genus Sagitta Quoy & Gaimard 1827
- Genus Serratosagitta Tokioka 1965
- Genus Solidosagitta Tokioka 1965
- Genus Vitreosagitta Kassatkina 2010
- Genus Zonosagitta Tokioka 1965
- Subfamily Flaccisagittinae Kassatkina 2007
- Family Sagittidae Claus & Grobben 1905
- Family ?Pterokrohniidae Bieri 1991
Fossil record
Due to their soft bodies, chaetognaths fossilize poorly. Even so, several fossil chaetognath species have been described.[1] Chaetognaths appear to have originated in the Cambrian Period. Complete body fossils have been formally described from the Lower Cambrian Maotianshan shales of Yunnan, China (Eognathacantha ercainella Chen & Huang[25] and Protosagitta spinosa Hu[26]) and the Middle Cambrian Burgess Shale of British Columbia (Oesia disjuncta Walcott[27]), a view challenged by Conway Morris (2009). A more recent chaetognath, Paucijaculum samamithion Schram, has been described from the Mazon Creek biota from the Pennsylvanian of Illinois. Chaetognaths were thought possibly to be related to some of the animals grouped with the conodonts. The conodonts themselves, however, are thought to be related to the vertebrates. It is now thought that protoconodont elements (e.g., Protohertzina anabarica Missarzhevsky, 1973), are probably grasping spines of chaetognaths rather than teeth of conodonts. Previously chaetognaths in the Early Cambrian were only suspected from these protoconodont elements, but the more recent discoveries of body fossils have confirmed their presence then.[28]
Infection by giant viruses
_and_bacteria_(E._coli).tif.jpg)
In 2018, reanalysis of electron microscopy photographs from the 1980s allowed scientists to identify a giant virus (Meelsvirus) infecting Adhesisagitta hispida; its site of multiplication is nuclear and the virions (length: 1.25 μm) are enveloped.[29] In 2019, reanalysis of other previous studies has shown that structures that were taken in 1967 for bristles present on the surface of the species Spadella cephaloptera,[30] and in 2003, for bacteria infecting Paraspadella gotoi,[31] were in fact enveloped and spindle-shaped giant viruses with a cytoplasmic site of multiplication. The viral species infecting P. gotoi, whose maximum length is 3.1 μm, has been named Klothovirus casanovai (Klotho being the Greek name for one of the three Fates whose attribute was a spindle, and casanovai, in tribute to Pr J.-P. Casanova who devoted a large part of his scientific life to the study of chaetognaths). The other species has been named Megaklothovirus horridgei (in tribute to the first author of the 1967 article). On a photograph, one of the viruses M. horridgei, although truncated, is 3.9 μm long, corresponding to about twice the length of the bacteria Escherichia coli. Many ribosomes are present in virions but their origin remains unknown (cellular, viral or only partly viral). To date, giant viruses known to infect metazoans are exceptionally rare.
References
- Vannier J, Steiner M, Renvoisé E, Hu SX, Casanova JP (March 2007). "Early Cambrian origin of modern food webs: evidence from predator arrow worms". Proceedings of the Royal Society B: Biological Sciences. 274 (1610): 627–33. doi:10.1098/rspb.2006.3761. PMC 2197202. PMID 17254986.
- "Sagittoidea Claus and Grobben, 1905". Integrated Taxonomic Information System. Retrieved February 8, 2012.
- "World Register of Marine Species".
- Bone Q, Kapp H, Pierrot-Bults AC, eds. (1991). The Biology of Chaetognaths. London: Oxford University Press. ISBN 978-0-19-857715-7.
- Barnes, Robert D. (1982). Invertebrate Zoology. Philadelphia, PA: Holt-Saunders International. pp. 1046–1050. ISBN 978-0-03-056747-6.
- Grigor, Jordan J.; Schmid, Moritz S.; Caouette, Marianne; St.-Onge, Vicky; Brown, Thomas A.; Barthélémy, Roxane-M. (2020-07-01). "Non-carnivorous feeding in Arctic chaetognaths". Progress in Oceanography. 186: 102388. doi:10.1016/j.pocean.2020.102388. ISSN 0079-6611.
- Haddock SH, Case JF (20 January 1994). "A bioluminescent chaetognath". Nature. 367 (6460): 225–226. Bibcode:1994Natur.367..225H. doi:10.1038/367225a0.
- Thuesen EV, Goetz FE, Haddock SH (October 2010). "Bioluminescent organs of two deep-sea arrow worms, Eukrohnia fowleri and Caecosagitta macrocephala, with further observations on Bioluminescence in chaetognaths". The Biological Bulletin. 219 (2): 100–11. doi:10.1086/BBLv219n2p100. PMID 20972255.
- Jordan CE (1992). "A model of rapid-start swimming at intermediate Reynolds number: undulatory locomotion in the chaetognath Sagitta elegans". Journal of Experimental Biology. 163 (1): 119–137.
- Bone Q, Kapp H, Pierrot-Bults AC, eds. (1991). "The Tetrodotoxin Venom of Chaetognaths". The Biology of chaetognaths. Oxford University Press. pp. 55–60. ISBN 978-0-19-857715-7.
- Grigor, Jordan J.; Schmid, Moritz S.; Fortier, Louis (2017-11-01). "Growth and reproduction of the chaetognaths Eukrohnia hamata and Parasagitta elegans in the Canadian Arctic Ocean: capital breeding versus income breeding". Journal of Plankton Research. 39 (6): 910–929. doi:10.1093/plankt/fbx045. ISSN 0142-7873.
- Pond, David W. (2012-06-01). "The physical properties of lipids and their role in controlling the distribution of zooplankton in the oceans". Journal of Plankton Research. 34 (6): 443–453. doi:10.1093/plankt/fbs027. ISSN 0142-7873.
- "Photoreception". Encyclopædia Britannica from Encyclopædia Britannica 2006 Ultimate Reference Suite DVD . 2009.
- Terazaki, M.; Miller, C. B. (1982-11-01). "Reproduction of meso- and bathypelagic chaetognaths in the genus Eukrohnia". Marine Biology. 71 (2): 193–196. doi:10.1007/BF00394629. ISSN 1432-1793.
- Systema Naturae 2000 Taxon: Phylum Chaetognatha per Margulis and Schwartz Archived November 27, 2005, at the Wayback Machine (select Margulis & Schwartz in 'Classification by')—last retrieved November 25, 2006
- Systema Naturae 2000 Taxon: Phylum Chaetognatha per Cavalier-Smith Archived November 27, 2005, at the Wayback Machine (select Cavalier-Smith in 'Classification by')—last retrieved November 25, 2006
- Matus DQ, Copley RR, Dunn CW, Hejnol A, Eccleston H, Halanych KM, Martindale MQ, Telford MJ (August 2006). "Broad taxon and gene sampling indicate that chaetognaths are protostomes". Current Biology. 16 (15): R575-6. doi:10.1016/j.cub.2006.07.017. PMID 16890509.
- Marlétaz F, Martin E, Perez Y, Papillon D, Caubit X, Lowe CJ, Freeman B, Fasano L, Dossat C, Wincker P, Weissenbach J, Le Parco Y (August 2006). "Chaetognath phylogenomics: a protostome with deuterostome-like development". Current Biology. 16 (15): R577-8. doi:10.1016/j.cub.2006.07.016. PMID 16890510.
- Papillon D, Perez Y, Caubit X, Le Parco Y (November 2004). "Identification of chaetognaths as protostomes is supported by the analysis of their mitochondrial genome". Molecular Biology and Evolution. 21 (11): 2122–9. doi:10.1093/molbev/msh229. PMID 15306659.
- Rieger V, Perez Y, Müller CH, Lipke E, Sombke A, Hansson BS, Harzsch S (February 2010). "Immunohistochemical analysis and 3D reconstruction of the cephalic nervous system in Chaetognatha: Insights into the evolution of an early bilaterian brain?". Invertebrate Biology. 129 (1): 77–104. doi:10.1111/j.1744-7410.2010.00189.x.
- Harzsch S, Müller CH (May 2007). "A new look at the ventral nerve centre of Sagitta: implications for the phylogenetic position of Chaetognatha (arrow worms) and the evolution of the bilaterian nervous system". Frontiers in Zoology. 4: 14. doi:10.1186/1742-9994-4-14. PMC 1885248. PMID 17511857.
- Fröbius AC, Funch P (April 2017). "Rotiferan Hox genes give new insights into the evolution of metazoan bodyplans". Nature Communications. 8 (1): 9. Bibcode:2017NatCo...8....9F. doi:10.1038/s41467-017-00020-w. PMC 5431905. PMID 28377584.
- Marlétaz F, Peijnenburg KT, Goto T, Satoh N, Rokhsar DS (January 2019). "A New Spiralian Phylogeny Places the Enigmatic Arrow Worms among Gnathiferans". Current Biology. 29 (2): 312–318.e3. doi:10.1016/j.cub.2018.11.042. PMID 30639106.
- Kassatkina AP (25 December 2012). "New genus (Entokrohnia) and new species of family Tokiokaispadellidae Salvini-Plaven, 1986 (Сhaetognatha) from south-west Pacific Ocean" (PDF). Zoosystematica Rossica. 21 (2): 318–322.
- Chen JY, Huang DY (October 2002). "A possible Lower Cambrian chaetognath (arrow worm)". Science. 298 (5591): 187. doi:10.1126/science.1075059. PMID 12364798.
- Hu SX (2005). "Taphonomy and palaeoecology of the Early Cambrian Chengjiang Biota from Eastern Yunnan, China". Berliner Paläobiologische Abhandlungen. 7: 1–197.
- Szaniawski H (2005). "Cambrian chaetognaths recognized in Burgess Shale fossils" (PDF). Acta Palaeontologica Polonica. 50 (1): 1–8.
- Szaniawski H (2002). "New evidence for the protoconodont origin of chaetognaths" (PDF). Acta Palaeontologica Polonica. 47 (3): 405–419.
- Shinn GL, Bullard BL (2018-09-19). San Martin C (ed.). "Ultrastructure of Meelsvirus: A nuclear virus of arrow worms (phylum Chaetognatha) producing giant "tailed" virions". PLOS ONE. 13 (9): e0203282. Bibcode:2018PLoSO..1303282S. doi:10.1371/journal.pone.0203282. PMC 6145532. PMID 30231047.
- Horridge GA, Boulton PS (1967-11-14). "Prey detection by Chaetognatha via a vibration sense". Proceedings of the Royal Society of London. Series B. Biological Sciences. 168 (1013): 413–419. Bibcode:1967RSPSB.168..413H. doi:10.1098/rspb.1967.0072.
- Casanova JP, Duvert M, Goto T (October 2003). "Ultrastructural study and ontogenesis of the appendages and related musculature of Paraspadella (Chaetognatha)". Tissue & Cell. 35 (5): 339–51. doi:10.1016/S0040-8166(03)00055-7. PMID 14517101.
External links
| Wikimedia Commons has media related to Chaetognatha. |
- Image of Pseudosagitta gazellae with a krill in its gut from the Tasmanian Aquaculture and Fisheries Institute
- Chaetognatha of the World – last retrieved December 13, 2006
{kind=link}