Testudo (genus)
Testudo, the Mediterranean tortoises, are a genus of tortoises found in North Africa, Western Asia, and Europe. Several species are under threat in the wild, mainly from habitat destruction.
| Testudo | |
|---|---|
 | |
| Four tortoises of the genus Testudo. Clockwise from left: Testudo graeca ibera Testudo hermanni boettgeri Testudo hermanni hermanni Testudo marginata sarda. | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Reptilia |
| Order: | Testudines |
| Suborder: | Cryptodira |
| Superfamily: | Testudinoidea |
| Family: | Testudinidae |
| Genus: | Testudo Linnaeus, 1758 |
| Type species | |
| Testudo graeca | |
| Synonyms | |
| |
Background
They are small tortoises, ranging in length from 7.0 to 35 cm and in weight from 0.7 to 7.0 kg.
Systematics
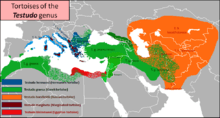
The systematics and taxonomy of Testudo is notoriously problematic. Highfield and Martin commented:
Synonymies on Testudo are notoriously difficult to compile with any degree of accuracy. The status of species referred has undergone a great many changes, each change introducing an additional level of complexity and making bibliographic research on the taxa extremely difficult. Most early and not a few later checklists contain a very high proportion of entirely spurious entries, and a considerable number of described species are now considered invalid – either because they are homonyms, non-binomial or for some other reason.[2]
Since then, DNA sequence data have increasingly been used in systematics, but in Testudines (turtles and tortoises), its usefulness is limited: In some of these, at least mtDNA is known to evolve more slowly in these than in most other animals.[3] Paleobiogeographical considerations suggest the rate of evolution of the mitochondrial 12S rRNA gene is 1.0-1.6% per million years for the last dozen million years or so in the present genus[4] and ntDNA evolution rate has been shown to vary strongly even between different population of T. hermanni;[5] this restricts sequence choice for molecular systematics and makes the use of molecular clocks questionable.
The following extant species are placed here:
- Russian tortoise or Horsfield's tortoise, T. horsfieldii[1]
- Hermann's tortoise, T. hermanni[1]
- Spur-thighed tortoise, Greek tortoise or common tortoise, T. graeca[1]
- Subspecies:
- Mediterranean spur-thighed tortoise, T. graeca graeca[1]
- Araxes tortoise, T. graeca armeniaca[1]
- Buxton's tortoise, T. graeca buxtoni[1]
- Cyrenaican spur-thighed tortoise, T. graeca cyrenaica[1]
- Asia Minor tortoise, T. graeca ibera[1]
- Morocco tortoise, T. graeca marokkensis[1]
- Nabeul tortoise, T. graeca nabeulensis[1]
- Souss Valley tortoise, T. graeca soussensis[1]
- Mesopotamian tortoise, T. graeca terrestris[1]
- Iranian tortoise, T. graeca zarudnyi[1]
- Subspecies:
- Egyptian tortoise or Kleinmann's tortoise, T. kleinmanni[1]
- Marginated tortoise, T. marginata[1]
The first two are more distinct and ancient lineages than the closely related latter three species. Arguably, T. horsfieldii belongs in a new genus (Agrionemys) on the basis of the shape of its carapace and plastron,[6] and its distinctness is supported by DNA sequence analysis.[7] Likewise, a separate genus Eurotestudo has recently been proposed for T. hermanni; these three lineages were distinct by the Late Miocene as evidenced by the fossil record.[8] Whether these splits will eventually be accepted remains to be seen. The genus Chersus has been proposed to unite the Egyptian and marginated tortoises which have certain DNA sequence similarities,[4] but their ranges are (and apparently always were) separated by their closest relative T. graeca and the open sea and thus, chance convergent haplotype sorting would better explain the biogeographical discrepancy.
Conversely, the Greek tortoise is widespread and highly diverse. In this and other species, a high number of subspecies has been described, but not all generally accepted, and several (such as the "Negev tortoise" and the "dwarf marginated tortoise") are now considered to be local morphs. Some, such as the Tunisian tortoise, have even been separated as a separate genus Furculachelys, but this is not supported by more recent studies.[9]
Mating
Testudo spp. are promiscuous creatures and they follow a polyandrous mating system.[10] Mating involves a courtship ritual of mechanical, olfactory and auditory displays elicited from the male to coerce a female into accepting copulation.[11] Courtship displays are very energetically costly for males, especially because females tend to run away from courting males.[12] The male will chase her, exerting more energy and elaborate displays of ramming and biting. Females are able to judge a male's genetic quality through these displays; only healthy males are able to perform costly courting rituals, suggesting endurance rivalry.[11] These are considered honest signals that are then used to influence pre- and post-copulatory choice, as females are the choosy sex.[10]
Female mate choice offers no direct benefits (such as access to food or territory or parental care).[13] There are, however, indirect benefits of mating with multiple males. Engaging in a polyandrous mating system offers a female guaranteed fertilization, higher offspring diversity and sperm competition to ensure that eggs are fertilized by a high quality male. This is in respect to the "good genes" hypothesis that females receive indirect benefits through her offspring by mating with a quality male, "a male's contribution to a female's fitness is restricted to [his] genes" (Cutuli, G. et al., 2014).
Mating order has no influence on paternity of a clutch so a female's inclination to mate with multiple males and her ability to store sperm allows for sperm competition and suggests cryptic female choice.[14] However, some species do show size-assortative, T. marginata, for example, where large males breed with large females and small males breed with small females.[11] Other species form hierarchies; during male-to-male competition the more aggressive male is considered alpha.[10] Alpha males are more aggressive with their courting as well and have higher mounting success rate than beta males.
A female's reproductive tract contains sperm storage tubules and she is capable of storing sperm for up to four years.[15] This sperm remains viable and when she goes a breeding season without encountering a male she is able to fertilize her eggs with the stored sperm. Storing sperm can also result in multiple paternity clutches; It is quite common among Testudo spp. females to lay a clutch that has been sired by multiple males. And females can lay one to four clutches a breeding season. Sexual dimorphism, promiscuity, long term sperm storage and elaborate courting rituals are factors that effect mate preference, sperm competition and cryptic female choice in genus Testudo.[10]
References
- Rhodin, Anders G.J.; Inverson, John B.; Roger, Bour; Fritz, Uwe; Georges, Arthur; Shaffer, H. Bradley; van Dijk, Peter Paul (August 3, 2017). "Turtles of the world, 2017 update: Annotated checklist and atlas of taxonomy, synonymy, distribution, and conservation status(8th Ed.)" (PDF). Chelonian Research Monographs. 7. ISBN 978-1-5323-5026-9. Retrieved October 4, 2019.
- Highfield, A. C. & Martin, J. (1989). "A revision of the Testudines of North Africa, Asia, and Europe. Genus: Testudo". Journal of Chelonian Herpetology. 1 (1): 1–12.
- "Mitochondrial DNA evolution at a turtle's pace: Evidence for low genetic variability and reduced microevolutionary rate in the Testudines". Molecular Biology and Evolution. 1992. doi:10.1093/oxfordjournals.molbev.a040735. PMID 1584014.
- Van Der Kuyl, Antoinette C.; Ph. Ballasina, Donato L.; Dekker, John T.; Maas, Jolanda; Willemsen, Ronald E.; Goudsmit, Jaap (2002). "Phylogenetic Relationships among the Species of the Genus Testudo (Testudines: Testudinidae) Inferred from Mitochondrial 12S rRNA Gene Sequences". Molecular Phylogenetics and Evolution. 22 (2): 174–183. doi:10.1006/mpev.2001.1052. PMID 11820839.
- Fritz, Uwe; Auer, Markus; Bertolero, Albert; Cheylan, Marc; Fattizzo, Tiziano; Hundsdorfer, Anna K.; Martin Sampayo, Marcos; Pretus, Joan L.; Široky, Pavel; Wink, Michael (2006). "A rangewide phylogeography of Hermann's tortoise, Testudo hermanni (Reptilia: Testudines: Testudinidae): Implications for taxonomy". Zoologica Scripta. 35 (5): 531–543. doi:10.1111/j.1463-6409.2006.00242.x.
- Khozatsky, L.I. & Mlynarski, M. (1966): Agrionemys - nouveau genre de tortues terrestres (Testudinidae) ["Agrionemys - a new genus of tortoises"]. [Article in French] Bulletin de l'Académie Polonaise des Sciences II - Série des Sciences Biologiques 2: 123-125.
- Fritz, Uwe; Široký, Pavel; Kami, Hajigholi; Wink, Michael (2005). "Environmentally caused dwarfism or a valid species—Is Testudo weissingeri Bour, 1996 a distinct evolutionary lineage? New evidence from mitochondrial and nuclear genomic markers". Molecular Phylogenetics and Evolution. 37 (2): 389–401. doi:10.1016/j.ympev.2005.03.007. PMID 16223676.
- De Lapparent De Broin, France; Bour, Roger; Parham, James F.; Perälä, Jarmo (2006). "Eurotestudo, a new genus for the species Testudo hermanni Gmelin, 1789 (Chelonii, Testudinidae)". Comptes Rendus Palevol. 5 (6): 803–811. doi:10.1016/j.crpv.2006.03.002.
- Van Der Kuyl, Antoinette C.; Ballasina, Donato LP; Zorgdrager, Fokla (2005). "Mitochondrial haplotype diversity in the tortoise species Testudo graeca from North Africa and the Middle East". BMC Evolutionary Biology. 5: 29. doi:10.1186/1471-2148-5-29. PMC 1097724. PMID 15836787.
- Cutuli, Giulia; Cannicci, Stefano; Vannini, Marco; Fratini, Sara (2014). "Influence of male courtship intensity and male-male competition on paternity distribution in Hermann's tortoise, Testudo hermanni hermanni (Chelonia: Testudinidae)". Biological Journal of the Linnean Society. 111 (3): 656–667. doi:10.1111/bij.12243.
- Sacchi, Roberto; Galeotti, Paolo; Fasola, Mauro; Ballasina, Donato (2003). "Vocalizations and courtship intensity correlate with mounting success in marginated tortoises Testudo marginata". Behavioral Ecology and Sociobiology. 55: 95–102. doi:10.1007/s00265-003-0685-1.
- Galeotti, Paolo; Sacchi, Roberto; Rosa, Daniele Pellitteri; Fasola, Mauro (2005). "Female preference for fast-rate, high-pitched calls in Hermann's tortoises Testudo hermanni". Behavioral Ecology. 16: 301–308. doi:10.1093/beheco/arh165.
- Cutuli, Giulia; Cannicci, Stefano; Vannini, Marco; Fratini, Sara (2013). "Influence of mating order on courtship displays and stored sperm utilization in Hermann's tortoises (Testudo hermanni hermanni)". Behavioral Ecology and Sociobiology. 67 (2): 273–281. doi:10.1007/s00265-012-1447-8.
- Johnston, Emily E.; Rand, Matthew S.; Zweifel, Stephan G. (2006). "Detection of multiple paternity and sperm storage in a captive colony of the central Asian tortoise, Testudo horsfieldii". Canadian Journal of Zoology. 84 (4): 520–526. doi:10.1139/Z06-023.
- Roques, S.; Díaz-Paniagua, C.; Andreu, A. C. (2004). "Microsatellite markers reveal multiple paternity and sperm storage in the Mediterranean spur-thighed tortoise, Testudo graeca". Canadian Journal of Zoology. 82: 153–159. doi:10.1139/Z03-228.
External links
- Mediterranean tortoises
- Tortoise Trust Web - Basic care of Mediterranean tortoises.
- James F. Parham; J. Robert Macey; Theodore J. Papenfuss; Chris R. Feldman; Oguz Türkozan; Rosa Polymeni; JeVrey Boore (2005). "The phylogeny of Mediterranean tortoises and their close relatives based on complete mitochondrial genome sequences from museum specimens" (PDF). Cite journal requires
|journal=(help)
Tortoise family | ||||
|---|---|---|---|---|
| ||||
| Genera |
|  | ||
| Agrionemys | ||||
| Aldabrachelys |
| |||
| Astrochelys | ||||
| Centrochelys | ||||
| Chelonoidis |
| |||
| Galápagos tortoise |
| |||
| †Cheirogaster |
| |||
| Chersina | ||||
| Chersobius | ||||
| †Cylindraspis | ||||
| Geochelone | ||||
| Gopherus |
| |||
| †Hadrianus |
| |||
| †Hesperotestudo |
| |||
| Homopus | ||||
| Indotestudo | ||||
| Kinixys | ||||
| Malacochersus | ||||
| Manouria | ||||
| †Megalochelys |
| |||
| Psammobates | ||||
| Pyxis | ||||
| Stigmochelys | ||||
| †Stylemys |
| |||
| Testudo |
| |||
Phylogenetic arrangement of turtles based on turtles of the world 2017 update: Annotated checklist and atlas of taxonomy, synonymy, distribution, and conservation status. Key: †=extinct. | ||||